
Abstract
Abstract
Activation-induced cytidine deaminase (AID) is the only enzyme that has been suggested as a putative DNA demethylase in mammals. However, very little is known about AID function as DNA demethylase of bovine differentiated cells toward pluripotent state. To investigate the effect of AID on DNA demethylation, bovine AID complementary DNAs were transfected into bovine differentiated cells, which were mostly methylated in the promoter regions of pluripotency genes. As a result, AID-transfected bovine cells started to transform into colonies at day 19 of transfection. The colonies derived from the transfected cells showed positive alkaline phosphatase (AP) staining and expression of pluripotency genes (OCT-3/4, NANOG, SOX2) and pluripotency-related antigens (SSEA-4, TRA1-60, TRA1-81), which have been widely used to characterize human embryonic stem cells. In particular, the levels of OCT-3/4 and NANOG expression were significantly increased in the AID-transfected cells when compared with the control and empty vector-transfected cells (p < 0.05). Finally, DNA demethylation in the promoter regions of pluripotency genes (OCT-3/4, NANOG) was significantly increased compared with the control (p < 0.05). These results demonstrate that the induction of the AID gene into bovine differentiated cells improves DNA demethylation and expression of pluripotency genes.
Introduction
T
In case of mammalian development, DNA methylation is epigenetically stable and is reversed twice in early embryo cells and in primordial germ cells (PGCs) (Li, 2002; Reik et al., 2001; Surani et al., 2001). The paternal pronucleus commonly undergoes rapid global demethylation in the zygote, a process whose biological significance is very little known (Iqbal et al., 2011; Okada et al., 2010). A second active DNA demethylation occurs in the PGCs to eliminate DNA methylation imprints established in the parental generation (Hajkova et al., 2010; Popp et al., 2010). Epigenetic reprogramming may be necessary for resetting the epigenetic state of the gamete's genome so that totipotency is restored to the embryonic genome.
This process is of interest for identifying how reprogramming takes place in a cell. This could help us understand how cell differentiation and specialized gene expression are preserved. Generally reprogramming of somatic nuclei is caused by epigenetic machinery comprising molecules that regulate epigenetic modifications (DNA methylation and histone modifications), which are involved in heritable gene expression patterns. DNA methylation plays an essential role in the regulation of gene expression and affects the interactions between these modifications and transcription factors or other DNA-binding proteins (Wagner et al., 2014). Reactivation of pluripotency genes occurring during nuclear reprogramming is related to active DNA demethylation (Bhutani et al., 2010).
A number of mechanisms have been proposed for the loss of methylation (Cedar and Verdine, 1999), involving direct demethylation (Bhattacharya et al., 1999), oxidative demethylation (Trewick et al., 2002), and 5-methylcytosine (5mC) base excision repair (BER) (Zhu, 2009). Of those, the mechanism for removing 5mC is closely related to the DNA repair system. This concept presents that cellular DNA repair assures genome stability from exogenous and endogenous DNA lesions and helps epigenetic gene regulation. Activation-induced cytidine deaminase (AID) and Apolipoprotein B RNA-editing catalytic component-1 (Apobec-1) have been shown to deaminate 5mC in single-stranded DNA (Morgan et al., 2004).
AID as a DNA demethylase in mammalian reprogramming has the activity of 5mC deaminase, resulting in generating thymine base opposite a guanine and yielding a G:T mismatch (Bransteitter et al., 2003; Pham et al., 2003). If this mismatch is repaired, a methylated cytosine can be replaced into an unmethylated one (Popp et al., 2010). In particular, AID was shown to be required for demethylation of the promoter regions, which was correlated with the induction of two well-known pluripotency genes (OCT-3/4 and NANOG). In addition, loss-of-function experiments with short interfering RNA (siRNA) knockdown to validate the role of AID in regulating OCT-3/4 and NANOG partially suppress OCT-3/4 demethylation and expression (Bhutani et al., 2013).
These results have suggested that AID plays a critical role to induce demethylation of the key pluripotency genes, which are an essential part of the reprogramming (Deng, 2010). OCT-3/4 acts as an important regulator of pluripotency cells and is thought to play a key factor in controlling murine, bovine, and porcine embryonic development (Kirchhof et al., 2000). Additionally, NANOG is a transcription factor and plays a key role in maintaining pluripotency, critically self-renewal of embryonic stem (ES) cells.
Several strategies have been used to reprogram differentiated cells, yet they are generally characterized by a low efficiency of reprogramming, reflecting remarkable stability of the differentiated state. Thus, we conducted these experiments to examine the effect of AID on DNA demethylation of bovine differentiated cells, which are highly methylated promoter regions of pluripotency genes. To construct AID expression vector, AID genes extracted from bovine immune tissues (lymph node and spleen) were inserted into the pCMV6-AC-IRES-GFP-Puro expression vector. After transfection into bovine somatic cells using a transfection reagent, the AID-transfected cells were cultured for a certain period of time, and then the activity of alkaline phosphatase (AP) and expression of pluripotency genes (OCT-3/4, NANOG, SOX2) and pluripotency-related antigens (SSEA-1, SSEA-4, Tra-1-61, Tra-1-80) were analyzed. Finally, the change in DNA demethylation in the promoter regions of pluripotency genes (OCT4, NANOG) was examined in the AID-transfected bovine cells.
Materials and Methods
All chemicals and reagents were purchased from Sigma-Aldrich Co., unless otherwise mentioned.
Preparation of bovine ear tissues and cell culture
The bovine ear tissues were collected from a Holstein cow. They were washed several times in Dulbecco's phosphate-buffered saline (D-PBS; ions free). Each tissue was chopped up into small pieces. The ear tissues were placed on 60-mm culture dishes and incubated at 39°C for 1 hour without medium. After that they were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% FBS, 1.25% MEM nonessential amino acid, 125 μM β-mercaptoethanol, 2% penicillin/streptomycin, and 0.5 μg/mL amphotericin B at 39°C in 5% CO2. When cells reached appropriate confluency, they were cultured in basic cell culture medium (DMEM containing 10% FBS, 1.25% MEM nonessential amino acid, 125 μM β-mercaptoethanol, 1% penicillin/streptomycin).
PCR amplification and TA cloning
The AID gene was obtained from bovine tissues (lymph nodes, spleen, and small intestine) according to NCBI database. Total RNAs were extracted from these tissues using the RNeasy Plus Mini Kit (Qiagen). The concentrations of RNA were measured by using a spectrophotometer. Complementary DNAs (cDNAs) were synthesized using the RevoScript® RT PreMix (Intron) and amplified by polymerase chain reaction (PCR) using Advantage 2 Polymerase Mix (Clontech). cDNA products were amplified by a set of outer and nested primers to ensure specificity. The first stage of PCR conditions was denaturation at 95°C for 10 minutes, followed by a further 40 cycles of denaturation at 95°C for 40 seconds, 55°C for 40 seconds, 72°C for 1 minute, and final annealing at 72°C for 10 minutes. The second stage of PCR conditions was denaturation at 95°C for 10 minutes, 40 cycles at 95°C for 40 seconds, 56°C for 40 seconds, 72°C for 1 minute, and 72°C for 10 minutes using AID inner primers, including start and stop codons.
To create the site of restriction enzymes (REs) into the AID gene, PCR was finally performed with RE site primer. AID and RE primers were used for sticky ends (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/cell). Gel electrophoresis was performed using 1% agarose gel. Sequence of primers is listed in Table 1. PCR products were purified using the QIAquick Gel Extraction kit (Qiagen), followed by ligation into the TOPO® vector (Invitrogen) and transformation into One shot TOP10 Competent Cells (Invitrogen). The AID gene was expressed in all of bovine lymph node, spleen, and small intestine tissues. Of those, only AID genes extracted from the lymph node were inserted into TA vectors to analyze full sequence of the AID gene. TA vectors with inserted AID gene were extracted and purified using the Gene All® Exprep™ Plasmid SV mini kit (GeneAll). The results of sequencing were analyzed by using BLAST (bl2seq, NCBI). The AID gene derived from the lymph nodes was 100% identical to that of the BLAST (Supplementary Fig. S2).
AID + RE sites + AID 2 inner primer.
AID, activation-induced cytidine deaminase; AT, annealing temperature; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; RE, restriction enzyme; RT-PCR, real-time polymerase chain reaction.
Construction of vector for inducible gene expression
The vector of pCMV6-AC-IRES-GFP-Puro (OriGene) was chosen for inducing expression of AID protein in bovine somatic cells. Both of pCMV6-AC-IRES-GFP-Puro vector and TA vector were digested with HindIII and XhoI (New England Biolabs) at the same time at 37°C for 2 hours. After incubation of the REs, the digested fragments were purified by running a preparative agarose gel. By doing this, we isolated AID fragments from the other sized TA vector and pCMV6-AC-IRES-GFP-Puro vector with sticky ends. The concentration of fragments was measured and equimolar concentration was regulated. After that AID fragments from TA vector were ligated to the pCMV6-AC-IRES-GFP-Puro vector. The complex was incubated using T4 DNA ligase (New England BioLabs) at 16°C overnight, and ligated vectors were transformed into Top 10 competent cells. The white colonies picked from a freshly streaked selective plate were incubated into LB medium containing the appropriate selective agent at 37°C overnight. TA vectors of the inserted AID gene were extracted and purified using the GeneAll® Exprep™ Plasmid SV mini kit (GeneAll).
Efficiency of transfection, selection of cloned cells, and cell culture
When bovine somatic cells reached to half of doubling in a six-well dish (1 × 105/10 cm2), 12 ratio conditions of cloned vector and transfection reagent (Lipofectamine LTX; Invitrogen) were diluted in DMEM (w/o serum and antibiotics). The mixture of transfection reagent was added to each cell to be transfected, and the cells were incubated in the cell culture medium without antibiotics at 37°C for 24 hours. The transfected cells were harvested with trypsin-EDTA (0.05% Trypsin) and formed a pellet using centrifuge (200 g, 5 minutes). The cell pellet was washed by D-PBS (w/o ions) and expression of green fluorescent protein in the cell pellet was analyzed using a flow cytometer (FACS calibur; Becton Dickinson).
On the other hand, the transfected cells were cultured in puromycin selection medium (DMEM containing 10% FBS, 1.25% MEM nonessential amino acid, 125 μM β-mercaptoethanol, 1% penicillin/streptomycin, 1.2 μg/mL puromycin) for 5–6 days. After the treatment, the transfected cells were cultured in stem cell culture medium (DMEM/F-12, 20% KSR [Knockout Serum Replacer; Gibco], 1.25% MEM nonessential amino acid, 0.2 mM β-mercaptoethanol, 2 mM
AP staining
To determine the activity of AP, AP staining was carried out in the bovine transfected cells. The transfected cells were fixed with fixative (4% paraformaldehyde in PBS) for 1–2 minutes and rinsed thrice with 1 × Rinse Buffer (20 mM Tris-HCl, pH 7.4, 0.15 M Nacl, 0.05% Tween-20, Ultra pure water) for 5 minutes. Stain solution (Naphthol/Fast Red Violet/Water solution = 1:2:1) was added to each well for 15 minutes and the number of positive AP staining colonies were counted. Image was captured using a phase-contrast microscope (LEICA).
Expression of pluripotency genes
Total RNAs were extracted from the bovine transfected cells. After genomic DNA elimination, the samples of RNA were ready for reverse transcription to synthesize first-strand DNA according to the manufacturer's instructions using Maxime™ PCR PreMix (i-Taq; Intron). The PCR products were examined by electrophoresis using 1% agarose gel. The primers used are described in Table 2.
Quantitative real-time PCR
Expression of pluripotent marker genes in the control, empty vector, and AID bovine transfected cells was further analyzed by quantitative real-time PCR (qRT-PCR). The level of gene expression was normalized relative to that of the reference gene (glyceraldehyde-3-phosphate dehydrogenase [GAPDH]). Real-time PCR was performed in a StepOne™ Real-Time PCR System using Power SYBR® Green PCR Master Mix (Life Technologies) for detection of double-stranded DNA product. The conditions of q-PCR were followed: initial step for 10 minutes at 95°C, followed by 40 cycles of 15 seconds at 95°C, annealing at temperatures listed in Table 3 for 1 minute, and final dissociation step for 15 seconds at 95°C, 1 minute at 60°C, and 15 seconds at 95°C. The sequences of mRNA of pluripotency genes of interest were obtained from GenBank. Amplification was performed according to the manufacturer's instructions.
Immunocytochemistry
For immunocytochemistry of AID bovine transfected cells, they were fixed with 4% paraformaldehyde for 15 minutes and then washed thrice with PBS-T (0.1% Tween-20 and 7.7 mM sodium azide in PBS) for 5 minutes. After being washed, the transfected cells were treated with blocking solution (5% BSA in PBS-T) for 1 hour at room temperature. The primary antibodies, ACIDA in rabbit (10 μg/mL; Abcam), OCT-3/4 in goat (1:100 dilution; Santacruz), NANOG in rabbit (1:100 dilution; Abcam), SOX2 in mouse (8 ng/mL; R&D System), SSEA-1 in mouse (1:100; Santacruz), SSEA-4 in mouse (1:100; Santacruz), Tra-1-60 in mouse (1:100; Santacruz), and Tra-1-81 in mouse (1:100; Santacruz), were added onto the cells and then incubated at 4°C overnight. The transfected cells were washed thrice with PBS-T for 5 minutes. Each primary antibody was detected using secondary antibodies (1:150 dilution; Invitrogen): Alexa Fluor® 568 Donkey Anti-Goat IgG (H + L), Alexa Fluor® 546 Goat Anti-Rabbit IgG (H + L), or Alexa Fluor® 555 Donkey Anti-Mouse IgG (H + L) at room temperature for 1 hour. The transfected cells were stained with Hoechst 33342 (5 μg/mL) at room temperature for 10 minutes and then examined under the fluorescence microscope (LEICA DM 2500; Switzerland).
Bisulfite sequencing
Genome DNAs (gDNAs) of the AID bovine transfected cells were extracted using QIAamp DNA Mini kits (Qiagen). A gDNA was treated with C:T conversion reagents from EZ DNA methylation-Gold kit (Zymo Research) for detecting the methylated cytidines. The mixed solutions were cycled at 98°C for 10 minutes and 64°C for 2.5 hours. Treated gDNAs amplified by PCR were inserted into TA vectors and then analyzed for bisulfite sequencing. Primers used for bisulfate sequencing are listed in Table 4. Sequencing data were aligned with known promoter sequences of pluripotency genes from NCBI using the BiQ analyzer and patterns of methylation of specific genes were identified.
Statistical analysis
To evaluate statistical differences in relative numbers, probability (P) values were established using one-way ANOVA with GraphPad Prism 5 (GraphPad Software, Inc.).
Results
Efficiency of transfection
To establish optimum condition of transfection of TA vector into bovine somatic cells, we have prepared 12 kinds of ratios with pCMV6-AC-IRES-GFP-Puro vector and Lipofectamine LTX as shown in Supplementary Table S1. The result of flow cytometer is shown in Supplementary Fig. S3, 1: 3 ratio of plasmid DNA and Lipofectamine LTX condition was shown as the highest (15.4%) when compared with other ratio conditions; 1: 3 ratio condition of plasmid DNA and Lipofectamine LTX was used in all transfection experiments. The colonies of transfected cells were founded on day 7 of culture of stem cell medium following the transfection. The average number of colonies was six in a six-well dish (1 × 105/10 cm2).
Expression of the AID gene in AID bovine transfected cells
The pCMV6-AC-IRES-GFP-Puro expression vector was generally designed to express the gene with a green fluorescent protein (GFP). After induction of pCMV6-AC-IRES-GFP-Puro expression vectors into bovine somatic cells, expression of green fluorescent protein was simultaneously verified in empty vector and AID expression vector bovine cells (Fig. 1). However, expression of GFP in the control was not identified. Interestingly, green fluorescent protein of the expression vector was detected only in cytoplasm of the transfected cells, whereas the nucleus of the cells was stained by propidium iodide.
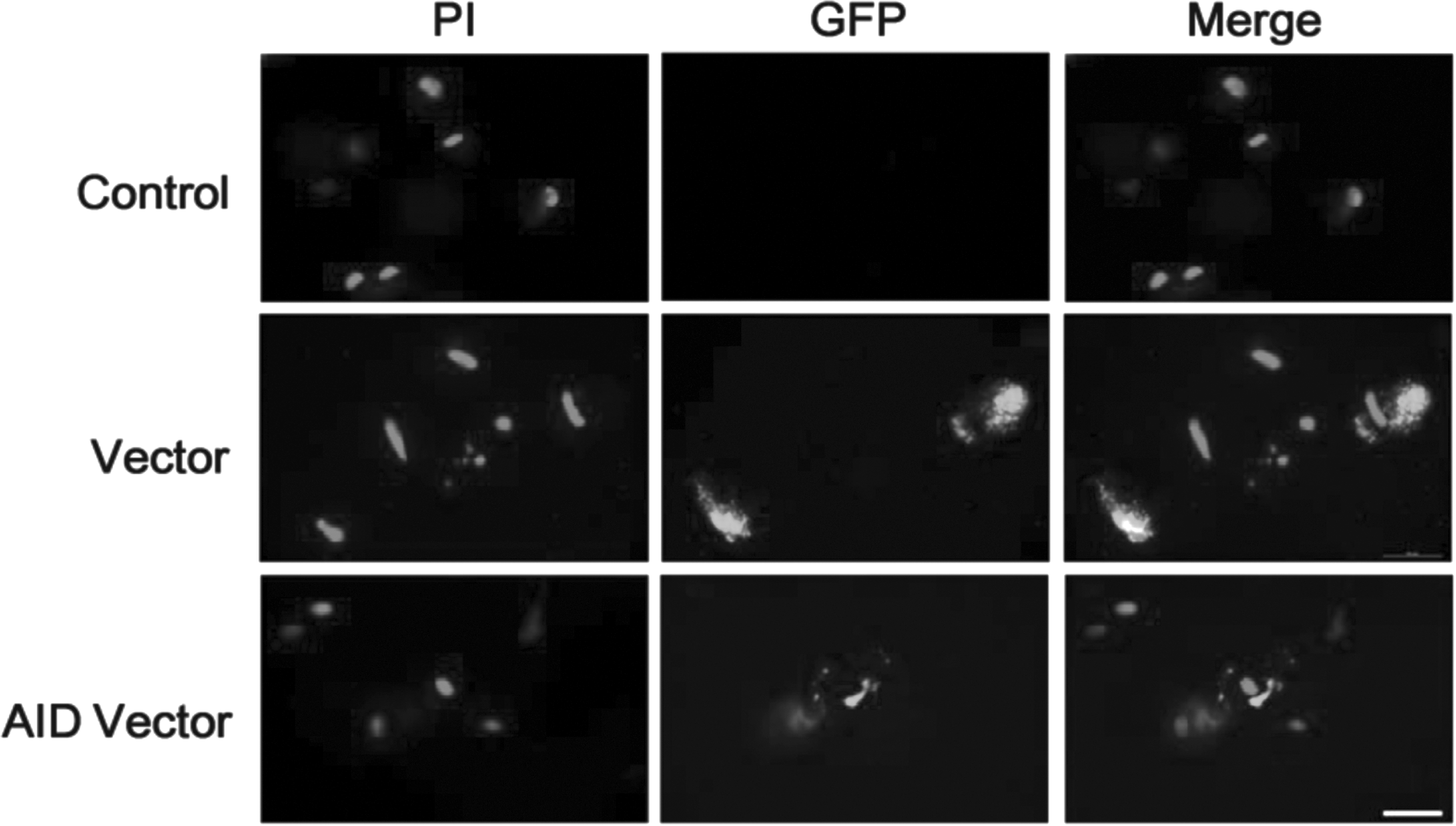
Identification of expression vectors in bovine transfected cells by immunocytochemistry assay. Bovine somatic cells were transfected with pCMV6-AC-IRES-GFP-Puro (vector) or pCMV6-AC-IRES-GFP-Puro vector with inserted AID gene (AID vector), respectively. The control was not transfected cells. Vector and AID vector-transfected cells were thoroughly selected by using puromycin (1.2 μg/mL) as a selection marker for 5–7 days. All group cells were cultured in stem cell culture medium. Scale bar = 200 μm. AID, activation-induced cytidine deaminase; GFP, green fluorescent protein as a reporter protein; PI, propidium iodide for staining the nucleus.
Morphological change in AID-transfected cells
The morphology of the bovine transfected cells was routinely observed following the induction of AID expression vector. After the transfected cells were thoroughly selected under stem cell culture medium containing puromycin for 5–6 days, they were placed on mouse embryonic feeder cells (MEFs) treated with mitomycin C. As a result, there was no morphological difference between the control, empty expression vector, and AID expression vector at day 7 of the transfection (Fig. 2a1, b1, and c1). On day 14, the morphology of AID-transfected cells had changed to long and spherical shapes unlike that of the control and empty expression vector cells (Fig. 2a2, b2, and c2). Eventually, only AID expression vector cells had started to transform into colonies at day 19 of the transfection, whereas the control and empty expression vector cells did not form a colony for the period of culture (Fig. 2a3, b3, and c3).
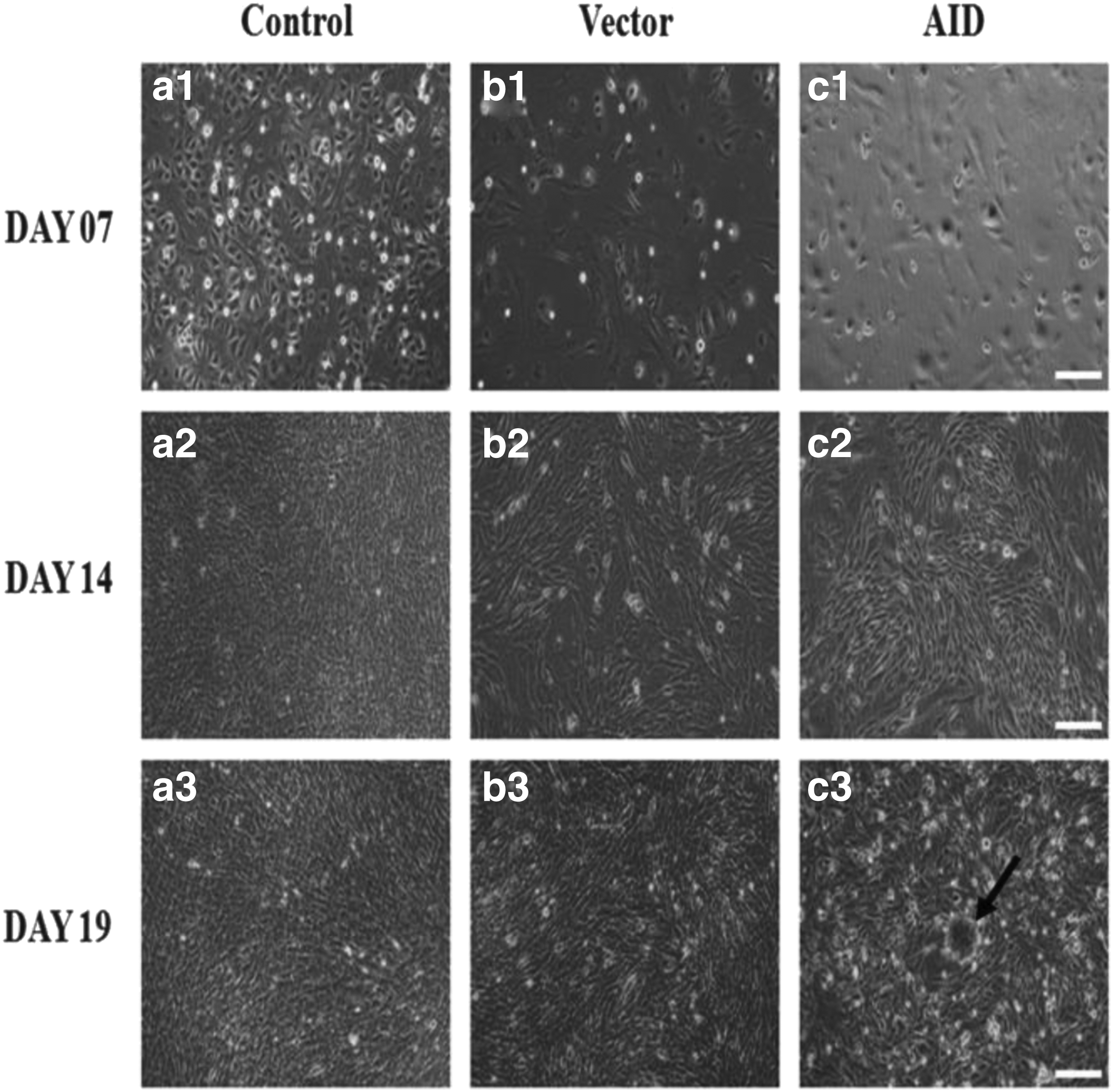
Representative morphologies of cells derived from AID bovine transfected cells.
AP staining in AID-transfected cells
Generally ES cells are characterized by high-level expression of AP as one of undifferentiated state markers. We examined the activity of AP in colonies generated from AID bovine transfected cells. As mentioned above, the colonies derived from the AID-transfected cells had begun to emerge at day 19 of the transfection (Fig. 2c3). Because AP staining of colonies derived from the AID-transfected cells was difficult at cell passage 1 after the transfection, the percentage of AP-positive colonies was calculated with the number of colony, hereafter passage 2. As shown in Figure 3A, the compact colonies shown like cell clusters were strongly stained for AP. In addition, the percentage of stained AP was calculated in the sorts of cell passages 2–3, 6–7, and 9–10, respectively (Fig. 3B). The percentage of AP-positive colonies at cell passage 9–10 was the highest compared with that of at cell passages 2–3 and 6–7.
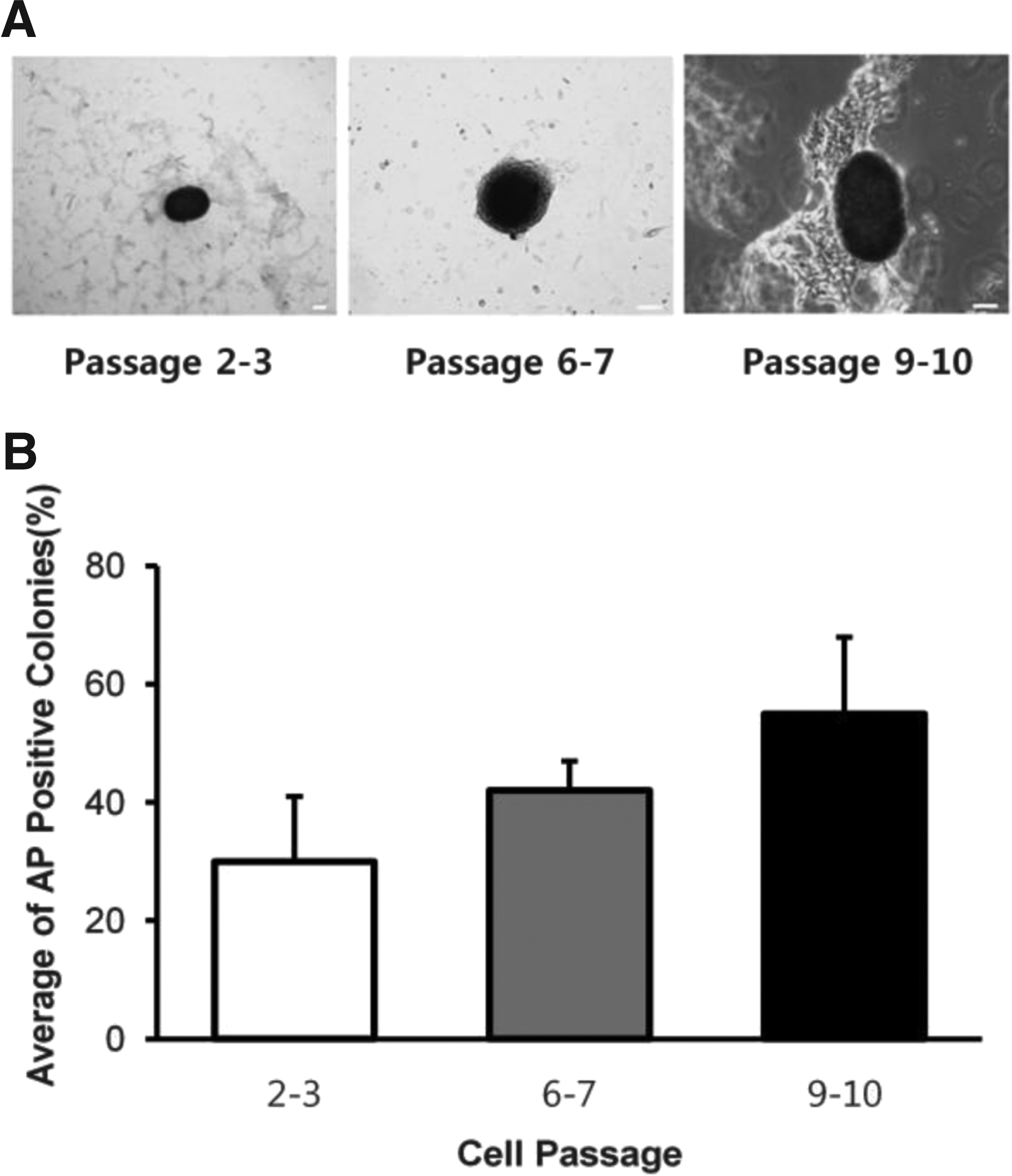
Representative morphologies and average of positive AP colonies classified by cell passages after induction of the AID gene.
Expression of pluripotency genes in AID bovine transfected cells
To investigate the expression of the AID gene and pluripotency genes such as OCT-3/4, NANOG, and SOX2 in the bovine somatic cells after induction of the AID gene, RT-PCR was performed by cDNAs reverse transcribed with mRNAs extracted from individual transfected bovine cells. The expression of the AID gene was induced only in AID-transfected cells cultured on MEFs or under feeder-free condition (w/o MEFs), whereas not detected in the control, empty vector-transfected cells, or MEFs (Fig. 4). Additionally, expression of pluripotency genes (OCT-3/4, NANOG, and SOX2) was also detected only in AID bovine transfected cells cultured with or without MEFs. On the contrary, these pluripotency genes were not expressed in the control, empty vector cells, and MEFs. GAPDH was used as the housekeeping gene. Oligonucleotide sequences are shown in Table 2.
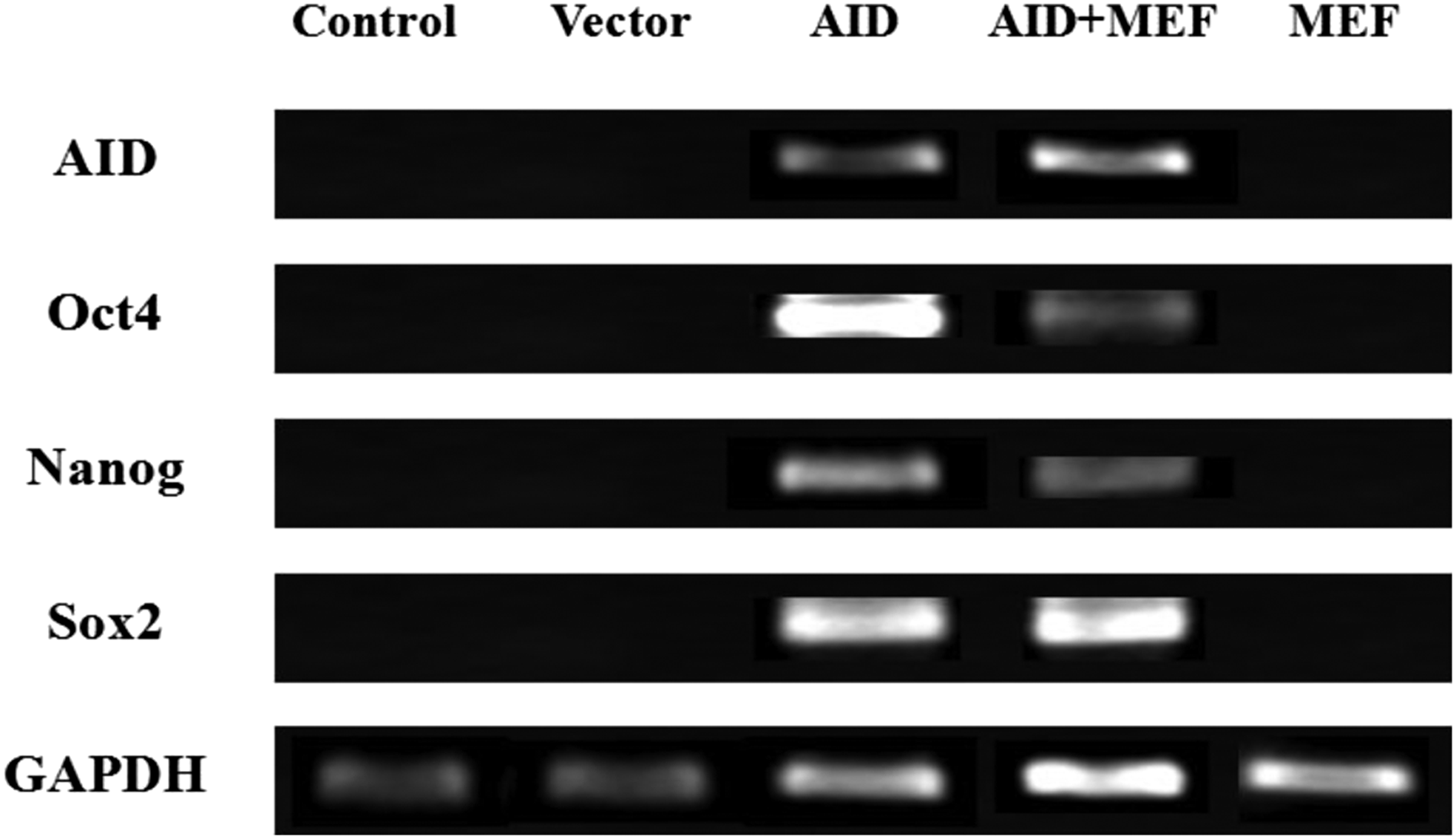
Analysis of expression patterns of pluripotency genes in AID bovine transfected cells by RT-PCR. The bovine transfected cells were selected from colonies at cell passage 6–7 after induction of the AID gene. Control: bovine somatic cells. Vector: bovine cells transfected with only pCMV6-AC-IRES-GFP-Puro expression vector. AID: bovine cells transfected with pCMV6-AC-IRES-GFP-Puro expression vector with inserted AID gene. AID + MEF: bovine cells cultured on the MEFs transfected with pCMV6-AC-IRES-GFP-Puro expression vector with inserted AID gene. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; MEF, inactivated mouse embryonic fibroblast feeder cell; RT-PCR, real-time polymerase chain reaction.
In addition, the expression level of two key pluripotency genes in the AID-transfected cells was measured by qRT-PCR. As shown in Figure 5, OCT-3/4 and NANOG of AID-transfected cells were upregulated when compared with those of the control and empty vector cells. Furthermore, the expression levels of OCT-3/4 and NANOG of the AID-transfected cells in passage 9–10 (Later) were increased higher than those of the transfected cells in passage 2–3 (Early) or 6–7 (Mid) (p < 0.05) (Fig. 5A, B). These data were analyzed with the use of 2−ΔΔCT (Livak) method, and the level of gene expression was normalized relative to that of the reference gene (GAPDH). Oligonucleotide sequences are shown in Table 3.
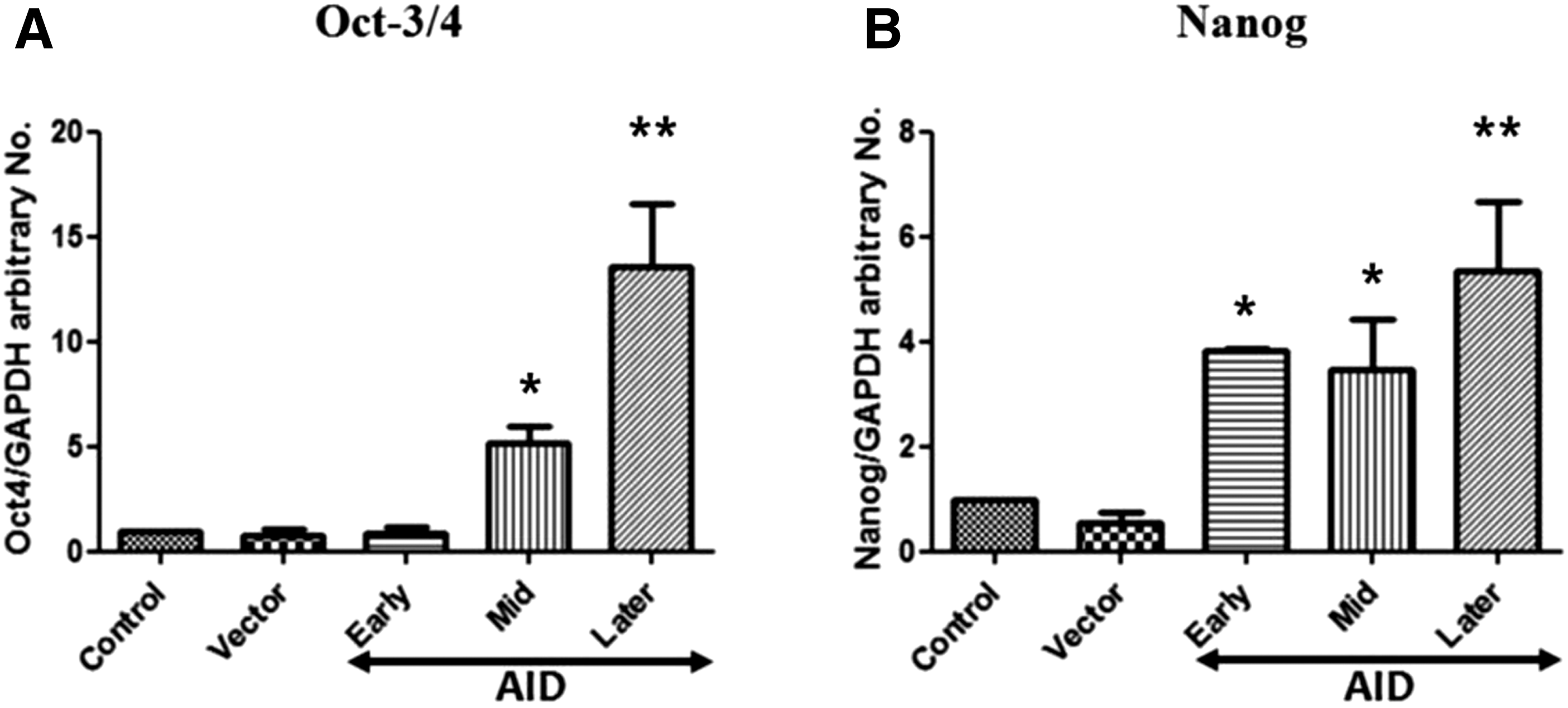
Comparison of levels of pluripotency gene expression in AID bovine transfected bovine cells by qRT-PCR analysis of OCT-3/4
Expression of AID and Pluripotency proteins in AID bovine transfected cells
To examine expression of AID protein, pluripotency proteins, and pluripotency-related antigens in the AID-transfected cells, the assay of immunocytochemistry was carried out. For immunostaining of AID protein, freshly isolated colonies at cell passage 6–7 after the transfection were fixed in 4% paraformaldehyde and then detected using anti-ACIDA antibody. As shown in Figure 6, colonies derived from AID-transfected cells were positively stained with AP. Pluripotency proteins (OCT-3/4, NANOG, and SOX2) were positively expressed within the compact spherical colonies (Fig. 7). In addition, pluripotency-related antigens (SSEA-4, Tra-1-60, and Tra-1-81) were positively stained in colonies generated from AID-transfected cells, whereas expression of SSEA-1 was not detected (Fig. 8).
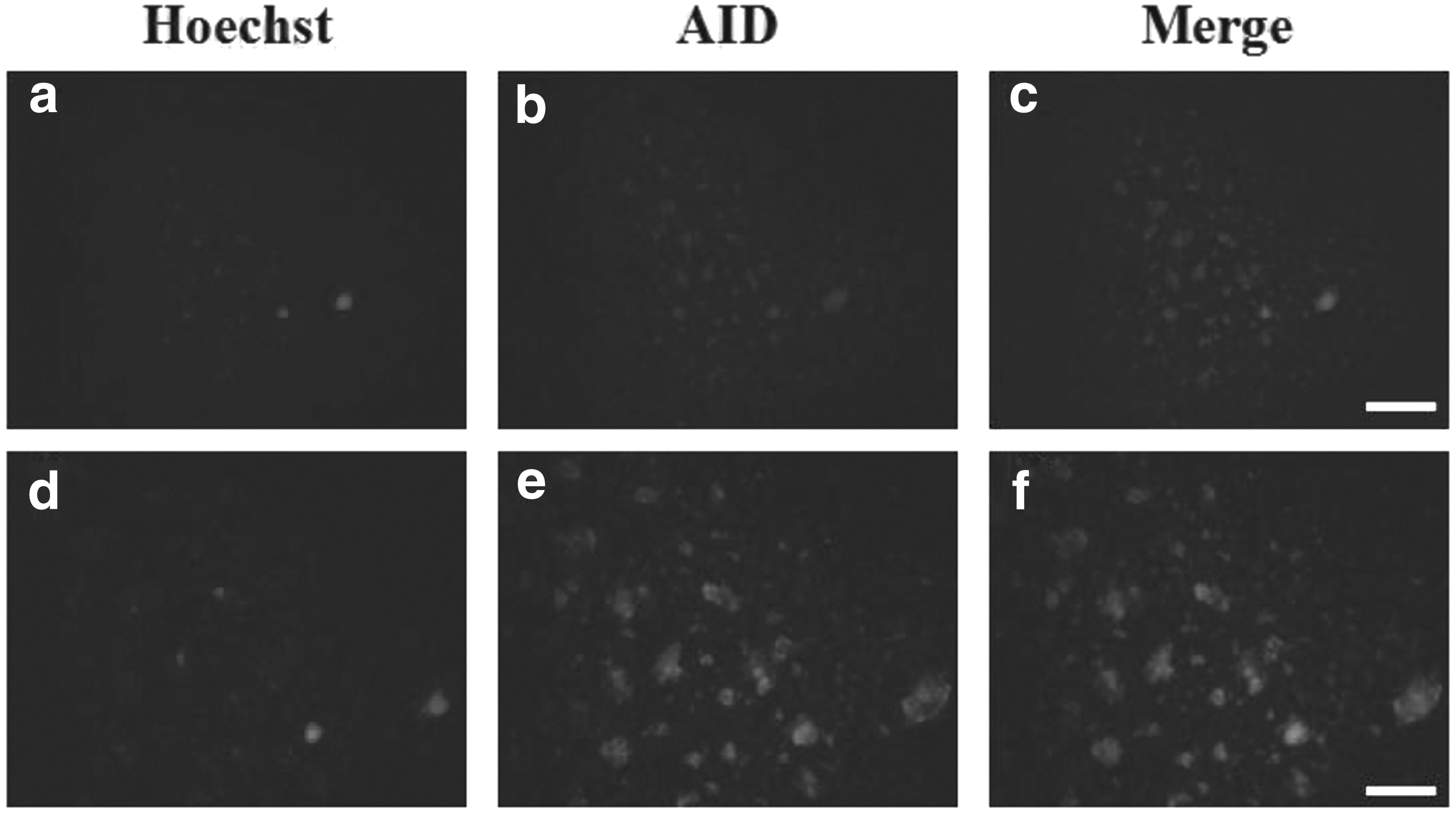
Expression of AID protein in AID-induced bovine cells. The assay of immunocytochemistry was carried out to detect expression of AID protein in AID bovine transfected cells. The transfected cells were selected at cell passage 6–7 after induction of the AID gene. Hoechst staining was performed to identify the nuclei of the cell. Hoechst 33342 staining
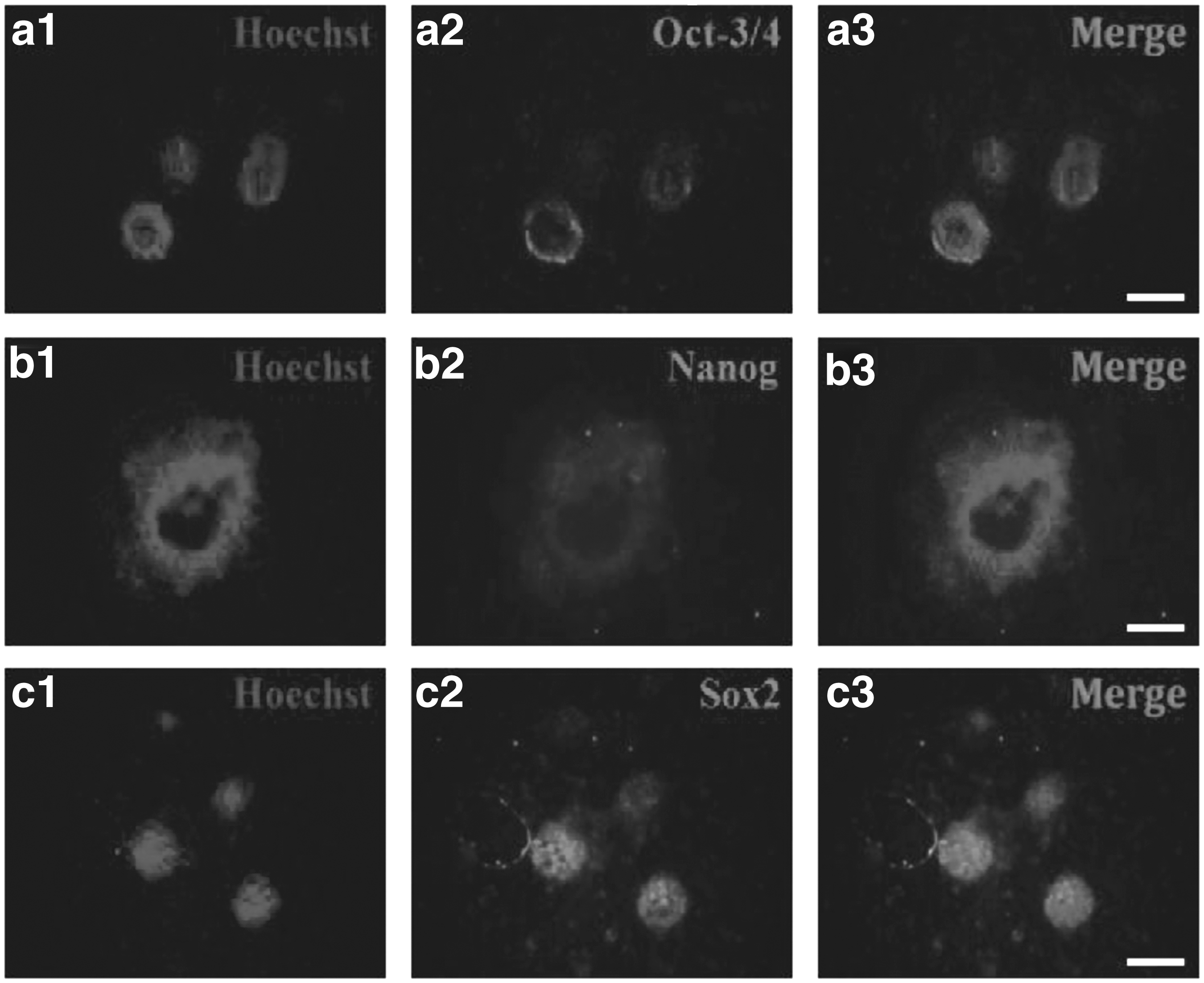
Expression of pluripotency proteins in AID-induced bovine cells. Immunocytochemistry of AID bovine transfected cells showing expression of pluripotency proteins, OCT-3/4
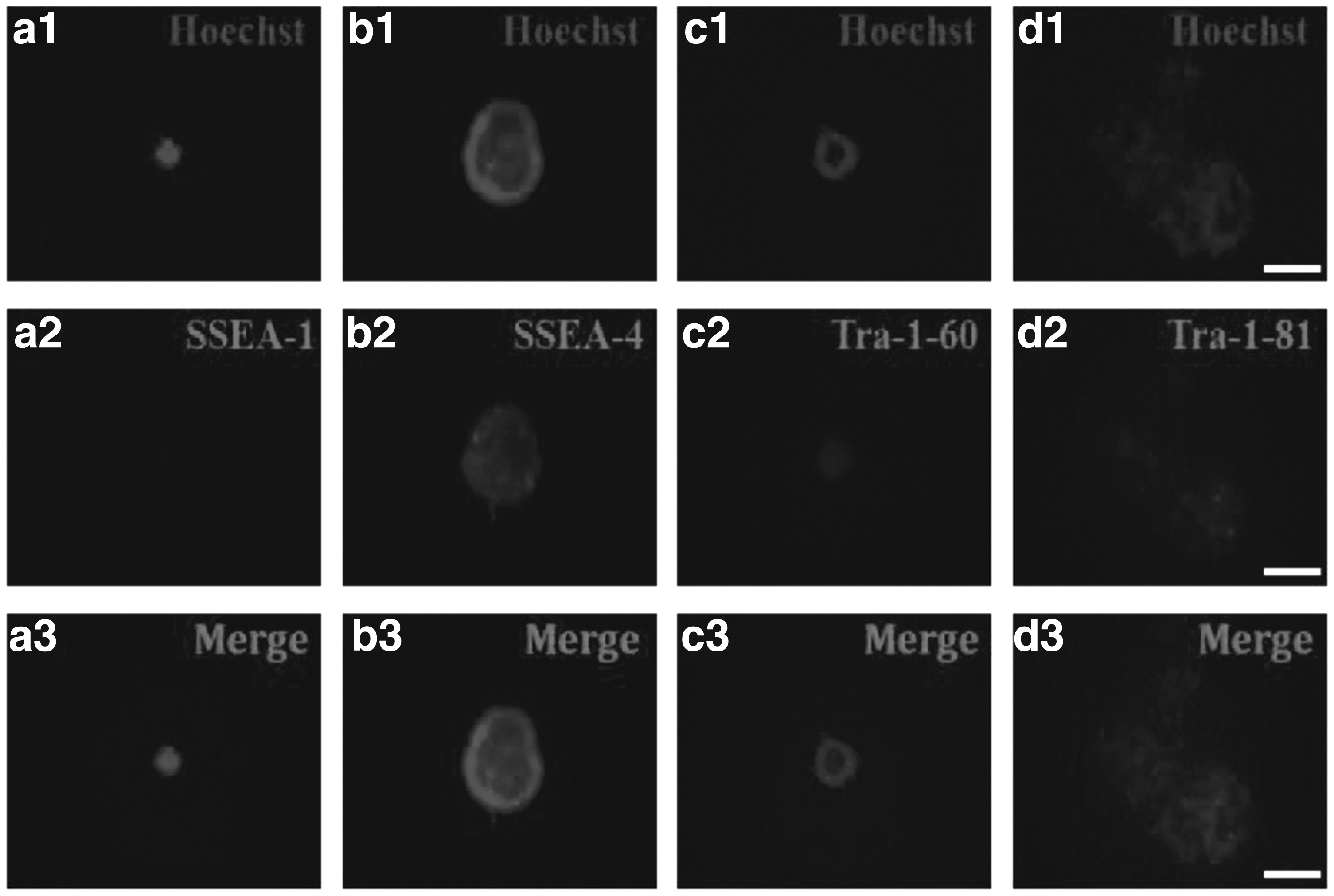
Expression of pluripotency-related antigens in AID-induced bovine cells. Immunocytochemistry of AID bovine transfected cells showing expression of pluripotency-related antigens, SSEA-1
DNA demethylation in pluripotency gene promoters of AID-transfected cells
We analyzed the state of DNA demethylation in the promoter regions of pluripotency genes (OCT-3/4, NANOG) of AID bovine transfected cells using the bisulfite sequencing method. Generally, the promoter regions of pluripotency genes were highly methylated in differentiated somatic cells when compared with those of pluripotency stem cells. As shown in Figure 9, the levels of DNA demethylation in OCT-3/4 and NANOG promoter regions of the AID-transfected cells were significantly increased when compared with that of the control (29.5% vs. 62.9% based on OCT-3/4) (58.8% vs. 88.1% based on NANOG) (p < 0.05). These data were analyzed by BiQ analyzer.
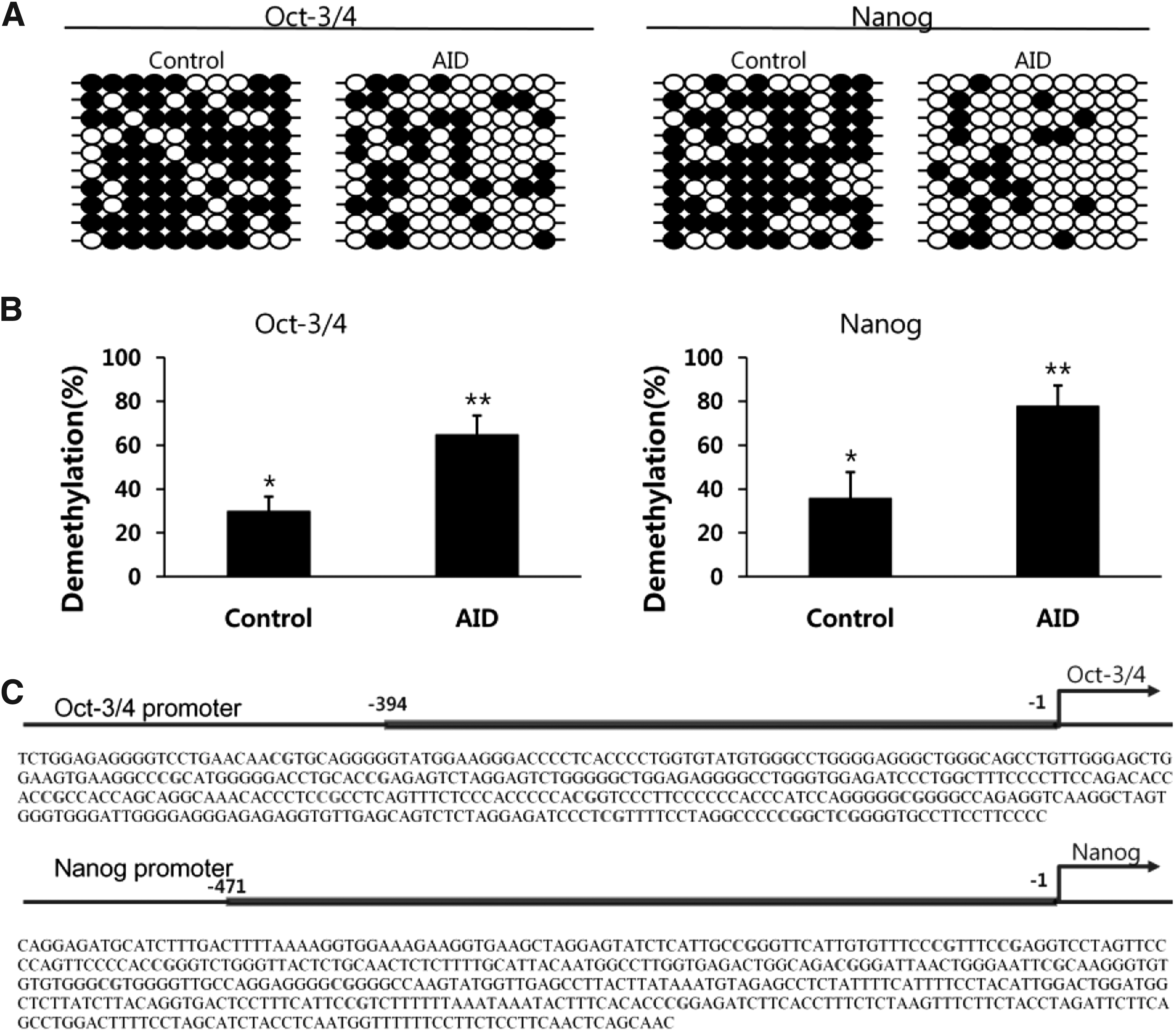
DNA demethylation in the promoter regions of pluripotency genes in AID-induced bovine cells.
Discussion
The discovery that reprogramming factors are able to convert differentiated state of a somatic cell into pluripotent one has remarkably expanded our knowledge of plasticity of the epigenome. So far, reprogramming of differentiated somatic cells toward pluripotent state can be induced by SCNT or oocyte extract method. However, incomplete reprogramming of the somatic genome or low frequency of SCNT efficiency was consistently reported (Rideout et al., 2001; Wilmut et al., 2002). In case of induced pluripotent stem cells (iPSCs), much attention has been paid to this recent technology for nuclear reprogramming. iPS cells can be induced from differentiated somatic cells transduced by only combination of transcription factors (Oct-3/4, Sox2, Klf4, and cMyc) with a retrovirus delivery system (Takahashi and Yamanaka, 2006). Even though the goal of reprogramming strategies is to generate therapeutic cells for clinical purpose, a significant obstacle is the tumorigenic potential of iPSCs (Okita et al., 2007).
Therefore, another strategy to minimize the use of tumorigenic transcription factors is required to replace with appropriate small molecules during nuclear reprogramming. The use of a histone deacetylase inhibitor for iPSCs could substitute in the absence of cMyc (Huangfu et al., 2008), and treatment with a histone methyltransferase inhibitor could induce the reprogramming of differentiated cells by using only Oct-3/4 and Klf4 transcription factors (Shi et al., 2008). In addition, the treatment of DNA methyltransferase inhibitors could improve overall efficiency of the nuclear reprogramming process (Mikkelsen et al., 2008). Recently, it was reported that enzymes such as the 10–11 translocation (Kriaucionis and Heintz, 2009; Tahiliani et al., 2009) and AID/APOBEC families (Bhutani et al., 2010; Cortellino et al., 2011; Guo et al., 2011; Popp et al., 2010) are one of the leading candidates for DNA demethylation in mammals.
Thus, in the present study, we paid attention to AID function as a demethylase for reprogramming in mammals. The AID gene is usually expressed in immune tissues (spleen, lymph node, and Peyer's patch) and plays a pivotal role in producing antibody diversity in B cells. Generally, AID induces deamination of 5-methylcytosine residues to thymine (T), which are repaired by BER system or mismatch repair (MMR). Cytosine methylation (formation of 5-methylcytosine (5mC) at CpG sites) is an important epigenetic modification by mediating transcriptional regulation, including transcriptional repression, X chromosome inactivation, and imprinting in mammals (Bird, 1992; Kass et al., 1997; Siegfried and Cedar, 1997).
The present study provided evidences of DNA demethylation induced by AID as a DNA demethylase. After transfection of the AID expression vector (pCMV6-AC-IRES-GFP-Puro) into bovine differentiated cells, the transfected cells were cultured on MEFs under stem cell culture condition. AID-transfected cells began to form colonies at day 19 of the transfection (Fig. 2c3), whereas no colony formations were formed in the absence of AID. Additionally, the AID gene and protein were expressed in only AID-transfected cells; however, the control and empty expression vector cells were not expressed. Next, we observed whether the AID bovine transfected cells can induce the characteristics of pluripotency such as the activity of AP and expression of pluripotency genes (OCT-3/4, NANOG, and SOX2) as important regulators in pluripotent cells. In the present experiment, induction of AID in the differentiated cells could induce both positive AP staining (Figs. 3 and 6) and OCT-3/4, NANOG, and SOX2 gene expression (Figs. 4 and 5). These results were quite similar to that of heterokaryon experiment (fused mouse ES cells and human fibroblasts), suggesting that human OCT-3/4 and NANOG transcripts were detected immediately after fusion (Bhutani et al., 2010).
However, in contrast with heterokaryons, the onset of expression of two key pluripotency genes in AID-induced bovine cells was delayed until colonies had formed (Fig. 5). It suggests that AID protein derived from the expression vector is bound to silent methylated OCT-3/4 and NANOG promoters and removes the methylated pluripotency promoter sites of differentiated somatic cells. In the different approaches to reprogramming, at least in some respects, timing difference existed. In Fig 3B, the percentage of AP-positive clones is highest in passage 9–10 (Later) and lowest in passage 2–3 (Early). However, the expression of pluripotent genes (OCT-3/4 and NANOG) was highest in passage 9–10 (Later), lower in passage 6–7 (Mid), and lowest in passage 2–3 (Early) according to Figure 5.
In addition, the result of immunocytochemistry assay of AID-transfected cells was synchronized with that of pluripotency gene expression. Protein expression of the key pluripotency genes (OCT-3/4, NANOG, and SOX2) was positively identified in only colonies derived from AID-induced bovine cells, but not in individual cells (Fig. 7). Additionally, pluripotency-related antigens (SSEA-4, Tra-1-60, and Tra-1-81) were detected in colonies derived from AID-induced bovine cells, but SSEA-1 was not detected (Fig. 8). Interestingly, the cell surface markers expressed in AID-induced bovine cells are quite similar to those in human embryonic germ cells, ES cells, and embryonal carcinoma (EC) cells. On the other hand, SSEA-1 marker antigen is usually expressed in only murine embryos, ES cells, and EC cells (Zhao et al., 2012).
Finally, the levels of DNA demethylation in OCT-3/4 and NANOG promoter regions of AID-induced bovine cells were significantly increased when compared with that of the control (29.5% vs. 62.9% in OCT-3/4) (58.8% vs. 88.1% in NANOG). Bhutani et al. (2010) presented that the levels of DNA methylation were decreased in the promoter regions of pluripotency genes (OCT-3/4 and NANOG) using siRNA-mediated knockdown. Furthermore, the induction of reprogramming of mouse fibroblasts to iPSCs was arrested and the frequency highly reduced in the absence of AID using different shRNAs (Bhutani et al., 2013). Reprogramming efficiency on AID overexpression improves (relatively modest) twofold when AID acts in conjunction with thymine-DNA (TDG) that has been implicated in the mammalian DNA demethylation pathway (Bhutani et al., 2011; Branco et al., 2012).
However, the injection of a methylated DNA fragment into zebrafish embryos did not affect either the methylation of genomic DNA or the expression level of aid, which has been shown to play a role in demethylation. Aberrant methylation was not observed at certain CpG islands following the injection of antisense morpholino oligonucleotides against AID and MBD4 (Shimoda et al., 2014). The role of AID in DNA demethylation is controversial because there is currently no evidence to support proposed roles of AID in DNA demethylation in Zebrafish embryos (Nabel et al., 2012; Shimoda et al., 2014).
Taken together, we demonstrated that DNA demethylation of pluripotency gene promoters and expression of pluripotency genes (OCT-3/4 and NANOG) are induced in bovine differentiated somatic cells after transfection of the AID gene. This strategy of nuclear reprogramming using this enzyme (AID) closely associated with DNA demethylation may greatly contribute to understand the plasticity of the epigenome.
Footnotes
Acknowledgments
This work was partly supported by the National Research Foundation of Korea funded by the Korean Government (NRF-2011-0013703) and Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ009418022015), Rural Development Administration, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.