
Abstract
Abstract
Embryonic body-like colony formation is a unique pattern in male germ cell cultures, including spermatogonial stem cells. However, detailed information of the colony formation has not yet been sufficiently reported in male germ cell culture. To elucidate the formation of germ cell-derived colony (GDC), glial cell-derived neurotrophic factor receptor alpha-1 (GFRα-1)-positive pig germ cells were isolated using an immunomagnetic cell isolation method and labeled with red- or green-fluorescent dye. In GDC culture, red-fluorescent-labeled germ cells were evenly distributed in the wells from day 1 to 4, and they clustered together at the time of GDC formation on day 6. Interestingly, feeder cells migrated to the site of colony formation as spermatogonia carriers. Furthermore, when freshly prepared green-labeled GFRα-1-positive germ cells were added, mixed-fluorescent dye (red and green) colonies were observed. On bromodeoxyuridine (BrdU) treatment, 58% ± 3.13% of germ cells were positive to protein gene product 9.5 but negative to BrdU cells. Immunocytochemistry and reverse transcription-polymerase chain reaction results showed that cultured GDC cells were positive to stem cell- and pig germ cell-specific marker genes. In conclusion, in vitro formation of GDCs is mainly dependent on the aggregation of single germ cells as well as on the slow proliferation of germ cells.
Introduction
I
Spermatogonial stem cells (SSCs) are the primary germ cells of spermatogenesis and male fertility and are similar to other tissue-specific stem cells (Phillips et al., 2010). So far, the establishment and culture of SSCs have been attempted in mammals (Sato et al., 2011b; Zheng et al., 2014). A culture of male germline stem cells from hamster and mouse testes has been maintained for 1 year and 5 months, respectively (Kanatsu-Shinohara et al., 2003, 2008). In humans, stage-specific, embryonic antigen-4-positive SSCs were cultured for 4 months without feeder cells (Kokkinaki et al., 2011); whereas in pigs, purified testicular cells that were cultured for 8–14 days formed colonies (Kuijk et al., 2009). Glial cell line-derived neurotrophic factor is important for self-renewal and survival of SSCs, and for their subsequent proliferation in the spermatogonia culture system (Aponte et al., 2006).
In addition, an in vivo system using the transplantation technique has provided evidence of SSCs, which are defined by their functional ability to reform spermatogenesis after colonization in a recipient's testes and to produce offspring; spermatozoa can also be produced by an in vitro culture system using mouse germline stem cells and spermatogonia (Abu Elhija et al., 2012; Oatley and Brinster, 2012; Ryu et al., 2005; Sato et al., 2011a). In pigs, glial cell-derived neurotrophic factor receptor alpha-1 (GFRα-1)-positive germ cells exhibit the characteristics of SSCs; the germ cell-derived colony (GDC) is effectively formed within 10 days at 31°C culture condition; and transplanted GDCs are colonized in the recipient's testes at 8 weeks post-transplantation (Lee et al., 2013a, 2013b). However, detailed elucidation of GDC formation has not yet been studied in male germ cell culture, although there is a report on embryonic stem cells (ESCs). To understand physiological cellular characteristics and movement of prospermatogonia (or spermatogonia) in vivo, it is important to verify migration and proliferation of germ cells in in vitro culture. In this study, we demonstrate germ cell adhesion and proliferation in GDC formation using cell labeling and live video images.
Materials and Methods
Testis preparation
A total of 10 testes from Yorkshire and Landrace crossed 5-day-old hybrid piglets were obtained from “Samwoo Livestock” (Yangpyeong, Korea) after castration. The testes were fixed in 4% paraformaldehyde (PFA; Sigma-Aldrich, St. Louis, MO) for histology and immunocytochemistry (ICC) analyses. The tunica albuginea of the testes was removed for testicular cell culture, whole-mount immunohistochemistry preparations, and immunomagnetic cell isolation. The study protocol and standard operating procedures were reviewed and approved by the Institutional Animal Care and Use Committee of Konkuk University (IACUC Approval No. KU13035).
Whole-mount immunohistochemistry
Enzyme A (5 mg/mL collagenase, 0.01 mg/mL DNAse I, 0.1 mg/mL soybean trypsin inhibitor, and 0.1 mg/mL hyaluronidase) was added to decapsulated testes at a five-fold volume of enzyme to testes (v/w) and incubated for 10 minutes at room temperature (RT). Seminiferous cords were collected and fixed in 4% PFA at 4°C overnight. They were washed three times with phosphate-buffered saline (PBS) and treated with 5% serum replacement (Gibco, Carlsbad, CA) and 0.1% Triton X-100 for blocking. The seminiferous cords were then incubated with anti-protein gene product 9.5 (PGP 9.5) raised in rabbits against human PGP 9.5 antigen (1:500; AbD Serotec, Raleigh, NC). Other cords were incubated with anti-GFRα-1 raised in rabbits against human GFRα-1 antigen (1:500; Santa Cruz Biotechnology, Santa Cruz, CA) at 4°C overnight and washed three times with PBS. Some of the cords were incubated with 1% bovine serum albumin (BSA) as a negative control. The seminiferous cords were incubated with anti-rabbit Alexa 488 (1:500; Invitrogen, Carlsbad, CA) for the GFRα-1 antibody and anti-rabbit Alexa 568 (1:500; Invitrogen) for the PGP 9.5 antibody. This was followed by incubation in 3,3′-diaminobenzidine for staining (Vector Laboratories, Burlingame, CA). The cords were then washed with PBS and observed under a light microscope (Nikon, Tokyo, Japan) at 200× magnification.
Collection of GFRα-1-positive germ cells and labeling
Collection of GFRα-1-positive germ cells was performed as previously described (Lee et al., 2013a). Briefly, the testes were treated with collagenase enzyme, the enzyme-treated testes were meshed using a 40-μm nylon mesh, and red blood cells (RBCs) were eliminated using RBC lysis buffer (Sigma-Aldrich). Two Percoll densities, 20% and 40% (Sigma-Aldrich), were used for germ cell collection. The single-cell suspension (2 mL of 5 × 106 cells/mL) was loaded on top of the gradient and centrifuged at 600 g for 10 minutes at 4°C. Interface layers were collected and analyzed by incubation on gelatin-coated dishes; germ cells were located at the interface between the 20% and 40% Percoll layers. The interface was diluted with a basic medium (Dulbecco's modified Eagle's medium; Gibco) containing 10% FBS (v/w), 2 mM
To investigate the formation of GDCs, GFRα-1-positive germ cells were collected using magnet-activated cell sorting (MACS) large-scale columns and MACS separators (Militenyi Biotech, Bergisch Gladbach, Germany), and they were labeled with PKH26 (red dye) or PKH67 (green dye) (Sigma-Aldrich), which was followed by addition to testicular cell culture. Immunomagnetic cell isolation was performed according to the manufacturer's protocol.
Cell culture for GDC formation
Decapsulated testes were weighed, and a five-fold volume (v/w) of enzyme A (0.5 mg/mL collagenase, 0.01 mg/mL DNAse I, 0.1 mg/mL soybean trypsin inhibitor, and 0.1 mg/mL hyaluronidase) was added for 10 minutes at RT. The testes were washed with PBS, enzyme B (5 mg/mL collagenase, 0.01 mg/mL DNAse I, and 0.1 mg/mL soybean trypsin inhibitor) was added at a five-fold volume (v/w) of the original weight of the testes for 10 minutes, and the testes were washed with PBS at RT. The testes were then meshed using a 40-μm nylon mesh, and RBCs were eliminated using RBC lysis buffer (Sigma-Aldrich). The isolated cells were used for ICC analysis, seeded onto 0.2% (w/v) gelatin-coated 12-well plates, and incubated at 31°C, 34°C, or 37°C in 5% CO2 (2 × 105 cells/well).
Stempro-34 medium (Gibco) was used for all processes, including derivation and culture of porcine SSCs. The medium was supplemented with insulin-transferrin-selenium (25, 100 μg/mL, or 30 nM), 6 mg/mL glucose, 2 mM
Live video imaging
Testicular cells were cultured in gelatin-coated 35-mm dishes (ibidi, Munich, Germany) with Stempro-34 medium at 31°C in 5% CO2. Live video images were captured from day 3 to 6 (first recording) and from day 7 to 10 (second recording); the medium was changed once every 3 days. Live video imaging during the testicular cell culture was performed using the Tokai Hit incubation system for microscopes (Tokai Hit Co., Ltd., Shizuoka-ken, Japan). Movies were recorded using the Nikon Eclipse TS 100 microscope (Nikon, Chiyoda-ku, Japan) and analyzed using the iSolution software program (iMT Technology, Inc., Seoul, Korea). Live imaging movie files were generated from a time series of images that were captured every minute for 72 hours for the first and the second recording.
Inhibition of germ cell aggregation in colony formation
Red-labeled GFRα-1-positive cells were cultured with 100 μM RGD peptides (an inhibitor of integrin-ligand interactions) (Santa Cruz Biotechnology) for 24 hours and added to testicular cell culture for colony formation. Images of the red-labeled germ cells were captured with an excitation filter of 450–560 nm and a fluorescent microscope (Nikon) at 100× magnification.
Alkaline phosphatase staining
Alkaline phosphatase (AP) staining was performed using a CBA-300 AP Staining Kit (Cell Biolabs, San Diego, CA), according to the manufacturer's instructions. Briefly, the colonies were fixed in fixation solution, washed three times with PBS, and incubated with AP staining solution for 20–30 minutes. Subsequently, the AP solution was removed, cells were washed with PBS, and the AP-stained cells were observed under a light microscope (Nikon) at 200× magnification.
Immunocytochemistry
After GDC formation in testicular cell culture, the colonies were collected and treated with a 0.25% trypsin-EDTA solution at RT for 5 minutes (Sigma-Aldrich). Single cells were fixed with 4% PFA (w/v) in PBS and washed with PBS. They were then attached to amino-silane-coated slides (Matsunami, Osaka, Japan) for ICC. The slides were incubated with anti-PGP 9.5 raised in rabbits against human PGP 9.5 antigen (1:100; AbD Serotec) and anti-GFRα-1 raised in mice against human GFRα-1 antigen (1:50; Santa Cruz Biotechnology) at 4°C overnight for double staining, and they were subsequently washed with PBS. The slides were incubated with anti-mouse Alexa 488 (1:500; Invitrogen) for the GFRα-1 antibody and anti-rabbit Alexa 568 (1:500; Invitrogen) for the PGP 9.5 antibody. Cell nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI) staining solution (Vector Laboratories) and observed with an excitation filter of 450–560 nm and under a fluorescent microscope (Nikon) at 400× magnification.
Bromodeoxyuridine treatment and detection
Bromodeoxyuridine (BrdU) Labeling Reagent (Invitrogen) was applied to the testicular cell culture. Ten micromolars of BrdU solution in DMSO was added to the culture media from day 1 to 5. When the colony was formed on day 12, the cells from the dishes were collected and separated using a 0.25% trypsin-EDTA solution. Single cells from the dishes were attached to amino-silane-coated slides (Matsunami). ICC analysis was performed with a PGP 9.5 and a BrdU antibody (AbD Serotec and Santa Cruz Biotechnology) The ICC procedure was identical to the PGP 9.5 and GFRα-1 staining procedures. The cells attached to the slide glass were incubated with anti-PGP 9.5 and anti-BrdU (1:100; Santa Cruz Biotechnology) at 4°C overnight for double staining. PGP 9.5- and BrdU-positive cells were counted by observation with an excitation filter of 450–560 nm and under a fluorescent microscope (Nikon) at 100× and 400× magnification.
Reverse transcription-polymerase chain reaction (RT-PCR)
Whether or not labeled cells have germ and stem cell characteristics as reported in a previous study, total RNA from the collected unlabeled, red- and red and green-labeled GDCs was isolated using the RNeasy Mini Kit (Qiagen, Venlo, The Netherlands). cDNA templates were prepared from 1000 ng of total RNA using the Maxime RT Premix kit (Intronbio, Seongnam, Korea). The cDNA synthesis conditions were one cycle of 60 minutes at 94°C. cDNA synthesis was stopped by heating (5 minutes at 95°C). Gene-specific primers for beta-2-microtubulin, PGP 9.5, GFRα-1, promyelocytic leukemia zink finger (PLZF), octamer-binding protein 4 (Oct4), Nanog, luteinizing hormone receptor, and GATA-binding protein 4 (GATA4) were obtained from a previous study (Lee et al., 2013a). The cycling conditions were 33 cycles, each consisting of 1 minutes at 94°C, 1 minute at 55°C–60°C, and 2 minutes at 72°C, for all genes. The PCR products of all genes were detected by electrophoresis on a 1.5% gel with Tris-acetate-EDTA buffer.
Results
Detection of germ cells in neonatal pig testes
In a previous study, we reported that PGP 9.5 and GFRα-1 are exclusively detected in the germ cells of the neonatal pig testes sections (Lee et al., 2013a). Whole-mount immunohistochemistry was performed with neonatal pig testes tubules for spatial observation of germ cell localization. Results showed that germ cells are localized in the seminiferous cord. PGP 9.5-positive germ cells were detected in almost all the germ cells in the seminiferous cords; the germ cells were either single or paired (Fig. 1A-a–d). Localization of PGP 9.5-positive germ cells in whole-mount immunohistochemistry spatially matches that of PGP 9.5-positive spermatogonia in immunohistochemistry (Lee et al., 2013a). GFRα-1-positive cells were detected in germ cells but not in all the germ cells in a cord (Fig. 1A-e–h). These data are consistent with our previous results, which showed that GFRα-1-positive and -negative germ cells exist in the neonatal pig testes (Lee et al., 2013a). ICC was performed on single testicular cells after enzyme treatment. PGP 9.5- and GFRα-1-positive germ cells were observed in testicular cells; however, the proportion of stained cells in testicular cells was small. The observed proportion of GFRα-1-positive germ cells was smaller than that of PGP 9.5-positive germ cells (Fig. 1B-a–h). Morphologically, the rounded and bigger cells were testicular germ cells (Fig. 1B-d, h).
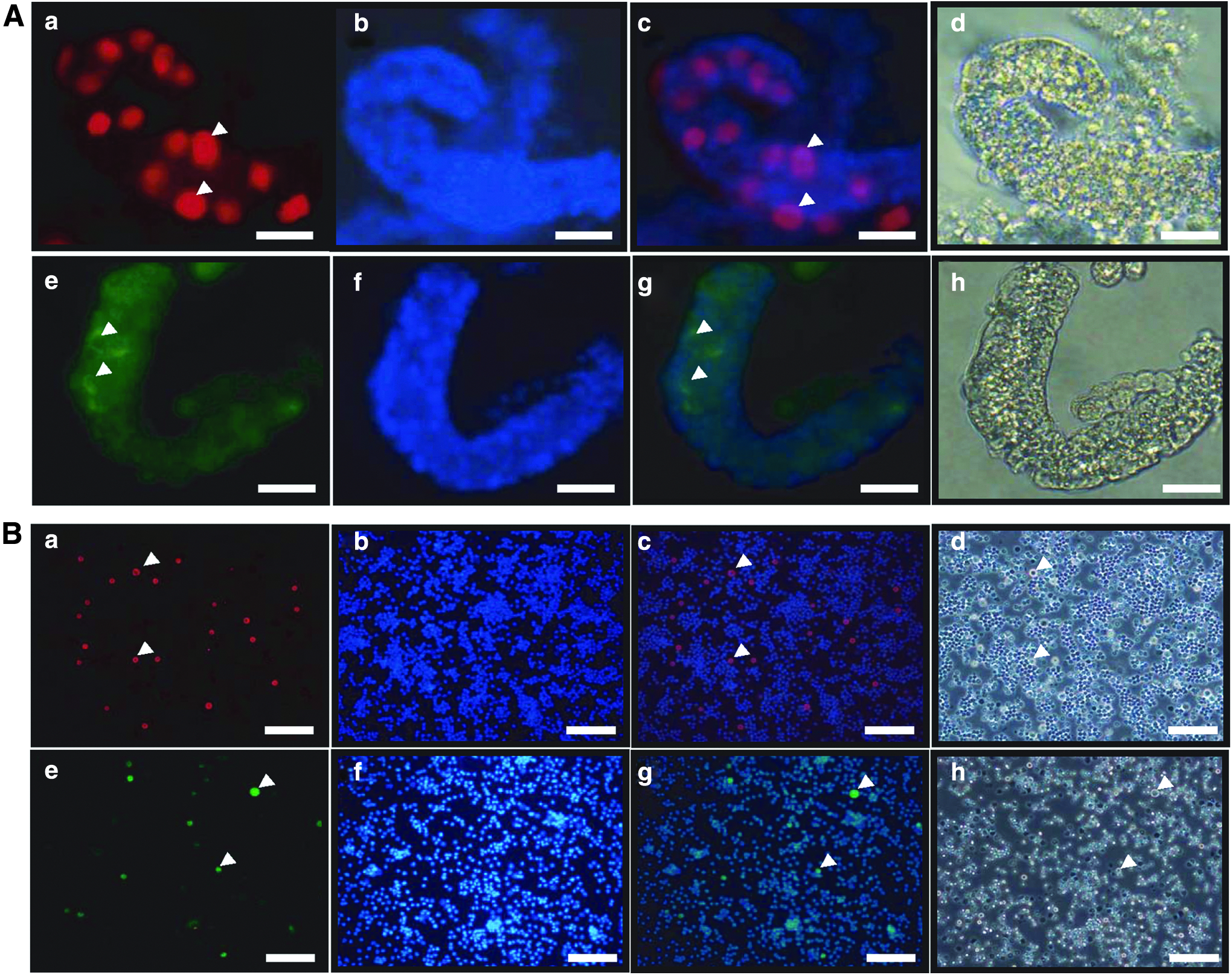
Detection of germ cells in the seminiferous cord and single cells isolated from neonatal pig testes.
Formation of germ cell-derived colonies
In our previous results, we showed that the optimal temperature for GDC in in vitro culture is 31°C (Lee et al., 2013b). In the current results, the formation of small colonies was observed at day 6 and an increase in the colony size was observed with time (Fig. 2A–D). Whether the germ cells were clustered in colony formation or not, red-labeled germ cells, which were GFRα-1 positive and collected by immunomagnetic cell isolation, were used as the tracker. GFRα-1-positive germ cells labeled with red-fluorescent dye were dispersed until day 3 (Fig. 2E). GDC formation with red-fluorescent GFRα-1 germ cells was observed on day 6 (Fig. 2F). As time progressed, the size of the colonies with red-fluorescent GFRα-1 germ cells increased on days 9 and 12 (Fig. 2G, H). To investigate GDC aggregation in more detail, green-fluorescent GFRα-1 germ cells were added to the GDC culture with the red-colored GFRα-1 germ cells at the time of colony formation (at day 6), and live video images were captured during colony formation.

GDC formation in culture with testicular cells and red-labeled GFRα-1-positive cells. Testicular cell culture and red-labeled GFRα-1 germ cells were cultured for 12 days.
Green-fluorescent GFRα-1 germ cells were attached to GDC with red-fluorescent GFRα-1 germ cells at 9 days (Fig. 3A–C). Green-fluorescent GFRα-1 germ cells, which were attached to GDC with red-fluorescent GFRα-1 germ cells, increased at 12 days (Fig. 3E–G). Merged images of colonies containing red- and green-fluorescent GFRα-1 germ cells showed that red- and green-fluorescent GFRα-1 germ cells aggregated independently in colony formation (Fig. 3C, G). Morphologically, the colonies were similar to the colonies in Figure 2 (Fig. 3D, H). As observed in the live video image from day 3 to 6, germ cells were carried by feeder cells that were proliferating at the bottom of the dish (Supplementary Video S1; Supplementary Data are available online at www.liebertpub.com/cell). In the live video image from day 7 to 10, the germ cells aggregated together with the aid of feeder cell proliferation and a colony formed with an increasing size (Supplementary Video S2). Cell aggregation was blocked with an integrin blocker (RGD peptides). GFRα-1-positive germ cells, which were treated with RGD peptides and labeled with red-fluorescent dye, were cultured with testicular cell culture.

Aggregation of germ cells and blockage of germ cell aggregation during GDC formation.
As a result, red-labeled GFRα-1-positive germ cells, which were treated with RGD peptides, did not participate in colony formation on days 3, 6, and 9 (Fig. 3I–K). Normal colonies and feeder cell proliferation were observed, as seen in Figure 2 (Fig. 3L–N). To detect the proliferating germ cells, double-staining ICC was performed in the GDC cells. BrdU was applied to forming GDCs from days 1 to 5. The colonies were not observed within day 6 but appeared on day 9, whereas normal colonies shown in Figure 2D were observed on day 12 (Fig. 4A–C). In the ICC results, PGP 9.5- and BrdU-positive germ cells were detected in the cells collected from the dishes at day 12; PGP 9.5-positive and BrdU-negative cells were also detected (Fig. 4D, E, G). Proliferating feeder cells were observed in BrdU-negative cells (Fig. 4E–H). The percentage of PGP 9.5-positive and BrdU-negative germ cells was 58% ± 3.13% (Fig. 4I).

Detection of proliferating germ cells in GDC formation. In BrdU treatment, images were captured on
Characterization of spermatogonia-derived colony cells
Molecular analyses were performed to investigate the germ and stem cell characteristics of the unlabeled, red- and red and green-labeled GDCs. Double-staining ICC results showed that PGP 9.5 and GFRα-1 proteins were detected in the cells from GDCs on day 12; these GDCs contained clusters of red-labeled germ cells (Fig. 5A-a, b). Their morphology was similar to that observed in our previous results (Fig. 5A-c, d) (Lee et al., 2013a). Furthermore, staining with AP, which is a universal pluripotent marker for all types of pluripotent stem cells, including ESCs, embryonic germ cells, and induced pluripotent stem cells (iPSCs), was performed. GDCs were positive for AP (Fig. 5B-a). Fortunately, red-fluorescent cells still remained in the colonies after AP staining (Fig. 5B-b). In RT-PCR results, PGP 9.5, GFRα-1, PLZF, Nanog, and Oct4, which are known to be early spermatogonia and stem cell markers, were expressed in unlabeled, red- and red and green-labeled GDCs; however, GATA4, which is a known Sertoli cell marker, was not expressed in GDCs (Fig. 5C). Unlabeled, red- and red and green-labeled GDCs have almost the same gene expression in RT-PCR results, which means that red or green labeling does not affect the colony's characteristics.
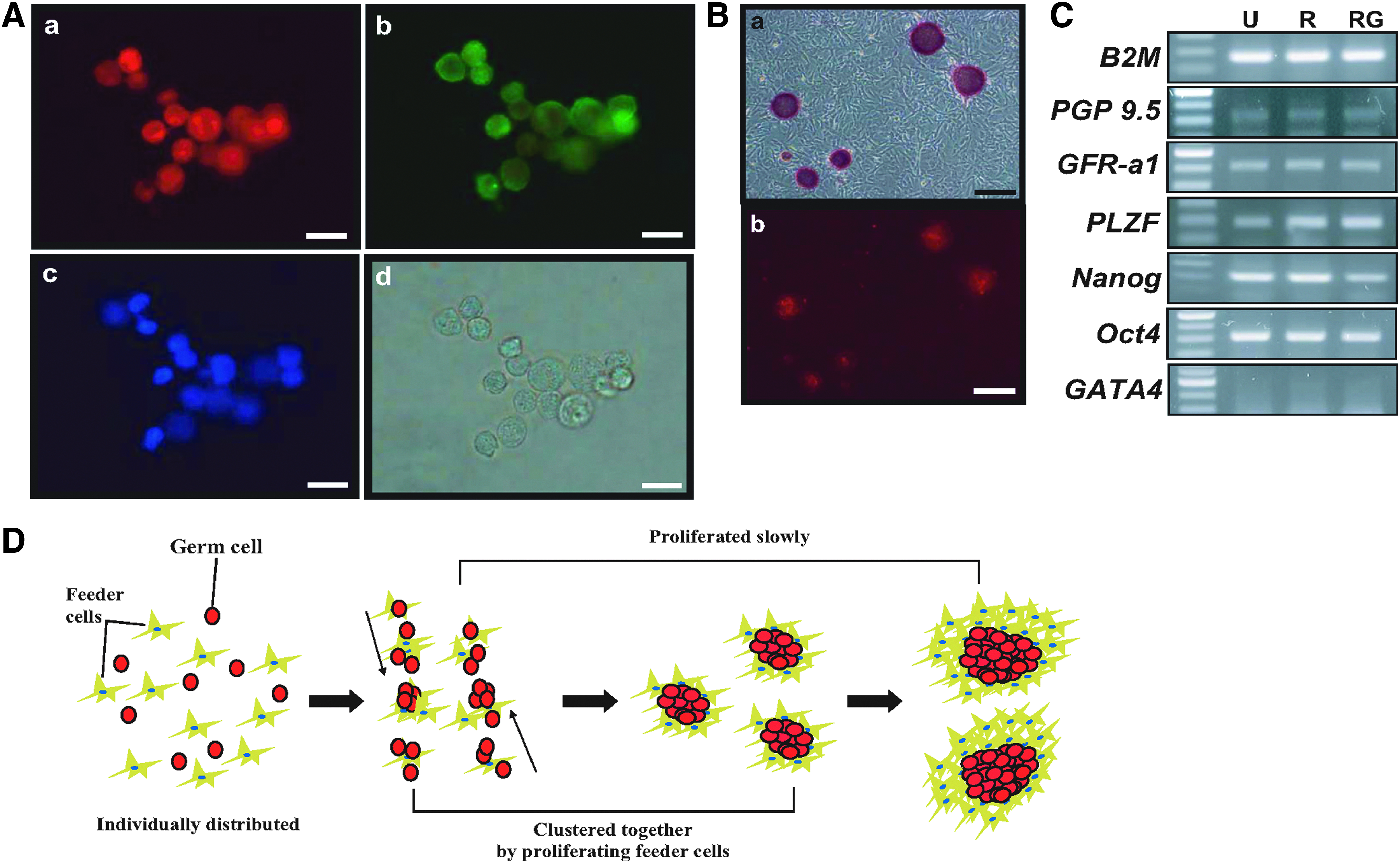
Characterization of germ cell-derived colonies collected on day 12 and the principle of GDC formation in vitro.
Discussion
Studies on colonies in ESCs and iPSCs have been reported; morphologically rounded stem cell colonies were formed in the culture of ESCs or iPSCs using feeder cells (Amit and Itskovitz-Eldor, 2012). There is a notable study by Kim et al. (2013) on colony formation of hESCs in feeder-free culture conditions. They reported that colony formation of enzymatic-dissociated hESCs was enhanced by pertussis toxin (PTX) treatment. PTX treatment resulted in the development of denser colonies formed by migration and aggregation (Kim et al., 2013). In a single cell-labeling experiment and live image capture, we found that red-labeled porcine germ cells clustered together to form colonies when red-labeled GFRα-1-positive germ cells were added to the total testicular cell culture. A similar finding has been reported in the colony formation experiment; a neural stem cells colony, which was differentiated from hESCs, was formed by aggregation of red and green hESCs that were cultured in the same well (Chaddah et al., 2012). These results suggest that aggregation of single germ cells, which have germ and stem cell characteristics, occurs in GDC formation in a manner similar to that observed in hESC colony formation.
In neonatal mice testes, seminiferous cords grow along with the increase of body size and contain only undifferentiated germ cells. The first meiotic division into spermatocytes occurs within 14–20 days (Montoto et al., 2012). Only germ cells are observed in the canine testis after 1–3 months; the size of the tubules and the number of germ cells increase over time (Lee et al., 2014). In the neonatal pig testes in the current study, only germ cells were observed in month 2, and the first spermatogenic cells (spermatocytes) appeared between 2 and 3 months (Supplementary Fig 1A–C).
These results indicate that before the differentiation of the first spermatogonia into spermatocytes, the proliferation of germ cells requires a longer duration in dogs and pigs than in mice. In the seminiferous tubule, the distribution of Sertoli cells before the first spermatogenesis is different. In humans, mice, dogs, and pigs, Sertoli cells are more abundant than germ cells in the perinatal or neonatal period. After the appearance of the first spermatocytes around the infantile or pubertal period, the number of Sertoli cells is smaller than in the neonatal or perinatal period; the seminiferous cord also grows, and the number of germ cells increases in the perinatal or neonatal period (Lee et al., 2014; Montoto et al., 2012; Sharpe et al., 2003) (Supplementary Fig. S1A–C).
In the author's previous study, similar results were reported in fish, stating that doublesex- and mab-3-related transcription factor 1 (DMRT1)-expressing Sertoli cells are abundant during early gonadal development in pufferfish (before spermatogenesis) (Lee et al., 2009; Yamaguchi et al., 2006). In live image capture, proliferating feeder cells that were attached on the dish carried germ cells; GDCs were formed by their movement; and GDC cells were proliferated as time went on. These results suggest that a number of testicular somatic cells (especially Sertoli cells) are involved in undifferentiated germ cell proliferation before the first spermatogenesis in vivo. In addition, it is clear that the period for the germ cell proliferation is different in each species before the first spermatogenesis. In SSC proliferation, murine SSCs expanded ∼5 × 1012-fold from the initiation of culture to 134 days, and it kept growing for more than 160 days (2 × 1014-fold expansion in total cell number) (Kanatsu-Shinohara et al., 2003). Hamster SSC proliferation was around 100-fold at day 100 in specific culture medium conditions (Kanatsu-Shinohara et al., 2008). Unlike the proliferation of mouse SSCs, the proliferation of porcine GDCs in testicular cell culture at 31°C was slow (Lee et al., 2013b). On the other hand, in bovine spermatogonia culture, the number of spermatogonia increased continuously and after 2 weeks, ∼2-fold more spermatogonia were detected than those at the start of the culture; the proliferation of spermatogonia was affected by the culture medium, temperature, and density, with a different proliferation pattern in different media (Creemers et al., 2002; Izadyar et al., 2003).
However, rapid proliferation of spermatogonia has not been observed in domestic animals. This is a similar result in comparison of undifferentiated germ cell proliferation between rodents and domestic animals as mentioned earlier. In our results, the percentage of BrdU- and PGP 9.5-positive germ cells that were collected from GDCs was less than 58%, and feeder cells carried germ cells toward GDC formation. Taken together, this suggests that the in vitro germ cell proliferation rate could be consistent with the in vivo germ cell proliferation rate, and that the feeder cells attached on the dish support the proliferation and aggregation of germ cell in vitro culture.
In conclusions, porcine GDC formation, in which feeder cells carry germ cells, consists of germ cells clustering and their slow proliferation in testicular culture (Fig. 5D). These results may help to understand the relationship between testicular germ and somatic cells. However, the reason behind the clustering of stem cells in an in vitro culture remains unclear. Further studies on the stem cell or SSC adhesion and migration in vitro are necessary.
Footnotes
Acknowledgments
The authors are grateful to Dr. Sung-Hwan Moon and Soon-Jung Park, PhD candidates at Konkuk University, for their help in preparing beautiful live image capture. This study was supported by a grant from the Cooperative Research Program for Agriculture Science & Technology Development (project no. PJ011865), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.