
Abstract
Abstract
The less differentiated the donor cells are used in nuclear transfer (NT), the more easily are they reprogrammed by the recipient cytoplasm. In this context, mesenchymal stem cells (MSCs) appear as an alternative to donor nuclei for NT. The amniotic fluid and adipose tissue are sources of MSCs that have not been tested for the production of cloned embryos in cattle. The objective of this study was to isolate, characterize, and use MSCs derived from amniotic fluid (MSC-AF) and adipose tissue (MSC-AT) to produce cloned calves. Isolation of MSC-AF was performed using in vivo ultrasound-guided transvaginal amniocentesis, and MSC-AT were isolated by explant culture. Cellular phenotypic and genotypic characterization by flow cytometry, immunohistochemistry, and RT-PCR were performed, as well as induction in different cell lineages. The NT was performed using MSC-AF and MSC-AT as nuclear donors. The mesenchymal markers of MSC were expressed in bovine MSC-AF and MSC-AT cultures, as evidenced by flow cytometry, immunohistochemistry, and RT-PCR. When induced, these cells differentiated into osteocytes, chondrocytes, and adipocytes. Embryo production was similar between the cell types, and two calves were born. The calf from MSC-AT was born healthy, and this fact opens a new possibility of using this type of cell to produce cloned cattle by NT.
Introduction
C
In this context, the choice of the donor cell appears to be important for improvement of the technique and may have a large effect on the progression of development (Miyoshi et al., 2003). Prevention of epigenetic errors has been suggested as indispensable to improving the success rate of animal cloning (Wang et al., 2011). This makes the availability of cells whose nuclei are able to undergo epigenetic reprogramming an important prerequisite for NT and makes the donor cells a crucial component in the cloning process (Fu et al., 2008; Wani et al., 2010).
There are significant differences in the ability of various cell lines to produce embryos, and the nuclei of cells that are less differentiated seem to be better at supporting full embryo development than cells that are completely differentiated (Hochedlinger and Jaenisch, 2002). In this sense, multipotent cells, such as embryonic blastomeres and embryonic stem cells, support the development of NT embryos at a higher efficiency when compared to somatic cells (Campbell et al., 2007). Results in some experiments with mice (Rideout et al., 2000; Wakayama et al., 1998) suggest that the undifferentiated state of the donor cells may increase the birth rate of cloned animals. Thus, mesenchymal stem cells (MSCs) emerge as a promising source of cells for use in NT.
MSCs are multipotent stem cells with well-defined characteristics and an intrinsic capacity for self-regeneration and differentiation into functional cell types (Baksh et al., 2004). These cells have a promising application in regenerative medicine for humans (Wagner et al., 2005) and domestic animals (Cremonesi et al., 2011). New sources of MSCs are being discovered and studied, such as the fetal components, including cells of the amniotic fluid and cells derived from the umbilical cord, and adipose tissue. MSCs have been characterized in some species of domestic animals, such as horses (De Mattos Carvalho et al., 2009) and dogs (Filioli Uranio et al., 2011). Recently, MSCs derived from Wharton's jelly (Cardoso et al., 2012) and amniotic fluid (Corradetti et al., 2013; Gao et al., 2014) have been characterized in cattle. Although MSCs derived from amniotic fluid (MSC-AF) and from adipose tissue (MSC-AT) are considered a new therapeutic tool for repair or regeneration of damaged organs (Ghorbani et al., 2014), and a promising source of donor cells for NT, they have not yet been used to produce cloned cattle embryos. If the undifferentiated state of the donor cells may affect the success rate of cloning as cited by Zheng et al. (2009), the use of MSC-AF and MSC-AT may increase cloning efficiency.
This study aimed to isolate, cultivate, and characterize bovine MSC-AF and MSC-AT and test their ability to produce blastocysts, pregnancies, and calves. The results showed that MSC-AT were able to generate a healthy calf with an efficiency greater than that reported in the literature, therefore indicating that such cells can be used in NT.
Materials and Methods
Ethics in experimentation
All of the procedures of experimentation were approved by the Ethics Committee in Animal Use at the University of Brasília (protocol no. 151101/2013).
Isolation and culture of MSCs from amniotic fluid cells
Ultrasound-guided transvaginal amniocentesis was performed to collect the amniotic fluid cells from four adult Guzerat cows, with pregnancies between 60 and 74 days. The procedure of amniocentesis was performed according to Garcia and Salaheddine (1997) and Kamimura et al. (1997), with modifications. Briefly, a 7.5-MHz convex transducer was connected to a Honda ultrasound system (model HS-1500V, Japan) and equipped with a 21-gauge needle 65 cm in length, localized in the tip transducer. When the needle reached the amniotic fluid, about 7 mL of fluid was aspirated with a 60-mL sterile syringe (Fig. 1). A volume of amniotic fluid sufficient to maintain the pregnancy was preserved.
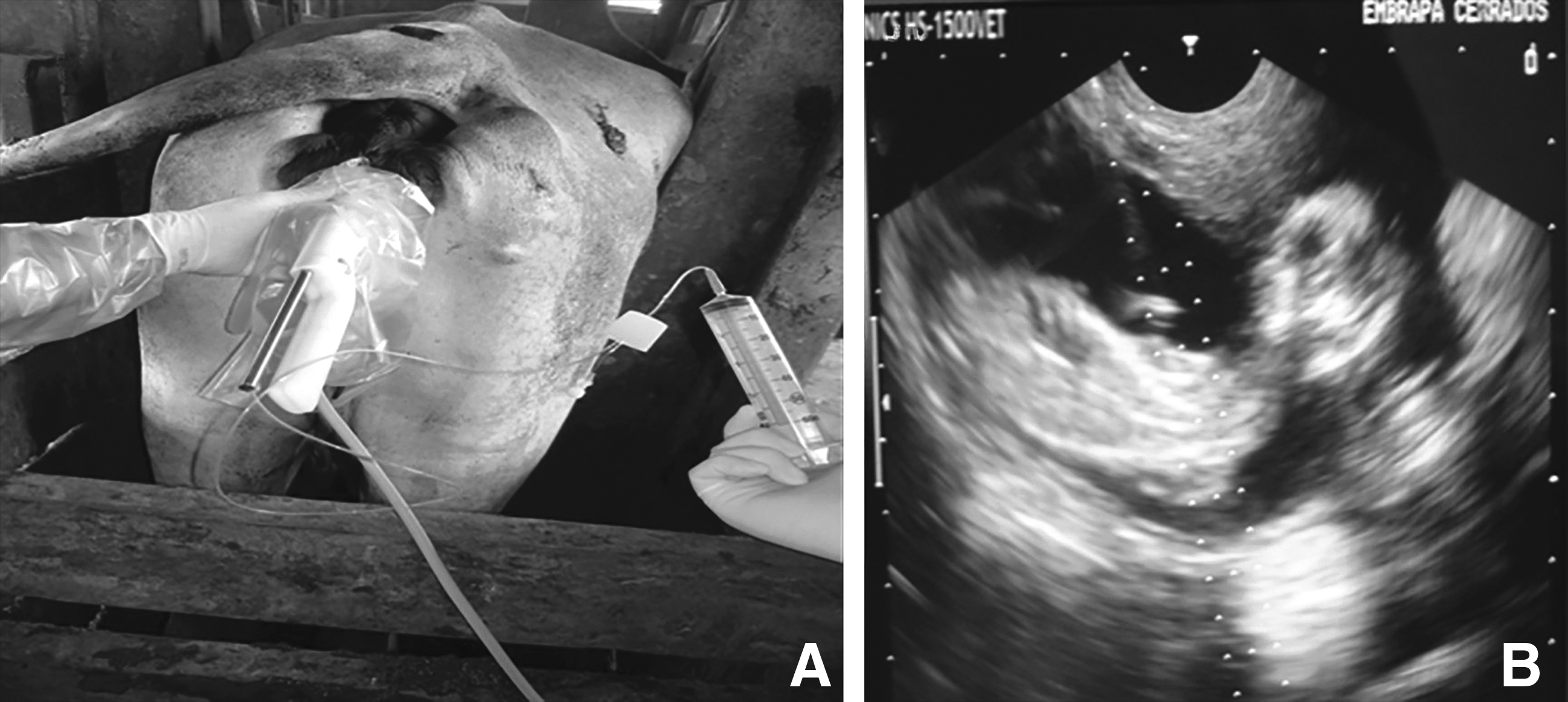
(
The amniotic fluid was centrifuged at 135 × g for 10 min, and the supernatant was discarded. The sediment was resuspended in 3 mL of AmnioMAX-II Complete Medium (Gibco-BRL/Life Technologies, Rockville, MD, USA). The cells were cultured in special culture flasks in an incubator with 5% CO2, 90% humidity, and 38.5°C.
Isolation and culture of MSCs from adipose tissue
The adipose tissue was collected from the same animals from which amniotic fluid was recovered. Seven-month-old calves underwent epidural anesthesia with lidocaine 2% (wt/vol). The area for collection of the tissue was shaved, cleaned, and sterilized with iodized alcohol. The 2-cm × 1-cm biopsy, consisting of skin and adipose tissue, was collected from the perineal region and immediately immersed in phosphate-buffered saline (PBS) at 37°C. In the laboratory, incisions were made with a sterile scalpel along the adipose tissue, and small biopsies (0.5 cm ×0.5 cm) were obtained, exposing a larger area of tissue to increase the contact region with the plastic surface of the plate. The explants were cultured in Dulbecco's Modified Eagle Medium (DMEM; Invitrogen Life Science, Rockville, MD, USA) supplemented with 10% fetal bovine serum (FBS).
In vitro multilineage differentiation assay
Adipogenic, osteogenic, and chondrogenic differentiations were performed according to the manufacturer's instructions (StemPro Differentiation Medium, Invitrogen). For the osteogenic differentiation procedure, cells were plated at 1 × 106 cells/mL in a six-well culture plate under the same culture conditions described above. After seeding for 48 h, the medium was replaced with osteogenic differentiation medium (2 mL/well), and the medium was replaced with fresh medium every 4 days. After 20 days of differentiation, the cells were fixed with 4% paraformaldehyde (Sigma-Aldrich, St. Louis, MO, USA), and calcium mineralization was assessed by Alizarin Red staining (Sigma-Aldrich). In this procedure, alkaline phosphatase activity was measured by p-nitrophenyl phosphatase (p-NPP) substrate reactions using ALP™ assay reagents (Sigma-Aldrich). In brief, cells were washed twice with 2 mL of PBS and with 0.2% Triton X-100 by shaking for 20 min. The cell layers were incubated with 500 μL of substrate (10 mM p-NPP, 1 mM MgCl2) for 30 min at 37°C. The reaction was stopped by adding 100 μL of 1 mM NaOH. The p-NNP formed was measured using a microplate reader with a 405-nm filter (Multiskan, LabSystems, NY, USA).
For adipogenic differentiation, 1 × 106 cells/mL were plated in the same manner described for osteogenic differentiation. The cultured medium was changed to 2 mL of StemPro Adipogenic Differentiation Medium, which was replaced every 24 h. After 15 days of differentiation, the cells were fixed with 4% paraformaldehyde and stained with Oil Red solution (0.3% Oil Red powder dissolved in 60% isopropanol) for 10 min. Thereafter, cells were washed with 60% isopropanol. The induction of adipogenic differentiation was apparent by intracellular accumulation of lipid rich vacuoles that were stained with Oil Red. To induce chondrogenic differentiation, similar culture conditions were used as described for adipogenic and osteogenic induction. For this, 1 × 106 cells/mL were seeded, and the StemPro Differentiation Medium was supplemented with 100 μL of chondrogenic inducer (Invitrogen). The cells were kept under this condition for 1 week, and the medium was replaced every 24 h. The cells were fixed with 4% paraformaldehyde and stained with Safranin O to identify glycosaminoglycans.
Immunocytochemistry
Cell staining was performed using the primary monoclonal antibodies described in Table 1. Cells were fixed with 4% paraformaldehyde for 15 min after 48 h of culture in Lab-Tek Chamber Slides (Nunc™, Thermo Scientific, Rochester, NY, USA). The cells were permeabilized for 10 min at room temperature in 0.4% Triton X-100 diluted in PBS. These were then incubated overnight with primary antibodies at 4°C. After three washes, the cells were incubated with secondary antibody rabbit anti-mouse fluorescein isothiocyanate (FITC; Sigma-Aldrich) at a 1:100 dilution. For nuclear staining, 4′,6-diamidino-2-phenylindole (DAPI; 1 mg/mL) was diluted in PBS and loaded onto samples for 15 min. The images were collected under an AxioImager A.1 light and an ultraviolet microscope connected to an AxioCam MRc digital camera (Carl Zeiss, Oberkochen, Germany). Images were processed using AxioVision 4.8 software (Carl Zeiss).
FC, flow cytometer; IC, immunocytochemistry; MSCs, mesenchymal stem cells.
Flow cytometry
Approximately 1 × 106 cells were harvested after detachment with 0.25% trypsin (Sigma-Aldrich), washed in PBS, and incubated for 18 h at 4°C with the same monoclonal antibodies described for immunocytochemistry (Table 1). After incubation with primary antibodies, the cells were washed three times with PBS plus 0.1% Triton X-100. A 1:50 dilution of the secondary antibody was added to 100 μL of cell suspension, and the cell suspension was incubated at 37°C for 30 min. The cell suspension was washed as previously described, and after the final wash, the cells were fixed with 4% paraformaldehyde. Data were acquired with the Attune™ Acoustic Focusing Cytometer system (Applied Biosystems, Foster City, CA, USA). The negative pattern was examined by applying the same cell suspension with the first incubation, and the result was included in the global compensation to exclude autofluorescence. A BL1-A (488-nm) filter (Applied Biosystems) was used in each analysis.
Reverse transcriptase polymerase chain reaction (RT-PCR)
Cell RNA was isolated from amniotic fluid and adipose tissue using TRIzol® (Invitrogen). RNA concentrations were measured by absorbance at 260 nm with Biophotometer (Eppendorf, Hamburg, Germany), and 2 μg of total RNA was used for the reverse transcription reaction using Superscript II® Reverse Transcriptase (Invitrogen). The cDNA was amplified using Platinum® Taq DNA Polymerase High Fidelity (Invitrogen). The primers used were designed according to the GenBank database (Table 2). The RNA templates were amplified at 33–45 cycles of 94°C (30 sec), 58°C to 61°C (30 sec), 72°C (1 min), followed by amplification at 72°C for 10 min. PCR products were detected with ethidium bromide in a 1.2% agarose gel.
F, forward; R, reverse.
Nuclear transfer
Skin fibroblasts, MSC-AF, and MSC-AT were used as donor nuclei in the NT procedure when 100% of confluence had been achieved. Briefly, oocytes from abattoir ovaries were matured for 18 h, and those with the first apparent polar body were selected for enucleation. The polar body and metaphase plate were removed with the aid of Narishige micromanipulators (model IM-9B, Narishige, East Meadow, NY, USA) using an Nikon inverted microscope (model Ti-S, Nikon Instruments, Melville, NY, USA), and one cell was injected into the perivitelline space.
Fusion of karyoplast–cytoplast couplets was performed in a microslide fusion chamber with stainless steel electrodes separated by a 1-mm gap. The chamber was filled with fusion buffer (0.3 M mannitol). The couplets were fused by two consecutive DC pulses (pulse, 140 volts; alignment frequency, 100 kHz; pulse time, 20 μs; amplitude, 0.7–14 peak volts) delivered by an electron cell fusion/activation system (Voltain EP-1, Cryologic, Blackburn, Victoria, Australia). Couplets were evaluated for fusion by microscopic examination within a 1-h period. Chemical activation was performed with a 5-min exposure to 5 μM of ionomycin and 4 h in 1.9 mM 6-dimethylaminopurine (DMAP). After activation, the embryos were cultured in synthetic oviductal fluid (SOF) with 5% FBS and co-culture of cumulus cells until 8 days. Mature, nonmanipulated oocytes were activated chemically, and embryos were cultured as described previously. These embryos were used as a parthenogenetic control.
Embryo transfer and pregnancy diagnosis
Blastocysts from both cell types were transferred to crossbred recipient cows aged from 2 to 5 years that presented natural estrus. The diagnosis of pregnancy in the recipient cows was carried out by ultrasound 35 days after embryo transfer. The pregnancy was monitored 45 and 60 days after embryo transfer and every 30 days until delivery.
Calving of cloned calves
When pregnancy presented a complication that prevented vaginal delivery, cesarean section was performed.
Microsatellite Analysis
The genomic DNA was compared among samples collected from donor animals and cloned calves using an automated DNA sequencer and kits specific for the amplification of polymorphic loci microsatellites, according to the protocol of the International Society for Animal Genetics (ISAG), instruction no. 74 of October 20, 2004 (Ministry of Agriculture Livestock and Supply–MAPA, Brazil) and Instruction no. 17 of 09 August 2012 (MAPA).
Statistical analysis
The experimental design was completely randomized, with four repetitions (NT procedures). The treatments were MSC-AF and MSC-AT. The variables in percentage format were transformed in “root arcsine (x/100).” The statistical verification of the significance of the treatments was done by analysis of variance (ANOVA). The t-test and Tukey test were used to compare the averages at 5% probability. All the analyses were performed by statistical software SAS version 9.1.2.
Results
Cell isolation
One animal that underwent amniocentesis aborted after the procedure, whereas the other three presented normal pregnancies and delivered healthy calves. The collection of adipose tissue was performed using two animals.
Characterization of MSC-AF and MSC-AT
The expression patterns found by immunophenotyping MSC-AF and MSC-AT revealed that 70% and 86% of isolated cells from amniotic fluid and adipose tissue, respectively, were positive for CD29, CD73, CD90, and CD105 MSC markers. MSC-AF were negative for CD34, CD44, and CD45 (Fig. 2), but MSC-AT were negative only for CD34 and CD45 (Fig. 3). Similarly, flow cytometry corroborated the results obtained by immunocytochemistry, as observed in Figure 4. MSC-AT were positive for CD44, whereas MSC-AF were not. Additionally, bovine-derived MSC-AF and MSC-AT exhibited a typical MSC genotype, with a gene expression profile that includes mesenchymal cell markers, as demonstrated by RT-PCR (Fig. 5).
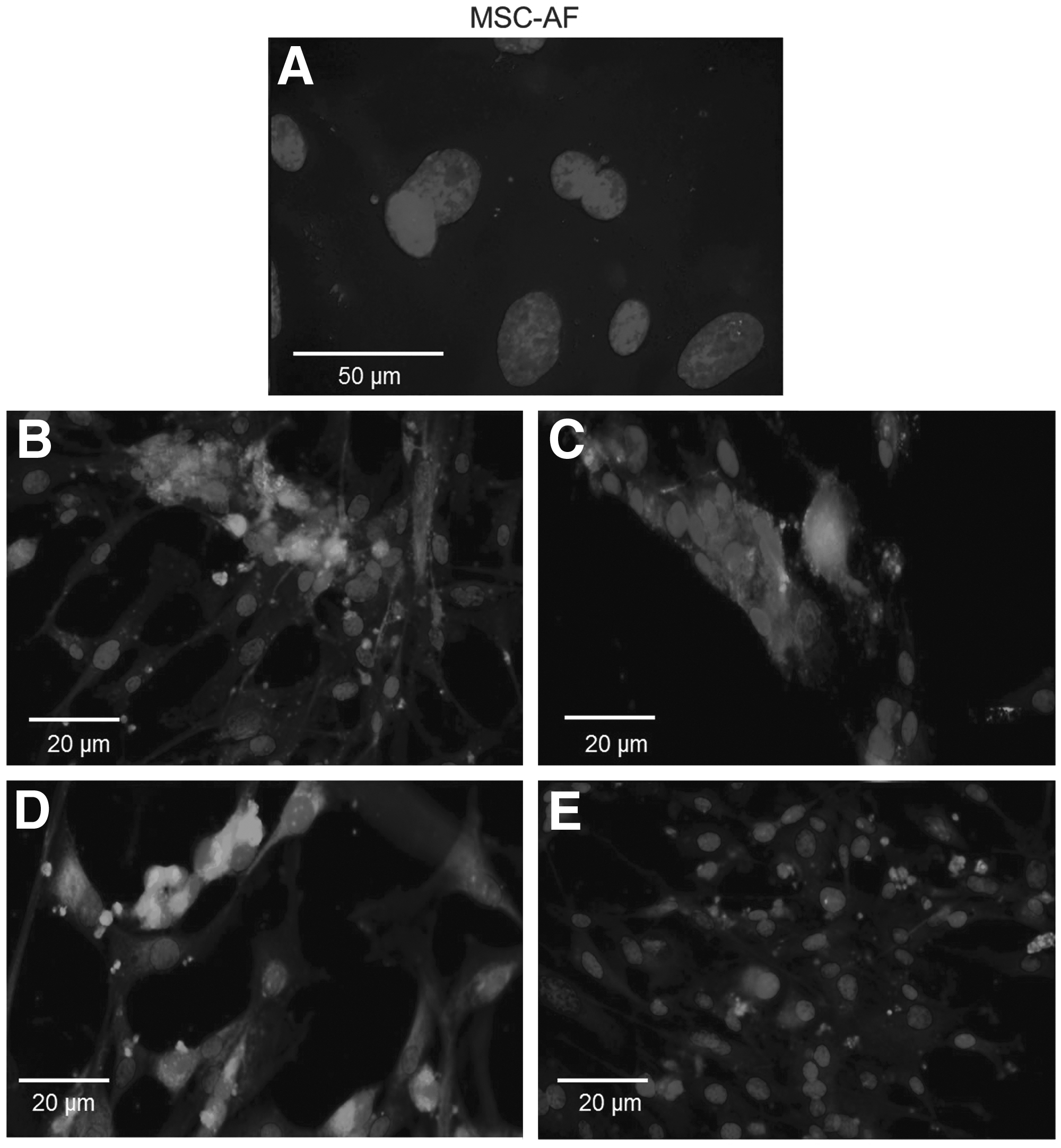
Micrographs of immunocytochemical analyses performed on MSC-AF, demonstrating that the cells had no surface markers CD34, CD44, and CD45 (
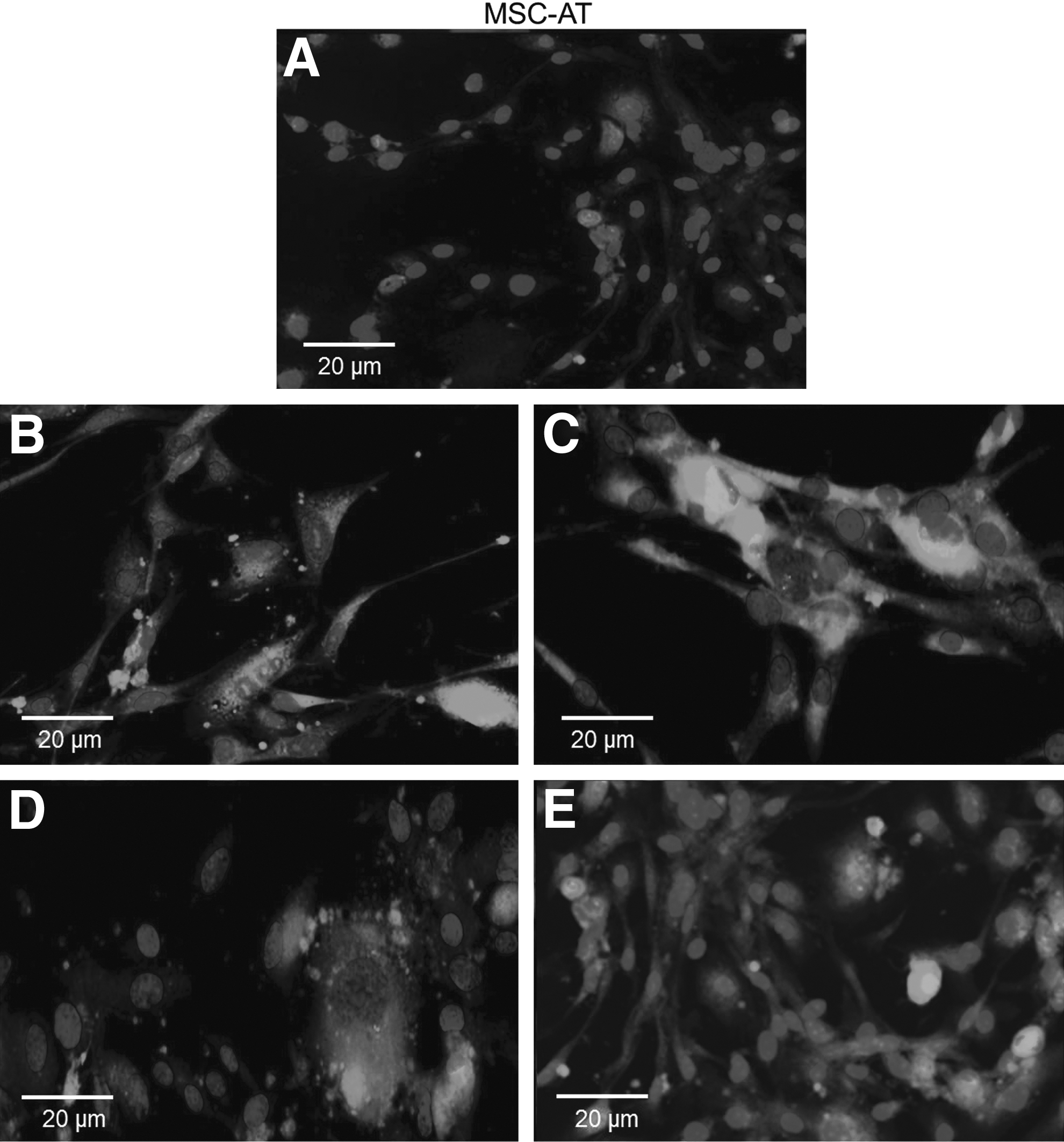
Micrographs of immunocytochemical analyses performed on MSC-AT, demonstrating that the cells had no surface markers CD34 and CD45 (
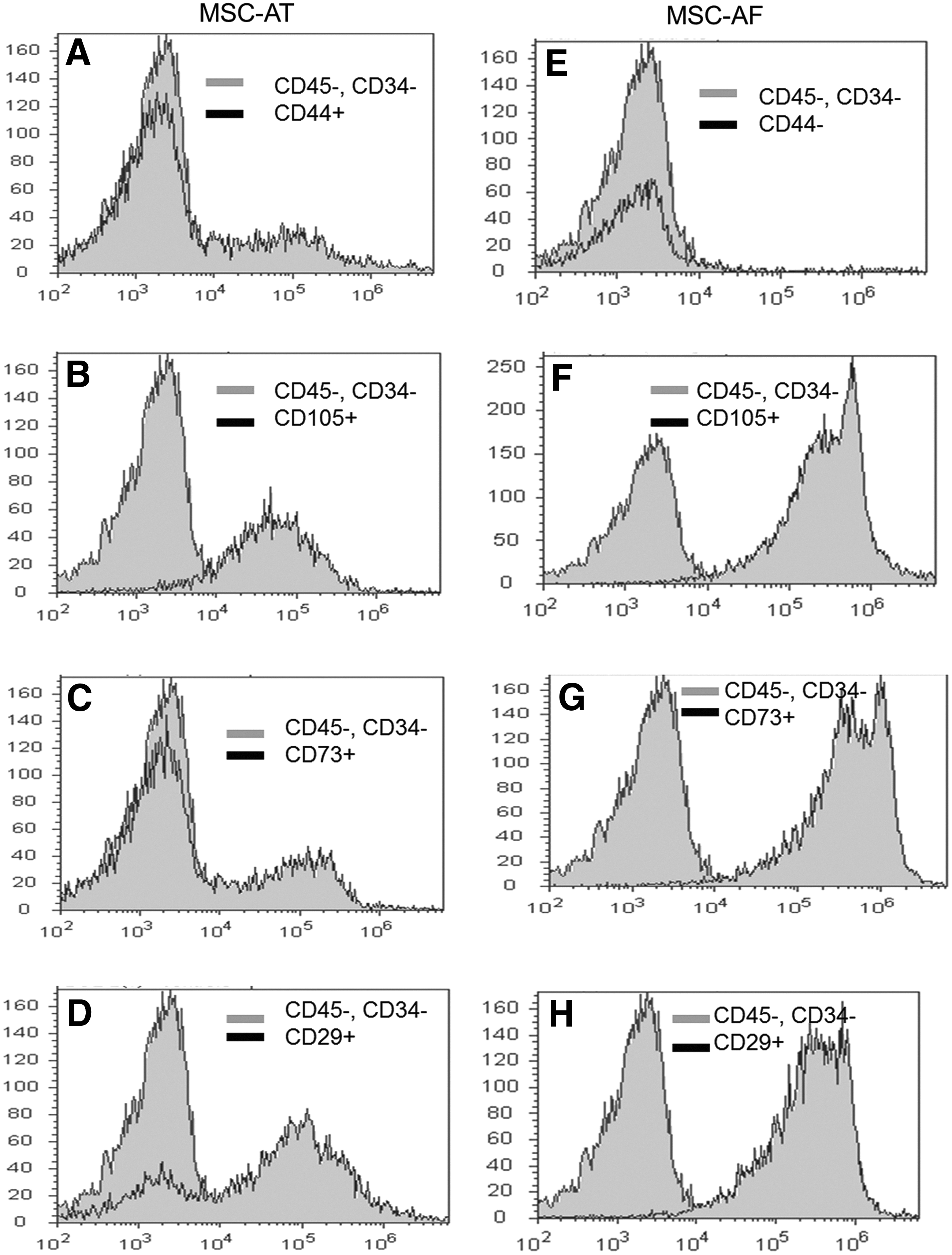
Histograms of flow cytometric analyses of surface marker expression in bovine mesenchymal stem cells derived from amniotic fluid (MSC-AF) and adipose tissue (MSC-AT). The cells presented the surface antigen required for their characterization as mesenchymal stem cells. Data were processed by Attune™ acoustic focusing cytometer and excluded autofluorescence as a global compensation tool (≤103). The y axis represents a log scale.
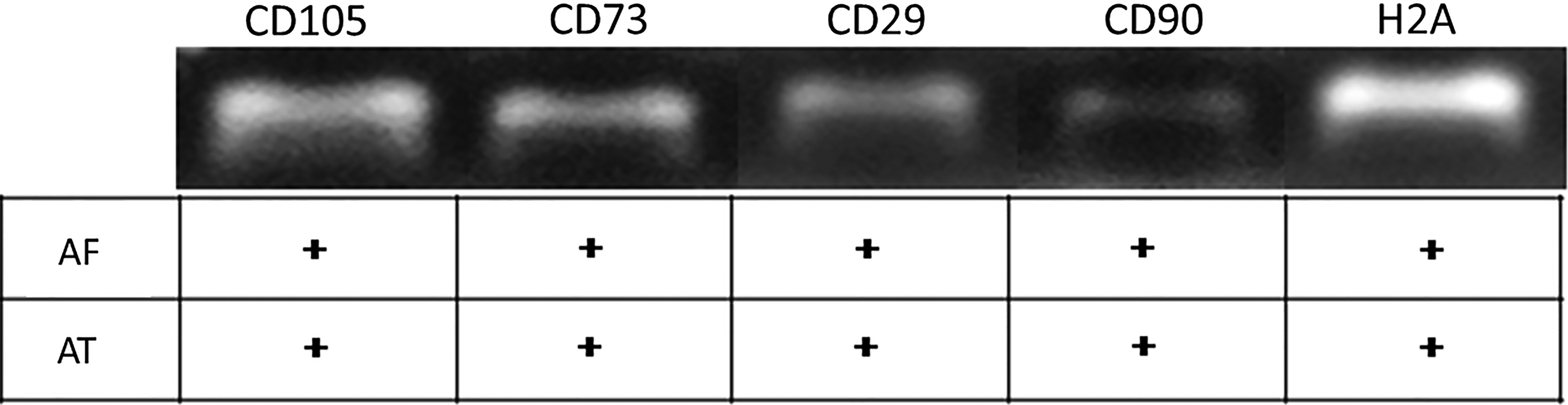
A typical profile of mesenchymal stem cells was exhibited by bovine-derived amniotic fluid (AF) and adipose tissue (AT) cells using reverse transcription-polymerase chain reaction.
The differentiation potential was verified by the ability of the cells to differentiate into osteocytes, and adipocytes, as confirmed by Alizarin Red, Safranin O, and Oil Red stain tests, respectively (Fig. 6). Cells fixed and stained with Alizarin Red demonstrated matrix calcification (Fig. 6A), indicating osteogenic differentiation. For chondrogenic differentiation after Safranin O staining, the presence of glycosaminoglycans was observed as red deposits in the cells (Fig. 6B). Finally, adipogenic differentiation was shown by lipid vacuoles inside the cells stained positively using Oil Red (Fig. 6C).
Phenotypic characterization of mesenchymal cells derived from bovine amniotic fluid and adipose tissue isolated after the tenth passage. (
Developmental ability of embryos cloned from different donor cells
The NT procedure using micromanipulators was used to test cloning competence of skin fibroblasts, MSCs from amniotic fluid, and MSCs from adipose tissue. The fusion rate was higher for MSC-AF than MSC-AT and skin fibroblasts (85.89%, 50.06%, and 44.85%, respectively). Rates of cleavage were superior for embryos derived from skin fibroblasts (70.64%) when compared with MSC-AF and MSC-AT rates (39.08 and 38.26, respectively). Finally, for blastocyst development rates (17.27%, 17.87%, 19.00%), there were no differences among embryos derived from MSC-AF, MSC-AT, and skin fibroblasts (Table 3).
Different superscript letters (a and b) in the columns represent significant difference (p < 0.05).
MSC-AF, mesenchymal stem cells derived from amniotic fluid; MSC-AT, mesenchymal stem cells derived from adipose tissue; FB, skin fibroblasts.
Pregnancies of embryos produced by NT
To evaluate the developmental efficiency of in vivo embryos produced from each cell type, eight blastocysts from MSC-AF were transferred to five recipient cows and four embryos from MSC-AT were transferred to four recipient cows. The highest pregnancy rate was obtained with MSC-AT embryos (Table 3). One pregnancy was established from MSC-AF (eight blastocysts/five recipients) and also one pregnancy was established from MSC-AT (four blastocysts/four recipients).
Birth of cloned calves
The cloned calf from MSC-AF was born at 277 days of gestation by cesarean surgery, because the cow recipient presented hydropsy. The cloned calf weighed 58.5 kg and died shortly after birth because of a mechanical obstruction of airways by fluid that made it difficult for the animal to breathe.
Unlike the previous calving, the calf from the MSC-AT was born after 291 days of gestation through assisted normal delivery. The calf was healthy and weighed 35 kg. Figure 7 shows the physical similarities between the donor cell animal shortly after birth and the clones obtained from MSC-AT.

(
Discussion
By the year 2002, more than 200 mammalian cell types were clearly distinguishable by morphology, but less than 5% of these cells had been tested as donors in NT (Oback and Wells, 2002). Our results, using mesenchymal cells from bovine amniotic fluid and adipose tissue in NT, expand the list of donor cell types that have generated live cloned calves. As has been previously established in several studies in the literature, with sheep (Wilmut et al., 1997), cattle (Cheng et al., 2011; Galli et al., 2002; Green et al., 2007; Hill et al., 2000; Kato et al., 2000; Kuroiwa et al., 2002), and buffalo (Shi et al., 2007), a few live births have been sufficient to assess the effectiveness of the technique. The use of these two cell types to produce cloned embryos was unprecedented, and the results showed that MSC-AT were able to generate a healthy calf with an efficiency greater than that reported in the literature, therefore indicating that such cells can be used in NT.
MSCs represent an attractive cell source for therapeutic applications due to their potential for secreting bioactive molecules that are both trophic and immunomodulatory in nature (Caplan, 2009). A lot of research has been done with the aim of increasing the therapeutic applications of MSCs (Alviano et al, 2007; Chiavegato et al, 2007; De Coppi et al, 2007; Kunisaki et al, 2007), but our study is the first report of the use of MSC-AF and MSC-AT for the production of live calves by NT. One of the few studies using MSC-AF in NT was done by Zheng et al. (2009) in swine, resulting in an increase in the rate of blastocyst production, which leads us to believe that these cells are less differentiated and, therefore, more applicable in cloning. Furthermore, Colleoni et al. (2005) also used MSCs derived from bone marrow in NT and obtained a 63.7% of blastocyst rate, but that study did not describe either the pregnancy or calf cloned from this cell type.
Our study proved it is possible to recover viable cells by ultrasound-guided transvaginal amniocentesis in vivo and to grow them with efficiency, unlike other experiments performed from collection by cesarean section (Gao et al., 2014) and at the time of exposure of the amniotic membrane at the beginning of a normal birth (Corradetti et al., 2013). The use of this type of cells is an alternative to recloning, because these cells can be recovered from a cloned fetus during early pregnancy. Recloning is characterized by the removal of a cloned fetus on the fortieth day of pregnancy for the isolation of fetal fibroblasts, in order to use younger and lesser differentiated cells in NT. This procedure is extremely criticized because it runs contrary to ethical standards for animal handling. With the use of MSC-AF isolated from live animals, there is no need to remove the fetus surgically.
In this study, cells isolated from amniotic fluid and adipose tissue presented immunophenotyping patterns required for characterizing these cells as MSCs, when they were revealed to be positive for CD105, CD29, CD73, and CD90 MSC markers and negative for CD45 and CD34. Both CD34 and CD45 are typical markers of hematopoietic and endothelial cell lines. There is a consensus in the literature that MSCs do not have these markers, a fact confirmed in the present study. These markers could indicate contamination with clotted blood at the time of collection.
Unexpectedly, we observed that MSC-AT were positive for the CD44 marker. According to Monteiro et al. (2010), the molecular marker CD44 (hyaluronate receptor) was expressed on the surface of the MSCs isolated from bone marrow from humans and mice. Furthermore, pluripotency was confirmed by the ability of bovine MSC-AF and MSC-AT to differentiate into adipocytes, chondrocytes, and osteocytes. According to Baksh et al. (2004), differentiation is an intrinsic capacity of MSCs.
Hiiragi and Solter (2005) demonstrated that nuclear reprogramming is essential in NT when they performed NT with a zygote cell as nuclear donor and obtained high rates of live births compared to two- and four-cell-stage embryos as nuclear donors. According to these authors, the zygote is totipotent and requires no or minimal reprogramming after NT, resulting in the higher success rates observed in this treatment. In the same way, in our experiment, the high pregnancy rates observed may be explained by the use of MSCs.
MSC-AT were able to generate one live and healthy animal in this study, thus highlighting them as an important cellular source for NT. Some advantages, such as easy accessibility, collection by a noninvasive method, and the lack of limitations for sex or age of the animal (Kubota et al., 2000), previously attributed to fibroblasts, may also be associated with the MSC-AT.
The fusion rate may vary according to the size of the cell used. The larger cell size of MSC-AF compared to MSC-AT was found, which possibly contributed to an appropriate contact between the donor cell membrane, the zona pellucida, and the plasma membrane of the oocytes. In addition, the size of MSC-AF facilitated its visualization at the time of fusion, making the procedure faster and more efficient. This difference in size may explain the variation in rates of fusion between cell types. These results corroborate the findings of Galli et al. (2002), who reported that the cells must be intact and medium to large size, so that the membranes are fused.
The overall efficiency of the technique described in the literature is from 0% to 5% (Heyman et al., 2002). However, when we used MSC-AF and MSC-AT, the rate of offspring born was higher, 25% and 12.5%, respectively, than that described in the literature. The calf from NT with MSC-AF was born weighing 58.5 kg, clearly showing a characteristic of large offspring syndrome (LOS).
In the case of LOS, a reduced ability for the neonate to survive is due in part to the high incidence of dystocia in these cases, and related calf deformities can be explained by the lack of space in the uterus for a large and heavy fetus (Garry et al., 1996). According to Young et al. (1998), the occurrence of this syndrome in cattle and sheep occurs after exposure to unusual environments both in vivo and in vitro. Besides these factors, Campbell et al. (2007) argue that LOS can be the result of abnormal reprogramming of the donor nucleus during cloning procedures. Studies examining the methylation status of imprinted genes in embryos, fetuses, and placentas of mice and sheep were consistent with the hypothesis that epigenetic dysregulation of the expression of imprinted genes is a key factor in the occurrence of LOS (Farin et al., 2006).
Although the calf from the cloning with amniotic cells died soon after birth, this cell type seems to be a good alternative to recloning. One option is the use of drugs that act in methylation or acetylation of cells and embryos whenever using MSC-AF in the NT procedure. This combination might improve the rates of live and healthy calves.
The calf from NT with MSC-AT was born through assisted vaginal delivery, weighed 35 kg, and showed a slight thickening of the umbilical cord as the only change. The physiological pregnancy and delivery observed with the use of this cell type opens a new possibility of using it as a donor in NT.
In conclusion, this study demonstrated the usefulness of MSC-AT in NT. Furthermore, adipose tissue appeared to be a promising cell source of MSCs to use in NT because it generated a healthy calf. The growing use of cloning both commercially and in research requires an improvement in the efficiency of the technique, and the advance in studies with MSCs from amniotic fluid and adipose tissue is important for increasing the efficiency of NT in cattle.
Footnotes
Acknowledgments
The authors thank Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação de Apoio à Pesquisa do Distrito Federal (FAPDF)–PRONEX/FAPDF (193.000.048/2009), Conselho Nacional de Desenvolvimento Cientifico e Tecnológico–CNPq (30116/2009-1), Financiadora de Estudos e Projetos–Finep (CTInfra-01/2001), and Embrapa (MP1 01.13.06.001.06.02.004) for financial support.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.