
Abstract
Abstract
Freshly ovulated rat eggs do not remain arrested at metaphase II (MII) and undergo exit from MII arrest with initiation of extrusion of the second polar body (PBII), a characteristic feature of abortive spontaneous egg activation (SEA). The biochemical and molecular changes during postovulatory aging-mediated abortive SEA remain poorly understood. We investigated the morphological, cellular, and molecular changes during postovulatory aging-mediated abortive SEA in eggs cultured in vitro. Our results suggest that postovulatory egg aging in vitro induced initiation of PBII extrusion in a time-dependent manner. Postovulatory aging increased Wee1 kinase and Thr-14/Tyr-15 phosphorylated cyclin-dependent kinase 1 (Cdk1) levels, whereas Thr-161 phosphorylated Cdk1 and cyclin B1 levels were significantly decreased in eggs cultured in vitro. The early mitotic inhibitor 2 (Emi2) level was significantly reduced, but anaphase promoting complex/cyclosome (APC/C) and mitotic arrest deficient protein (MAD2) levels were increased initially and then reduced during a later period of in vitro culture. These results suggest that an increased Wee1 kinase level modulated the specific phosphorylation status of Cdk1, increased Cdk1 activity, and decreased the cyclin B1 level. Furthermore, the decreased Emi2 level was associated with an increased level of APC/C and decreased level of cyclin B1, which resulted in maturation promoting factor (MPF) destabilization and finally led to postovulatory aging-mediated abortive SEA in rat eggs cultured in vitro.
Introduction
I
Postovulatory aging-mediated abortive SEA in rat eggs was reported for the first time by Keefer and Schuetz in 1982 and later by Zernicka-Goetz in 1991 (Keefer and Schuetz, 1982; Zernicka-Goetz, 1991). Thereafter, it was reported in several mammalian species, including mice (Abbott et al., 1998; Lord et al., 2013), rat (Chaube et al., 2007; Premkumar and Chaube, 2013, 2014; Prasad et al., 2015b), hamster (Juetten and Bavister, 1983), porcine (Ito et al., 2003, 2004; Ruddock et al., 2001), bovine (Sergeev and Norman, 2003), and human eggs (Combelles et al., 2011; Escrich et al., 2011; Lu et al., 2006). Recent studies from our laboratory suggest that postovulatory aging-mediated abortive SEA shows initiation of PBII extrusion that is never completed and further arrest at the metaphase III- (MIII) like stage due to scattering of chromosomes throughout the egg cytoplasm (Premkumar and Chaube, 2013, 2014).
Biochemical and molecular changes during postovulatory aging-mediated abortive SEA remain poorly understood. Few studies suggest that insufficient release of calcium (Ca2+) from internal stores, like the endoplasmic reticulum (ER), through ryanodine receptor (RyR) channels thereby increases cytosolic-free Ca2+ and is involved during postovulatory aging-mediated abortive SEA (Premkumar and Chaube, 2013, 2014, 2015). The aged eggs are unable to handle such an increased cytosolic free Ca2+ level, and maturation promoting factor (MPF) destabilization is triggered (Prasad et al., 2015b; Premkumar and Chaube, 2013, 2014). Wee1 destabilizes MPF by increasing the Thr-14/Tyr-15 phosphorylated cyclin-dependent kinase 1 (Cdk1) level and decreasing Thr-161 phosphorylated Cdk1 as well as cyclin B1 levels (Kubiak et al., 2008; Oh et al., 2011; Prasad et al., 2015b). However, MPF stabilization requires reduced phosphorylation at Thr-14/Tyr-15 of Cdk1, increased phosphorylation at Thr-161 of Cdk1, and association with cyclin B1 (Kubiak et al., 2008; Prasad et al., 2015a, b). The stabilized MPF is required for the maintenance of MII arrest in ovulated eggs of several mammalian species (Madgwick and Jones, 2007).
The maintenance of MII arrest is also achieved due to the presence of cytostatic factors (CSFs), such as early mitotic inhibitor 2 (Emi2), anaphase promoting complex/cyclosome (APC/C), and mitotic arrest deficient protein (MAD2) in the cytoplasm of eggs (Kubiak et al., 2008; Madgwick and Jones, 2007). Emi2 maintains MII arrest by inhibiting APC/C activity (Schmidt et al., 2005). On the other hand, APC/C degrades cyclin B1 through ubiquitin-mediated proteolysis that destabilizes MPF (Kubiak et al., 2008; Nabti et al., 2008; Rauh et al., 2005; Schmidt et al., 2005). Cyclin B1 degradation as well as inactivation of MAD2 result in meiotic exit from diplotene arrest and progression to the MII stage (Homer et al., 2005; Zhang et al., 2004). Although MPF destabilization/stabilization plays a major role in meiotic exit from MII arrest (Oh et al., 2011, 2013; Premkumar and Chaube, 2013, 2014, 2015), studies suggest that specific phosphorylation of Cdk1 and involvement of Cdk1 activity associate with meiotic resumption from diplotene arrest (Davydenko et al., 2013; Jang et al., 2014; Pandey et al., 2014). However, it remains unclear whether the specific phosphorylation status of Cdk1 as well as cyclin B1 level and Cdk1 activity are involved during postovulatory aging-mediated abortive SEA. Therefore, the present study was aimed at discovering the time-dependent changes in Emi2, APC/C, MAD2, Wee1, the total and specific phosphorylation status of Cdk1 and cyclin B1 levels, and Cdk1 activity during postovulatory aging-mediated abortive SEA in rat eggs cultured in vitro.
Materials and Methods
Chemicals and preparation of culture medium
Chemicals used in this study were purchased from Sigma Chemical Co. (St. Louis, MO, USA) unless stated otherwise. Culture medium-199 (M-199; HiMedia Laboratories, Mumbai, India) was prepared as per the manufacturer's protocol. The pH was adjusted to 7.2 ± 0.05 and osmolarity was 290 ± 5 mOsmol. Antibiotics (
Superovulation induction and collection of MII arrested eggs
Female rats (22–25 days old; 50 ± 5 grams, sexually immature) of Charles-Foster (CF) strain were housed in an air-conditioned, light-controlled room with food and water ad libitum and subjected to the superovulation induction protocol [20 IU pregnant mare serum gonadotropin (PMSG) for 48 h followed by 20 IU human chorionic gonadotropin (hCG) for 14 h]. Experimental animals were killed after mild ether anesthesia. The ovaries along with fallopian tubes were collected in prewarmed culture medium. The ampulla of the fallopian tube was punctured using a 26-gauge needle and ovulated cumulus–oocyte complexes (COCs) were collected in culture medium. COCs were exposed to 0.01% hyaluronidase for 3 min at 37°C followed by repeated manual pipetting to remove the encircling cumulus cells. Denuded eggs were washed three times with culture medium, and eggs arrested at the MII stage possessing PBI were used for the entire in vitro studies. The experiment was repeated three times to confirm our results. This project was approved by Animal Ethical Committee of the University vide letter No. Dean/11-12/CAEC/266.
Analyses of morphological and chromosomal status during postovulatory egg aging in vitro
To induce egg aging in vitro, a group of 12–14 MII arrested eggs was transferred to a petri dish containing 2 mL of medium and cultured for various times (1, 2, 3, 5, and 7 h) in a CO2 incubator (Galaxy 170R incubator, Eppendorf, New Brunswick, USA) at 37°C. At the end of each incubation period, eggs were removed, washed, and observed for in vitro aging-mediated morphological changes using a light microscope (model E200, Nikon Eclipse, Tokyo, Japan). Three independent experiments were conducted to collect eggs sufficient for biochemical and immunofluorescence analysis. The meiotic status of aging eggs was confirmed by analyzing the chromosomal status using Hoechst 33342. For this purpose, 10–12 eggs were incubated with Hoechst 33342 (10 μg/mL) for 10 min at 37°C. Thereafter, eggs were removed and washed 10 times with phosphate-buffered saline (PBS). Eggs were then observed under a fluorescence microscope (Model Ni-U, Nikon Eclipse) at 350 nm for the analyses of chromosomal status at 400× magnification.
Analyses of Emi2, APC/C, Wee 1, MAD2, Thr-161 phosphorylated Cdk1, Thr-14/Tyr-15 phosphorylated Cdk1, total Cdk1, and cyclin B1 levels
To analyze Emi2, APC/C, Wee 1, MAD2, Thr-161 phosphorylated Cdk1, Thr-14/Tyr-15 phosphorylated Cdk1, total Cdk1, and cyclin B1 levels, eggs were exposed to their specific antibodies, i.e., anti-pThr-161(cat. no. sc-12341), anti- pThr-14/Tyr-15 (sc-12340), cdc2 p34 PSTAIRE antibody (sc-53), anti-cyclin B1 antibody (sc-752), anti-Wee1 antibody (sc-9037), anti-Emi2 antibody (sc-68894), anti-APC/C antibody (sc-896), anti-MAD2 antibody (sc-6329), all purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Immunofluorescence analysis was carried out following our previously published protocol (Prasad et al., 2015b). In brief, 12–14 eggs from each group were fixed with 4% buffered formaldehyde. Air-dried slides were washed with PBS and permeabilized using Triton X-100 (0.01% in PBS) for 10 min at 37°C. Blocking of the nonspecific sites was done using blocking buffer [2.5% PBS and bovine serum albumin (BSA) solution] at 37°C for 30 min. After that, slides were exposed to their specific primary antibodies (1:500 dilution in blocking buffer; 100 μL) at 37°C for 2 h. Slides were washed three times (5 min each wash) with PBS and exposed to 100 μL of secondary antibody [fluorescein isothiocyanate (FITC)-labeled or tetramethylrhodamine (TRITC)-labeled; 1:1000 dilutions in blocking buffer] for 1 h at 37°C in a CO2 incubator.
At the end of incubation, slides were washed five times with PBS and mounted with fluorescence mounting medium (VECTASHIELD Antifade Mounting Medium, Vector Laboratories, Inc., Burlingame, CA, USA) to prevent photobleaching and observed under a fluorescence microscope (Model Ni-U, Nikon Eclipse) at 465 nm for FITC and 540 nm for TRITC at 400× magnification. Three independent experiments were conducted to confirm our observations, and representative photographs are shown in the Results section below. A total of 36–42 eggs were used for corrected total cell fluorescence (CTCF) analysis. All parameters for each egg were kept constant, and the entire area was selected for the analysis of immunofluorescence intensity using ImageJ software.
Analysis of Cdk1 activity by enzyme-linked immunosorbent assay
The Cdk1 activity in egg lysate was analyzed using a MESACUP Cdc2/Cdk1 kinase assay kit (MBL International Corporation, Nagoya, Japan) following the manufacturer's protocol. In brief, 50 eggs from each group were transferred to a microcentrifuge tube containing 50 μL of hypotonic lysis buffer (5 mM Tris, 20 mM EDTA, 0.5% TritonX-100, pH 8) for 1 h on ice. The lysate was centrifuged at 10,000 × g at 4°C for 15 min, and the clear supernatant was used for the quantitative estimation of Cdc2/Cdk1 kinase activity. In brief, 2.5 μL of the sample or sample buffer was added to the appropriate microcentrifuge tube and 2.5 μL of 10× cdc2 reaction buffer was added. Next, 2.5 μL of biotinylated MV peptide and 15 μL of distilled water were added to each microcentrifuge tube; 2.5 μL of 1 mM adenosine triphosphate (ATP) solution was added to each microcentrifuge tube and incubated for 30 min at 30°C. The reaction was terminated by adding stop reagent and centrifuged at 14,000 ×g for 15 sec.
The reaction mixture (100 μL) was transferred to each microwell coated with monoclonal antibody and incubated for 1 h at 25°C. The microwell was washed four times with wash buffer and 100 μL of streptavidin-POD conjugate was added in each well. After 30 min of incubation at 25°C, the microplate was washed four times with wash buffer and incubated for 5 min with substrate solution (100 μL) provided with the MESACUP Cdc2/Cdk1 kinase assay kit (MBL International Corporation, Nagoya, Japan). Finally, 100 μL of stop solution was added to each well, and the microplate was read at 492 nm within 10 min using a microplate reader (ECIL, Hyderabad, India). Samples from three independent experiments were run in one assay to avoid inter-assay and intra-assay variation, which was found to be 1.9%.
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM) of three independent experiments. All percentage data were subjected to arcsine square-root transformation before statistical analysis. Data were analyzed by analysis of variance (ANOVA) followed by Bonferroni post hoc analysis (v. 17.0, SPSS, Inc., Chicago, IL, USA). A probability of p < 0.05 was considered significant.
Results
Postovulatory aging induces PBII extrusion
Figure 1 shows postovulatory aging-mediated abortive SEA from MII arrest. Freshly ovulated eggs were arrested at the MII stage and possess PBI (Fig. 1A). Culture of these eggs in medium resulted in initiation of the extrusion of PBII (Fig. 1B) in a time-dependent manner (one-way ANOVA, F = 719.306, p < 0.001; Fig. 1E). After 7 h of in vitro culture, almost 40% of eggs showed complete extrusion of PBII (Fig. 1C), whereas remaining eggs showed incomplete extrusion of PBII (Fig. 1B). The postovulatory aging-mediated morphological stages were further confirmed by chromosomal status in the present study (Fig. 1D). The presence of a haploid set of chromosomes (Fig. 1D1, grey arrow; yellow arrow in color image) in the egg cytoplasm and another haploid set of chromosomes in PBI (Fig. 1D1, white arrow) confirms the MII stage. The postovulatory aging-mediated abortive SEA was confirmed by the presence of a haploid set of chromosomes inside the first polar body (Fig. 1D2, white arrow) and movement of the chromosomes from the metaphase plate (Fig. 1D2, white arrow head; red arrow in color image). SEA was confirmed by the presence of a haploid set of chromosomes in both polar bodies (PBI, Fig. 1D3, grey arrow; yellow arrow in color; PBII, Fig. 1D3, white arrow) as well as the presence of haploid chromosomes in the egg cytoplasm (Fig. 1D3, white arrow head; red arrow in color).
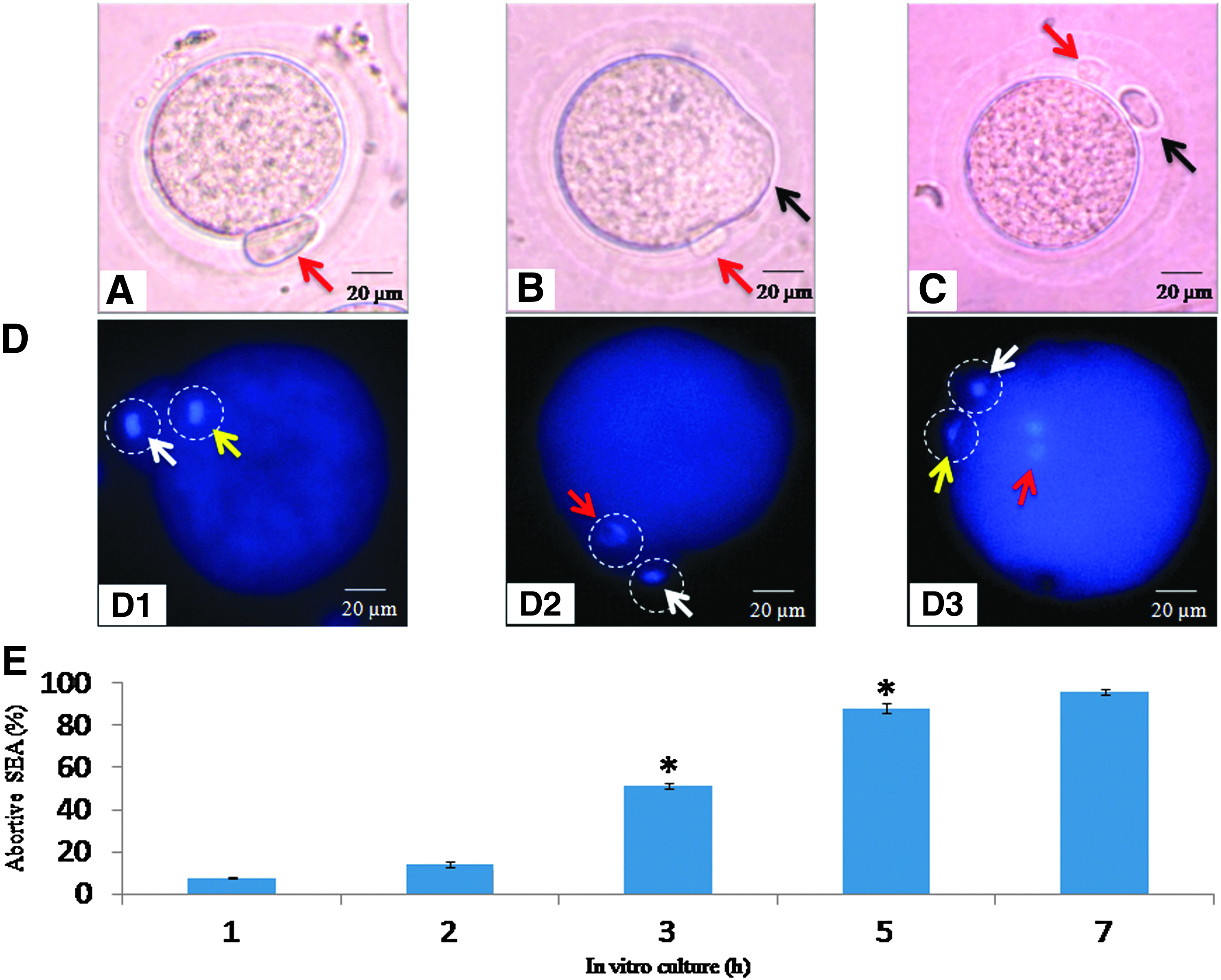
Representative photographs showing morphological changes during egg aging in vitro. (
Emi2 level decreases during postovulatory egg aging in vitro
As shown in Figure 2, postovulatory aging reduced immunofluorescence intensity of Emi2 (Fig. 2A2–A6) as compared to control eggs (Fig. 2A1). The reduced level of Emi2 was maintained until 7 h of in vitro culture (Fig. 2A2–A6) as evidenced by CTCF analysis of immunofluorescence intensity (one-way ANOVA, F = 372.196, p < 0.001; Fig. 2B).
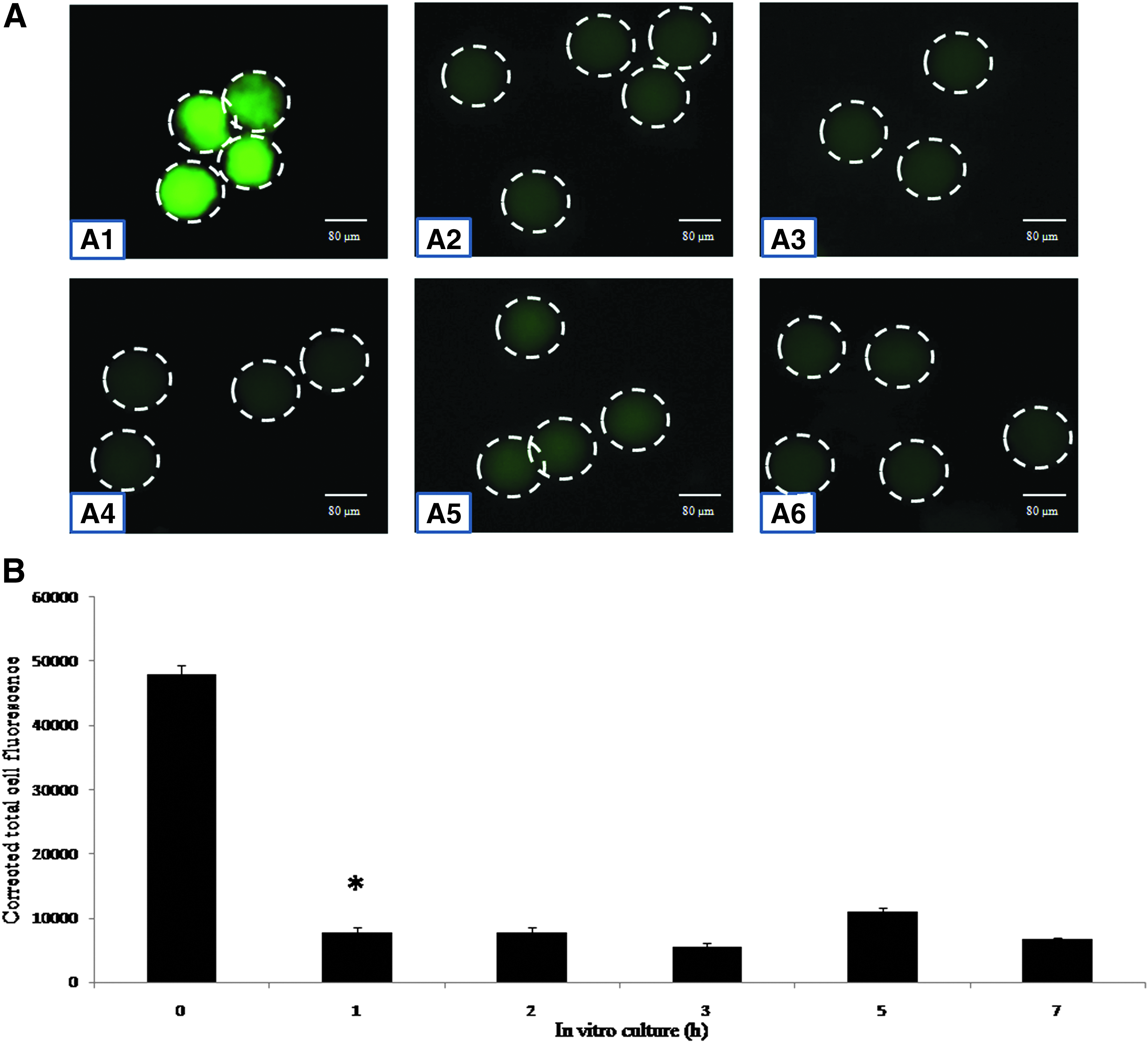
(
APC/C level first increases and then decreases during egg aging in vitro
Figure 3 shows postovulatory aging-mediated changes in the immunofluorescence intensity of APC/C in eggs cultured in vitro. The postovulatory aging increased immunofluorescence intensity of APC/C initially (Fig. 3A1–A3) and then decreased after 2 h of in vitro culture (Fig. 3A4–A6). The CTCF analysis of fluorescence intensity (Fig. 3B) further suggests a time-dependent (one-way ANOVA, F = 369.584, p < 0.001; Fig. 3B) increase of APC/C during the initial period of in vitro aging (2 h) and a decline thereafter in a time-dependent manner (one-way ANOVA, F = 202.796, p < 0.001; Fig. 3B).
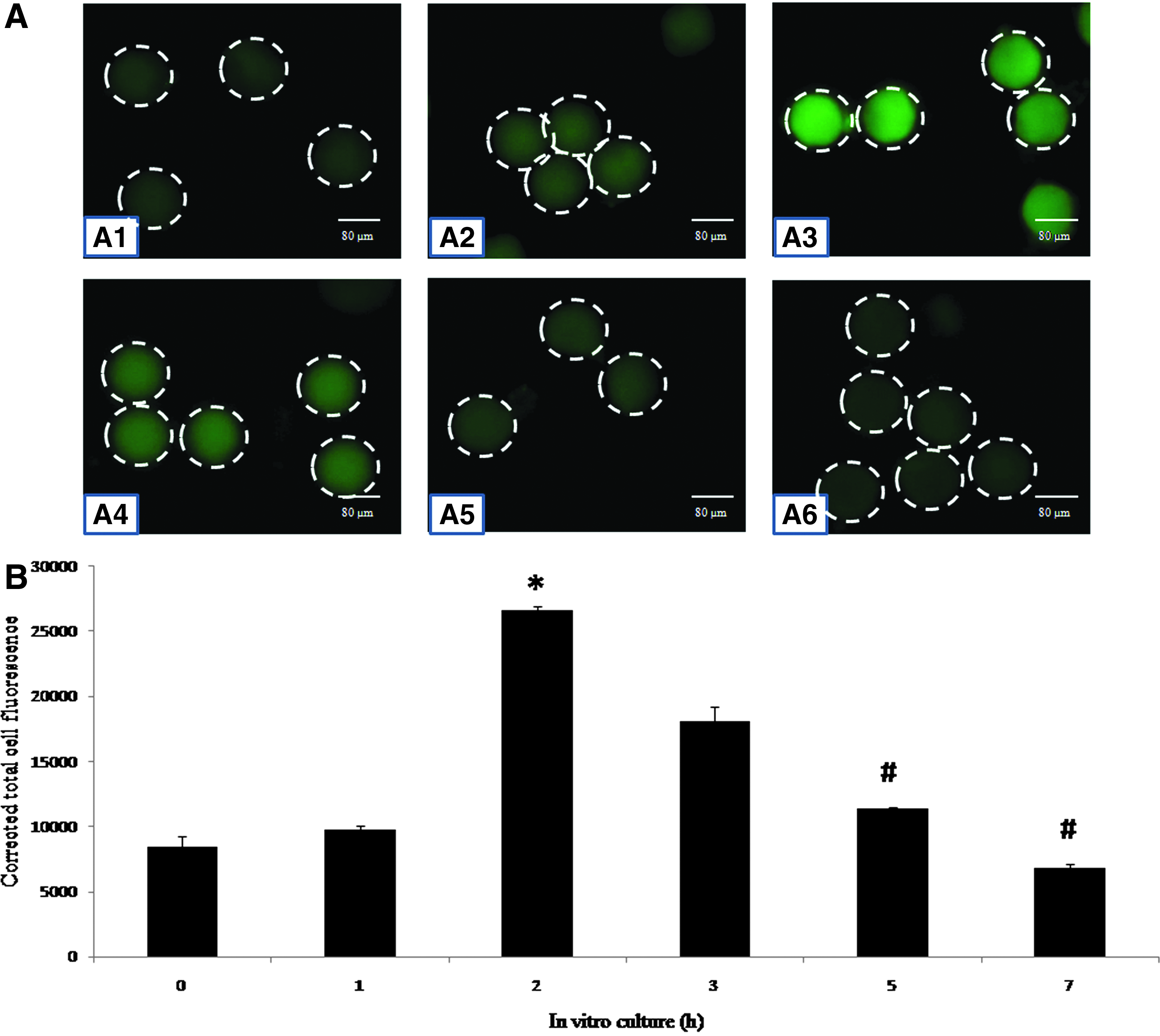
(
MAD2 level first increases and then decreases during egg aging in vitro
Figure 4 shows immunofluorescence intensity of MAD2 in eggs cultured in vitro. Postovulatory aging increased immunofluorescence intensity of MAD2 during the initial period (up to 2 h; Fig. 4A1–A3) and then declined gradually after 7 h of in vitro culture (Fig. 4A3–A6). CTCF analysis of immunofluorescence intensity further suggested a time-dependent increase of MAD2 (one-way ANOVA, F = 20.819, p < 0.05; Fig. 4B) during the initial period and a gradual decline during the later period of in vitro culture (one-way ANOVA, F = 13.84, p < 0.05; Fig. 4B).
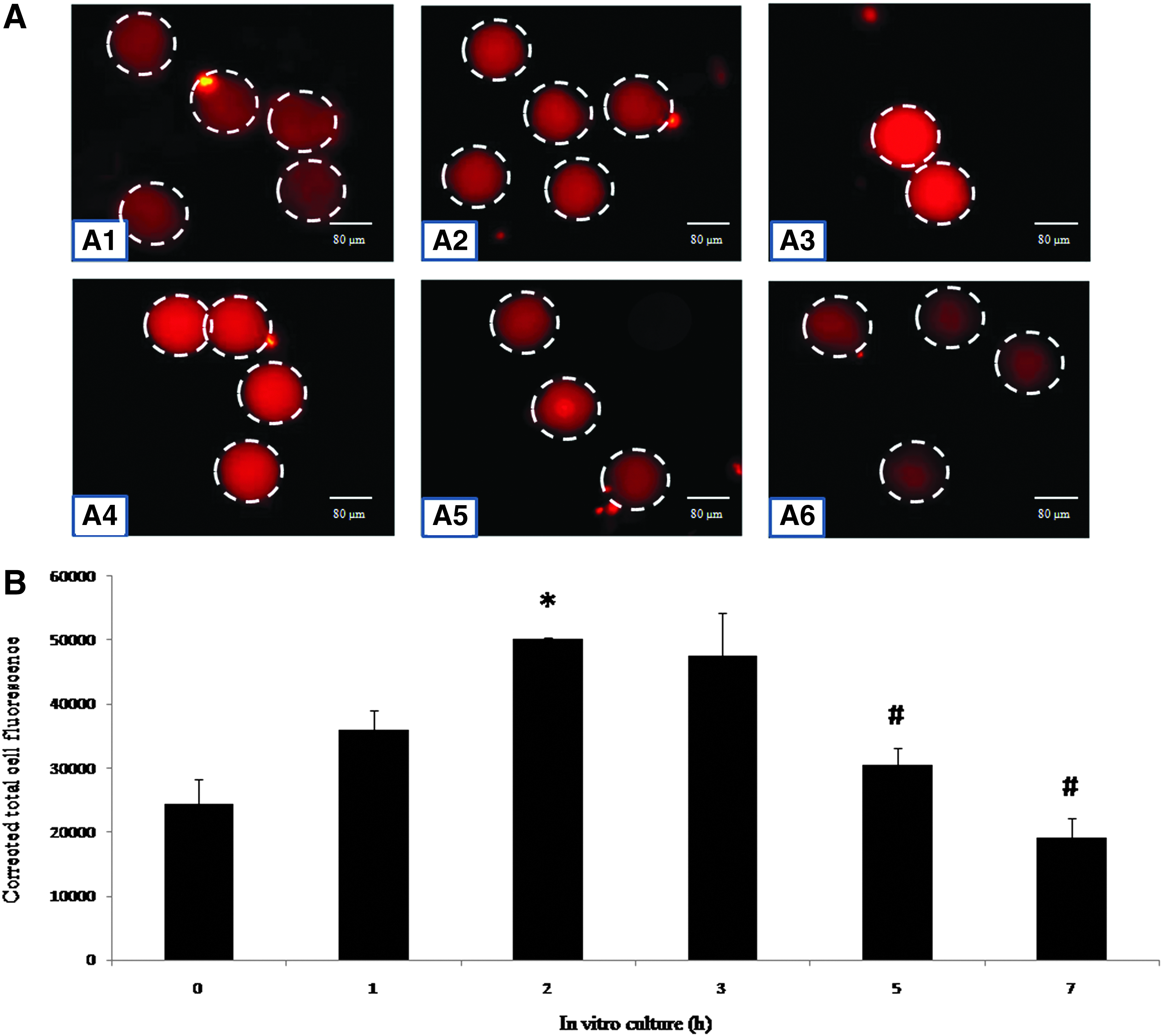
(
Cyclin B1 decreases during egg aging in vitro
Figure 5 shows the immunofluorescence intensity of cyclin B1 in eggs cultured in vitro. Postovulatory egg aging resulted in a gradual decrease of immunofluorescence intensity of cyclin B1 in eggs cultured in vitro (Fig. 5A1–A6). CTCF analysis of the immunofluorescence intensity of cyclin B1 further suggests that postovulatory aging was associated with a time-dependent (one-way ANOVA, F = 1577.025, p < 0.001; Fig. 5B) decrease of cyclin B1 in eggs cultured in vitro.
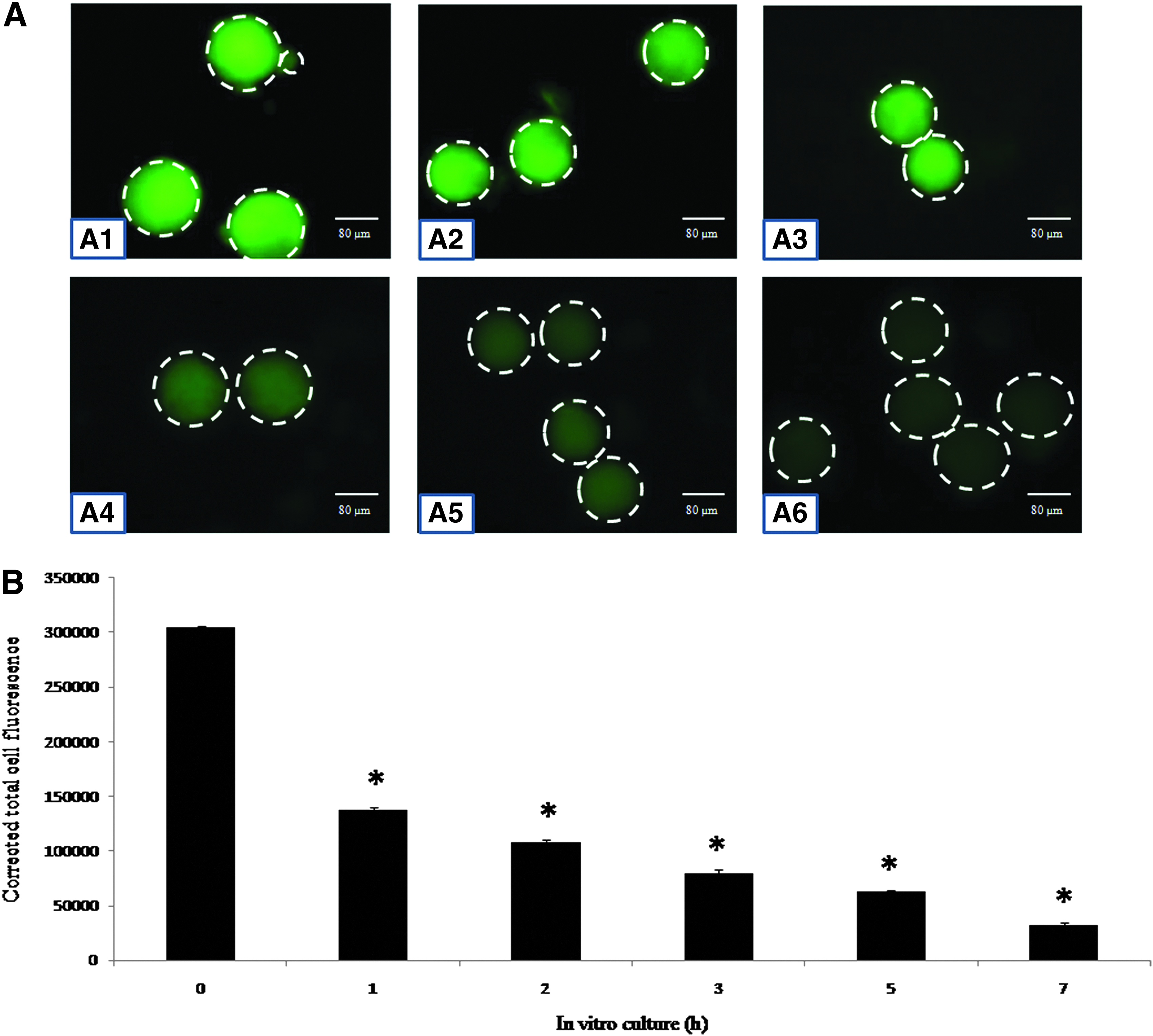
(
Wee1 level changes during egg aging in vitro
Figure 6 shows changes in the Wee1 level in eggs cultured in vitro. The immunofluorescence intensity of Wee 1 increased during the initial period (Fig. 6A1–A4) and then reduced significantly (p < 0.001) during later stages of in vitro culture (Fig. 6A5–A6). CTCF analysis of Wee1 fluorescence intensity (Fig. 6B) revealed that the culture of MII arrested eggs in medium increased the Wee1 level in a time-dependent manner (one-way ANOVA, F = 19.3, p < 0.001; Fig. 6B) during the initial period (up to 3 h) and then fluctuated during later stages of in vitro culture (up to 7 h).
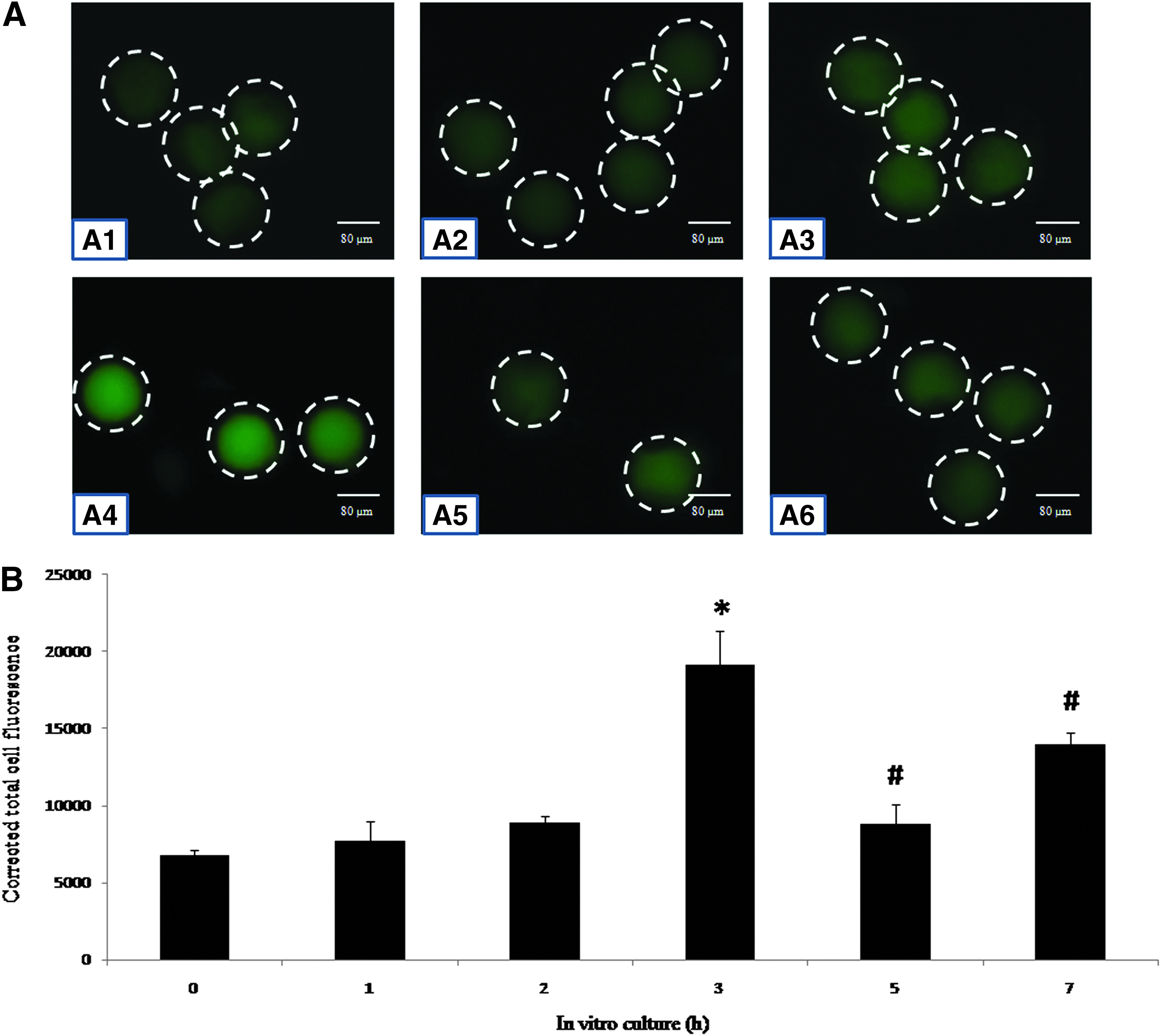
(
Specific phosphorylation of Cdk1 is altered during in vitro egg aging
As shown in Figure 7, postovulatory aging was associated with a gradual increase of the Thr-14/Tyr-15 phosphorylated Cdk1 level (Fig. 7A1–A4) and then declined during later stages of in vitro culture (Fig. 7A5–A6). CTCF analysis further confirmed a time-dependent increase of the Thr-14/Tyr-15 phosphorylated Cdk1 level (one-way ANOVA, F = 47.378, p < 0.001; Fig.A7) during the initial period and then a decrease during the later period of in vitro culture (one-way ANOVA, F = 39.651, p < 0.001; Fig. A7). On the other hand, the Thr-161 phosphorylated Cdk1 level gradually declined during egg aging in vitro (Fig. 7B1–B6). CTCF analysis of fluorescence intensity of Thr-161 phosphorylated Cdk1 decreased in a time-dependent manner (one-way ANOVA, F = 126.710, p < 0.001; Fig. B7). The total Cdk1 level did not change significantly during in vitro studies (Fig. 7C1–C6).
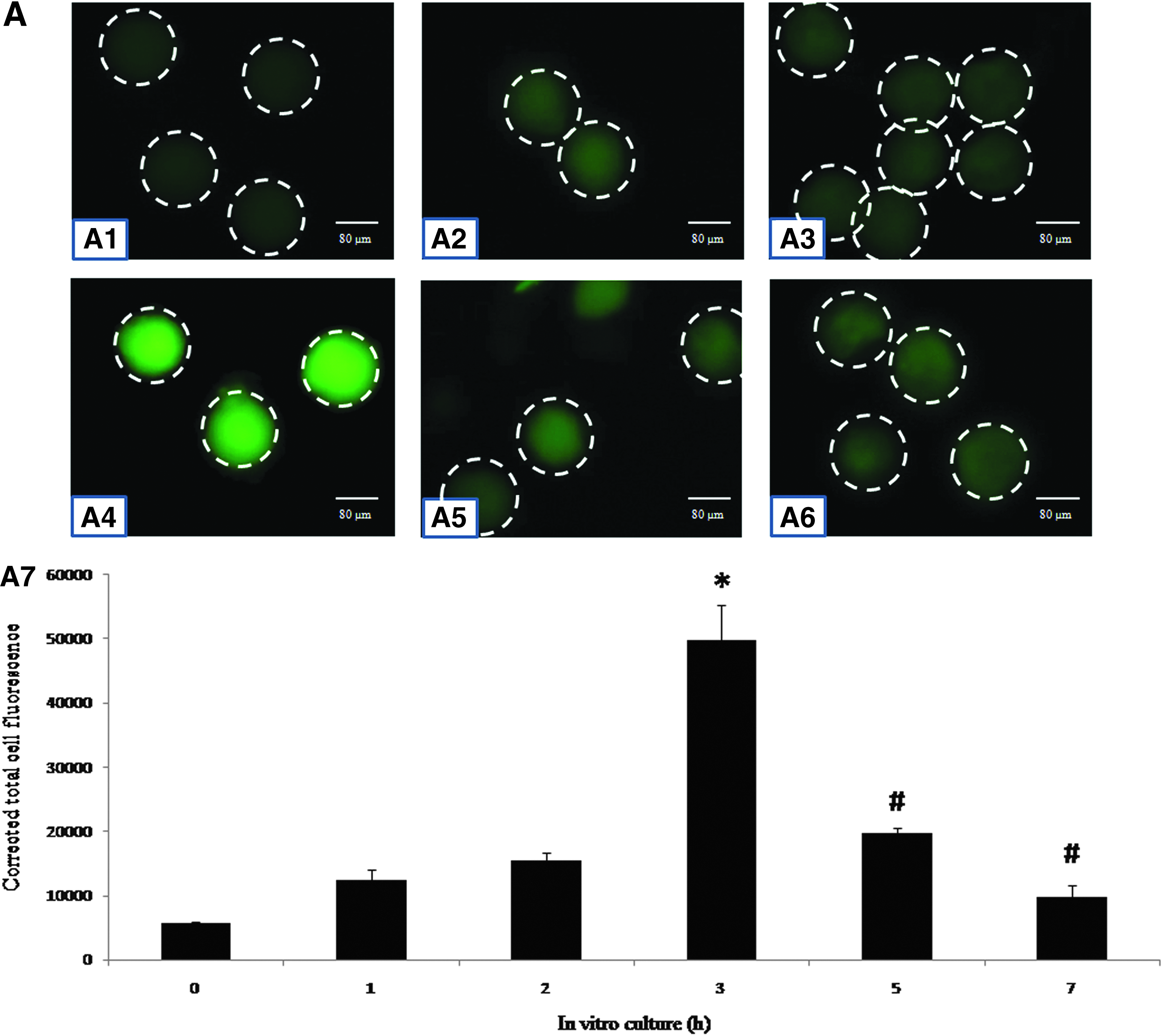
(
Cdk1 activity increased during egg aging in vitro
Figure 8 shows Cdk1 activity in eggs cultured in vitro. Postovulatory aging resulted in a time-dependent (one-way ANOVA, F = 145.134, p < 0.001) increase of Cdk1 activity during the initial period (up to 5 h), whereas enzyme activity declined significantly thereafter if the culture period was extended up to 7 h.
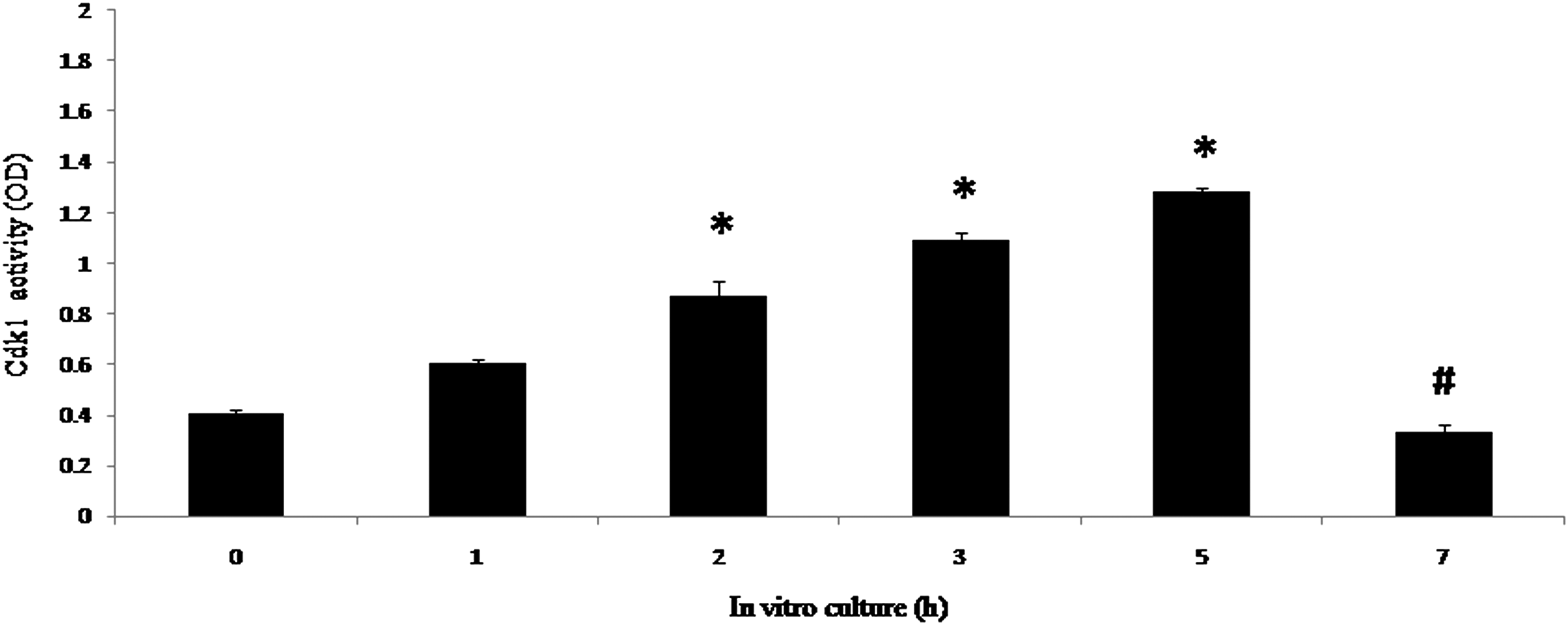
Effect of postovulatory aging on Cdk1 activity during egg aging in vitro. Postovulatory aging increased Cdk1 activity in a time-dependent manner during the initial 5 h of culture and declined thereafter. Data are mean ± SEM of three independent experiments and analyzed by the Student t-test. (*) p < 0.001; (#) p < 0.001.
Discussion
A growing body of evidence suggests that the ovulated eggs do not remain arrested at the MII stage and resume meiosis spontaneously during in vitro aging in several mammalian species (Chebotareva et al., 2011; Miao et al., 2009; Premkumar and Chaube, 2014, 2015). Postovulatory aging mediates abortive SEA and initiation of PBII extrusion, which are characteristic features of SEA (Chebotareva et al., 2011; Prasad et al., 2015b; Premkumar and Chaube, 2013, 2014); this deteriorates egg quality and limits ART outcome (Miao et al., 2009; Prasad et al., 2015a). However, biochemical and molecular changes that drive postovulatory aging-mediated abortive SEA remain poorly understood. Results of the present study suggest that the freshly ovulated eggs are arrested at the MII stage, and culture of these eggs for 7 h resulted in initiation of PBII extrusion in a time-dependent manner. About 40% of eggs showed complete PBII extrusion if cultured for 7 h in vitro. Meiotic stages of eggs were reconfirmed on the basis of chromosomal status. These data further strengthen previous observations that postovulatory egg aging mediates abortive SEA in several mammalian species, including rat (Chebotareva et al., 2011; Miao et al., 2009; Prasad et al., 2015b; Premkumar and Chaube 2013, 2014).
Emi2 is one of the upstream cytostatic factors that maintain stabilized MPF and thereby MII arrest in freshly ovulated eggs (Madgwick and Jones, 2007). A possibility exists that the reduction or degradation of Emi2 during postovulatory egg aging may result in MPF destabilization under in vitro culture conditions. Our results suggest a transient decrease after 1 h of Emi2 level, which remained at its minimum level until 7 h of in vitro culture. Similarly, a high level of Emi2 maintains MII arrest (Kubiak et al., 2008; Madgwick and Jones, 2007), whereas reduction in its level triggers a meiotic exit from MII arrest during postovulatory egg aging in vitro (Kubiak et al., 2008; Rauh et al., 2005).
Emi2 is responsible for the inhibition of APC/C during maintenance of MII arrest in ovulated eggs (Hansen et al., 2007; Sako et al., 2014; Schmidt et al., 2005; Wu and Kornbluth, 2008). On the basis of these studies, we propose that the reduction of Emi2 may result in the activation of APC/C. Our data suggest that the APC/C level increased significantly during the initial period of egg aging (up to 2 h) and then reduced to the level of the control after 7 h of in vitro culture. These results suggest that the initial decrease of Emi2 level after 1 h might have resulted in the increase of APC/C expression after 2 h of in vitro culture. The reduced APC/C expression could be associated with abortive SEA during in vitro egg aging. MAD2 is another CSF that inhibits APC/C activity by binding with its adapter cdc20 (Fang et al., 1998; Kubiak et al., 2008; Yu, 2002). Our results suggest that the MAD2 level increased during the initial period (2 h) and then gradually declined after 7 h of in vitro culture. These results are in agreement with previous findings that the decrease of the MAD2 level is associated with APC/C activation and thereby cyclin B1 degradation (Homer et al., 2005; Kubiak et al., 2008).
Once the APC/C is active, it triggers cyclin B1 degradation through a ubiquitin-mediated pathway (Fry and Yamano, 2006; Kubiak et al., 2008; Passmore, 2004; Peters, 2002). The increased APC/C activity during the initial period of in vitro culture might have induced cyclin B1 degradation. This possibility was further strengthened by our results that cyclin B1 was reduced in a time-dependent manner during egg aging in vitro. The decrease of the cyclin B1 level during postovulatory aging-mediated abortive SEA has been reported in rat eggs (Chesnel et al., 2006; Kubiak et al., 2008; Murray et al., 1989; Premkumar and Chaube, 2013, 2014; Prasad et al., 2015b). Cyclin B1 degradation is not solely responsible for MPF destabilization and thereby meiotic exit from MII arrest (Chesnel et al., 2006); changes in specific phosphorylation of Cdk1 may also trigger MPF destabilization and thereby meiotic resumption from MII arrest (Oh et al., 2011).
Cdk1 is a catalytic unit of the MPF heterodimer, and its association with cyclin B1 is required for MPF stabilization. Association of Cdk1 and cyclin B1 depends upon changes in the phosphorylation status of Cdk1 at Thr-14/Tyr-15 and Thr-161 (Oh et al., 2011, 2013; Prasad et al., 2015b). Wee1 kinase is an upstream modulator of the Thr-14/Tyr-15 phosphorylation status of Cdk1 (Oh et al., 2011). On the basis of these studies, we hypothesize that changes in the Wee1 kinase level could modulate the specific phosphorylation status of Cdk1. This notion is further strengthened by our data that the Wee1 level increased during first 3 h of egg aging in vitro. Thereafter, its level declined gradually and was comparable to the control after 7 h of egg aging.
A similar pattern of changes in the Thr-14/Tyr-15 phosphorylated Cdk1 level was observed in aging eggs in vitro. The Thr-161 phosphorylated Cdk1 level declined in a time-dependent manner, whereas the total Cdk1 level remained unchanged during 7 h of in vitro aging. These data further strengthen previous observations that the high level of Wee1 kinase associates with an increased Thr-14/Tyr-15 phosphorylated Cdk1 level and a decreased Thr-161 phosphorylated Cdk1 level during postovulatory aging-mediated abortive SEA (Oh et al., 2011, 2013; Prasad et al., 2015a,b; Premkumar and Chaube, 2014, 2015).
Recent studies suggest that increased Cdk1 activity is required for meiotic resumption from diplotene arrest in bovine and porcine oocytes cultured in vitro (Davydenko et al., 2013; Jang et al., 2014). On the basis of these studies, we propose that increased Cdk1 activity could be associated with the abortive SEA in aged eggs. Our data suggest that a time-dependent increase of Cdk1 activity associates with postovulatory aging-mediated abortive SEA. These data together with previous studies suggest that both specific phosphorylation and dephosphorylation of Cdk1 and Cdk1 activity are associated with postovulatory aging-mediated abortive SEA in rat eggs cultured in vitro. However, a correlation between these two events warrants further investigation.
In summary, our results suggest that postovulatory egg aging decreases Emi2 as well as MAD2 levels. Decreased levels of CSFs might have resulted in the activation of APC/C, leading to cyclin B1 degradation in aged eggs cultured in vitro. Furthermore, postovulatory egg aging increased Wee1 and Thr-14/Tyr-15 phosphorylated Cdk1 levels but decreased Thr-161 phosphorylated Cdk1 as well as cyclin B1 levels, which resulted in MPF destabilization and increased Cdk1 activity. Thus, destabilized MPF and increased Cdk1 activity are associated with postovulatory aging-mediated abortive SEA in rat eggs cultured in vitro.
Footnotes
Acknowledgment
This study was financially supported by Department of Biotechnology, Ministry of Science and Technology, Government of India.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.