
Abstract
Abstract
Transgenic animal producing technology has improved consistently over the last couple of decades. Among the available methods, somatic cell nuclear transfer (SCNT) technology was officially the most popular. However, SCNT has low efficiency and requires a highly skilled individual. Additionally, the allo-SCNT nuclear reprogramming mechanism is poorly understood in the gnotobiotic miniature pig, which is a candidate for xenotransplantation, making sampling in oocytes very difficult compared to commercial hybrid pigs. Therefore, interbreed SCNT (ibSCNT), which is a combination of miniature pig and commercial pig (Landrace based), was analyzed and was found to be similar to SCNT in terms of the rate of blastocyst formation (12.6% ± 2.9% vs. 15.5% ± 2.2%; p > 0.05). However, a significantly lower fusion rate was observed in the ibSCNT compared to normal SCNT with Landrace pig somatic cells (29.6% ± 0.8% vs. 65.0% ± 4.9%). Thus, the optimization of fusion parameters was necessary for efficient SCNT. Our results further revealed that ibSCNT by the whole-cell intracytoplasmic injection (WCICI) method had a significantly higher blastocyst forming efficiency than the electrofusion method (31.1 ± 8.5 vs. 15.5% ± 2.2%). The nuclear remodeling and the pattern of changes in acetylation at H3K9 residue were similar in both SCNT and ibSCNT embryos.
Introduction
G
In this study, we describe the efficiency of ibSCNT using commercial pig oocytes and somatic cells from gnotobiotic miniature pigs at our Bio-Organ Research Center (BORC). We report that optimization of fusion parameters is necessary to obtain satisfactory fusion of miniature pig donor cells with Landrace oocytes. Alternatively, the whole-cell intracytoplasmic injection (WCICI) method of nuclear transfer can be used to obtain better results.
Methods and Materials
Animals
Gnotobiotic miniature pig located in animal facilities at the BORC, Konkuk University (Seoul, Korea).
Oocyte retrieval and in vitro maturation
Prepubertal pig ovaries were collected from a local slaughterhouse and transported to the laboratory in saline maintained at 35°C–38°C. Cumulus oocyte complexes (COC) were aspirated from follicles (26 mm diameter) using 18G needle syringes. The COC were washed twice in TL-HEPES medium containing 1 mg/mL bovine serum albumin (BSA) and matured in Tissue Culture Medium 199 with Earle's salt (TCM199; Gibco BRL) supplemented with 25 mM NaHCO3, 10% (v/v) pig follicular fluid, 0.57 mM cysteine, 0.22 μL/mL sodium pyruvate, 25 μg/mL gentamycin sulfate, 0.5 uL/mL p-FSH (Folltropin V; Vetrepharm), 1 μg/mL estradiol-17B, and 10 ng/mL epidermal growth factor under mineral oil at 39.8°C in a humidified atmosphere of 5% CO2 in air for 42–44 hours, as described earlier (Gupta et al., 2006).
Somatic cell nuclear transfer
Preparation of donor cells
Donor cells for nuclear transfer were prepared essentially as described earlier (Gupta et al., 2007). Briefly, primary fetal fibroblasts were isolated from the ear of newly born piglets and were cultured on 60-mm tissue culture dishes (Falcon BD) in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL) supplemented with 10% (v/v) fetal bovine serum (FBS; Gibco BRL). After 7 days of culture, cells were trypsinized and resuspended in DMEM supplemented with 10% (v/v) FBS. Cells were routinely maintained on 50-mL tissue culture flasks (Falcon BD) up to two to seven passages and were used as donor cells for nuclear transfer after attainment of confluence to synchronize their cell cycle stage to G0/G1.
Enucleation and nuclear transfer
Enucleation and nuclear transfer were performed as described earlier (Gupta et al., 2007) with partial modifications. Briefly, in vitro matured oocytes were stripped of cumulus cells in TL-HEPES supplemented with 0.1% (w/v) hyaluronidase and washed three times in TL-HEPES containing 0.1% (w/v) BSA. Denuded oocytes were incubated for 20 minutes in the HEPES-buffered TCM199 (HTCM; Gibco BRL) medium supplemented with 10% (v/v) FBS, 7.5 μg/mL cytochalasin B (CB), and 5 μg/mL Hoechst 33342. Enucleation was then performed by aspirating the first polar body and adjacent cytoplasm (∼30% of ooplasm) using a beveled pipette (∼25 μm internal diameter) in the HTCM medium. Successful enucleation was confirmed by ultraviolet-assisted visualization of fluorescent metaphasic plate in the aspirated cytoplasm contained within the enucleation pipette. Enucleated oocytes were subsequently reconstructed by inserting a small-sized (∼15 μm in diameter), smooth bordered donor cell into the perivitelline space of each enucleated oocyte using the same pipette used for enucleation. Reconstructed oocytes were placed in the TCM199 medium added with lectin (10 μg/mL) for 30 minutes to allow recovery. Fusion was achieved at room temperature in a chamber with two platinum electrodes at 1 mm apart overlaid with fusion solution that was composed of 0.3 M mannitol, 0.1 mM CaCl2, and 0.1 mM MgSO4. The reconstructed oocytes were aligned manually with a fine Pasteur pipette, so that the contact surface between the donor cell and oocyte was parallel to the electrodes. Membrane fusion was induced with single DC pulse of 2.1 kV/cm for 30 μseconds delivered by a BTX Electro Cell Manipulator 2001 (BTX). This pulse was also utilized to simultaneously induce activation of the reconstructed embryos. Following activation, activated oocytes were cultured in the North Carolina State University Medium 23 (NCSU23) supplemented with 7.5 μg/mL CB for 4 hours before transferring to an in vitro culture (IVC) medium for further culture. In case of WCICI method of SCNT, fusion step was eliminated; instead, cells were directly injected into the enucleated cytoplast using a new injection pipette (∼8 to 10 μm in diameter), and reconstructed oocytes were allowed to reprogram the injected donor cell for 1, 2, or 4 hours before progressing to the activation step (Das et al., 2010).
IVC of embryos
Embryos were cultured in the NCSU23 medium supplemented with 0.4% essentially fatty acid-free BSA for 5 days and in the NCSU23 medium supplemented with 10% (v/v) FBS for subsequent 2 days as described earlier (Gupta et al., 2007). The cleavage rate was assessed on day 2 and the blastocyst rate on day 7 of culture. At 168 hours of IVC, blastocysts were harvested for assessment of embryo quality by Hoechst 33342 staining.
Fluorescent staining for assessment of nuclear remodeling
Changes in chromatin configuration of cloned embryos were observed by staining them, at 3, 6, 12, and 24 hours after nuclear transfer, with the Hoechst 33342 stain as described earlier (Uhm et al., 2007). Briefly, embryos were fixed for 5 minutes in fixative solution containing 2% formalin and 0.25% glutaraldehyde. Fixed embryos were then mounted on clean glass slides and stained with a glycerol-based Hoechst 33342 (12.5 μg/mL) staining solution for 10 minutes. Embryos were examined under an epifluorescence microscope at ×400 magnification and designated as premature chromatic condensation (PCC), nuclear envelope breakdown (NEBD), pseudopronucleus (pseudo-PN), nuclear swelling (NS), or cleaved embryos according to the morphological characteristics of the nucleus.
Immunocytochemistry for assessment of acetylation at H3K9
Immunocytochemistry for analysis of acetylation at ninth Lysine residue of Histone 3 (Ac-H3K9) was carried out as described earlier (Yeo et al., 2005). Briefly, embryos were removed of zona pellucida using Acid Tyrode's solution, washed twice in phosphate buffered saline (PBS)-BSA, and fixed at 4°C for 30 minutes in PBS containing 4% (w/v) paraformaldehyde. The fixed embryos were then washed three times with PBS-BSA and permeabilized with 0.5% (v/v) Triton-X 100 in PBS-BSA for 1 hour, following which they were incubated with rabbit anti-acetyl-H3-K9 (1:100; Upstate Biotechnology) at 39°C for 1 hour. After being washed six times in PBS-BSA-0.05% Tween 20 for 10 minutes each, the oocytes were incubated with FITC-labeled anti-rabbit IgG (1:1000 dilution; Molecular Probes) for 30 minutes at 39°C. The embryos were then mounted on slides and stained with 300 nM 4′,6′-diamidino-2-phenylindole and observed using a fluorescent microscope with Apotome (Axioplan; Carl Zeiss). Images were captured digitally using appropriate filter sets and processed using software (Axiovert 200; Carl Zeiss).
Statistical analyses
Statistical analyses were carried out using SAS software (Statistical Analysis System, Inc.) for chi-square test and analysis of variance where appropriate. The percentage data were subjected to arc sine transformation before statistical analyses. Data are presented as mean ± standard error of the mean. Differences at p ≤ 0.05 were considered significant.
Results
In vitro development of ibSCNT embryos
Cloned miniature pig embryos were successfully produced by electrofusion of miniature pig fibroblasts into commercial Landrace pig oocytes by ibSCNT. The ibSCNT embryos had a similar (p > 0.05) ability to cleave (71.4 ± 2.6 and 68.8% ± 4.4%) and form blastocysts (15.5 ± 2.2 and 12.6% ± 2.9%) as was obtained with electrofusion of Landrace fibroblasts into Landrace oocytes by SCNT (Table 1). The SCNT and ibSCNT blastocysts also contained a similar number of nuclei/blastocysts (43.1 ± 9.0 and 43.8% ± 9.3%; p > 0.05). However, the miniature pig fibroblasts fused with Landrace oocytes at a lower (p < 0.05) frequency than observed with Landrace pig fibroblasts (29.6 ± 0.8 vs. 65.0% ± 4.9%) and required the optimization of fusion parameters. An optimal fusion rate (80.3%) was obtained with a BTX 2001 pulse generator in HV mode using a single DC pulse of 1.8 kV/cm voltage for 30 μseconds (Table 2).
Values with different superscripts (a, b, c) within column denote a difference (p < 0.05).
Values within parenthesis indicate the number of embryos.
ibSCNT, interbreed SCNT; SCNT, somatic cell nuclear transfer.
BLS, BLS CF-150/B (Biological Laboratory Equipments); BTX, BTX ECM 2001 (Harward Apparatus). Fusion chamber description: BTX model 450–1, Microslide 1 mm, Catalog No. 45-0104. Value within parenthesis indicates the number of oocytes.
In vitro development of ibSCNT embryos
Enucleated Landrace oocytes could also direct the embryonic development of miniature pig fibroblasts after WCICI. Interestingly, the blastocyst rate obtained with WCICI was significantly higher (31.1% ± 8.5%) than those observed with electrofusion (15.5% ± 2.2%) of ibSCNT (p < 0.05) (Table 3). There was no significant difference between the two groups in the rate of cleavage (71.4 ± 2.6 vs. 76.9% ± 8.3%; p > 0.05). However, the in vitro development ability of the WCICI embryos was dependent on the time interval between whole-cell injection and oocyte activation (Table 2). The best development was observed when activation was applied at 2 hours postcell injection.
Values within parenthesis indicate the number of embryos.
Values with different superscripts (a, b) within column denote a difference (p < 0.05).
EF, electrofusion; WCICI, whole-cell intracytoplasmic injection.
Nuclear remodeling following ibSCNT
The reconstructed embryos produced by nuclear transfer of miniature pig donor cells into Landrace pig oocytes were examined for changes in their chromatic configuration at 3, 6, 12, and 24 hours after nuclear transfer. NEBD and PCC were observed within 3 hours of nuclear transfer. This was followed by NS and pseudo-pronuclear formation. Occasionally, the reconstructed oocytes showed a cleavage division when observed at 24 hours after nuclear transfer. The proportion of oocytes in different chromatic configuration during different time duration is shown in Table 4.
Values within parenthesis indicate the number of reconstructed embryos.
Dividing cells; NEBD, nuclear envelop breakdown; PCC, premature chromatic condensation; Swelling, nuclear swelling; PN, one pseudo-pronucleus.
Changes in histone acetylation at H3K9 residue following ibSCNT
The changes in Ac-H3K9 following ibSCNT of miniature pig donor cells into the Landrace pig oocytes is shown in Figure 1. The Ac-H3K9 signal rapidly disappeared with the occurrence of PCC and started reappearing with the NS. Ac-H3K9 was fully recovered with the formation of pseudo-PN. The pattern of changes in Ac-H3K9 was similar in both SCNT (Fig. 1A) and ibSCNT embryos (Fig. 1B).
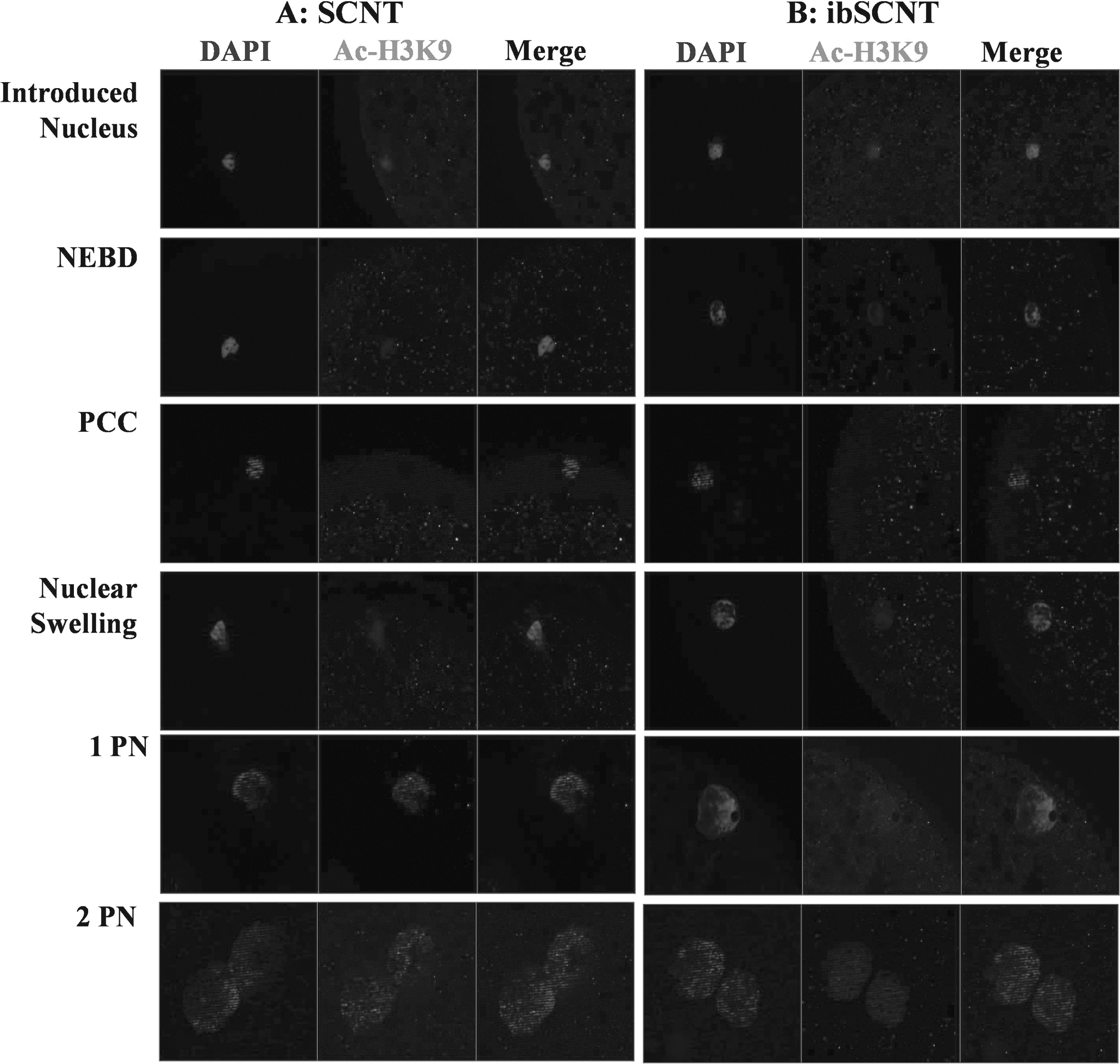
Changes in histone acetylation at H3K9 residue (Ac-H3K9) following nuclear transfer of Landrace pig (
Discussion
Generation of human organs for xenotransplantation requires the establishment of a system for the isolation and rearing of gnotobiotic piglets in an absolute barrier-sustained facility and the development of SCNT technology for gene targeting (Garkavenko et al., 2008; Kolber-Simonds et al., 2004). We previously described the production, rearing, and microbiological monitoring of gnotobiotic miniature pigs at our BORC (Hwang et al., 2015). In this study, we describe a method for producing cloned miniature pig embryos by ibSCNT using commercial Landrace pig oocytes.
We observed that commercial Landrace oocytes could successfully reprogram the somatic nuclei of miniature pigs to direct embryonic development at least up to the blastocyst stage. However, the electrofusion ability of miniature pig cells with Landrace oocytes was lower than that obtained with Landrace pig cells. Consequently, a large number of karyoplast–cytoplast couplets were lost. The cause for this lower fusion efficiency of miniature pig donor cells with Landrace pig oocytes is unclear, but this difference might be due to differences in the surface structure of the plasma membrane of minipig and commercial Landrace pig donor cells (Stachowiak et al., 2007). This seemed likely because changes in fusion parameters (amplitude, width, pulse, and type of electrical voltage settings) could improve the fusion efficiency without compromising their developmental ability.
To overcome this problem, we attempted direct whole-cell microinjection into the Landrace oocyte using the WCICI method originally developed by Lee et al. (2003b) and modified by Das et al. (2010). Lee et al. (2003b) observed that the plasma membrane of somatic cells can undergo dissolution within the ooplasm and that the nucleus could be effectively reprogrammed to a full-term piglet after WCICI. Consistent with previous reports, we observed that enucleated Landrace oocytes could direct the embryonic development of miniature pig fibroblasts upon WCICI and thereby can provide an efficient and less labor-intensive protocol of ibSCNT. This is consistent with those reported in goat SCNT (Chen et al., 2007), pig SCNT (Das et al., 2010; Lee et al., 2003a), and panda-rabbit interspecies SCNT (Jiang et al., 2004). Interestingly, the rate of blastocyst formation was dramatically higher in the WCICI method than in the electrofusion method. The efficiency of blastocyst production was increased substantially when the time from injection to activation increased from 0.0 to 1–2 hours. Improvement in blastocyst rate by increasing the time between injection and activation has been reported in pig (Lee et al., 2003b) through SCNT and in yak embryos through ibSCNT (Li et al., 2007). The highest rate of blastocyst formation was observed when activation was performed at 2 hours postcell injection. This improvement may be attributed to maturation promoting factor (MPF) activity in the recipient oocytes after donor cell injection. MPF activity is maximal in MII oocytes and exposure of the donor genome to high levels of MPF has been reported to be necessary to reprogram a donor nucleus from a differentiated cell. Therefore, the delay of 2 hours after injection may have allowed the donor genome to be exposed to high MPF levels in the oocyte cytoplasm and therefore may have improved the development of cloned embryos into blastocysts (Lee et al., 2003b). More importantly, the use of WCICI completely eliminated the loss of karyoplast–cytoplast couplets due to the failure of electrofusion and therefore increased the total number of blastocysts formed from a given number of oocytes and donor cells used for SCNT. However, the WCICI method requires more skill as a large proportion (∼20%) of oocytes is lysed during the manipulation procedure.
The remodeling and reprogramming of the donor cell nucleus within the cytoplasm of the recipient oocyte to organize the first embryonic division are vital for the success of SCNT (Yin et al., 2003). Therefore, the dynamics of nuclear remodeling events following nuclear transfer of minipig donor cell into commercial Landrace pig recipient cytoplast was examined in the present study. We observed that NEBD and PCC occurred within 3 hours of ibSCNT, while ∼45% and 41% of reconstructed embryos showed NS and pseudo-pronuclear formation within 6 hours of ibSCNT, respectively. These data therefore demonstrated that the Landrace pig recipient cytoplast could remodel the nucleus of miniature pig donor cells. Results further showed that the miniature pig donor nucleus rapidly deacetylated following nuclear transfer into Landrace pig oocytes and remained deacetylated during NEBD and PCC. Re-establishment of acetylation occurred with formation of pseudo-PN formation. This suggests that commercial Landrace pig cytoplast could efficiently reprogram the minipig donor nucleus and supports the hypothesis that factors existing in ooplasm for somatic nucleus reprogramming may be conserved (Chen et al., 2006; Liu et al., 2004).
In summary, cloned miniature pig embryos can be successfully obtained by ibSCNT into Landrace pig oocytes using both electrofusion and WCICI. A reprogramming timing of 2 hours may improve the cloning efficiency with the WCICI method.
Footnotes
Acknowledgment
This work was carried out with the support of the “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ01119603),” Rural Development Administration, Republic of Korea, and written as part of Konkuk University's research support program for its faculty on sabbatical leave in 2014.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.