
Abstract
Abstract
The aim of this study was to assess the effect of insulin-like growth factor-1 (IGF-1) on the developmental competence of somatic cell nuclear transfer (SCNT) bovine embryos. First, the expression levels of IGF-1 receptor (IGF-1R) and IGF-1 in the oocytes and embryos of different developmental stages were examined. Then the effects of exogenous IGF-1 on the development of SCNT embryos were evaluated both in vitro and in vivo. The results showed that IGF-1 was not expressed in both IVF and SCNT embryos, whereas IGF-1R could be detected throughout the preimplantation stages in both protein and mRNA levels. Also, exogenous IGF-1 had no obvious impact on the developmental competence of IVF embryos. However, it could improve the developmental competence of SCNT embryos in terms of blastocyst developmental rate (31.3% vs. 43.2%, p < 0.05), total cell number (93.0 ± 9.9 vs. 101.0 ± 9.8, p < 0.05), ratio of inner cell mass (ICM) to trophectoderm (TE) (0.29 ± 0.006 vs. 0.39 ± 0.005, p < 0.05), and apoptosis index in day 7 blastocysts (2.5 ± 0.22 vs. 8.7 ± 0.41, p < 0.05) compared to the control group. Although no statistical difference in pregnancy rate and birth rate was observed after embryo transfer, there was an upward tendency in both examined terms in the IGF-1–supplemented group when compared with the control group. In conclusion, the present study showed that supplementing exogenous IGF-1 to the culture medium has an obvious positive effect on the development competence of SCNT embryos.
Introduction
T
Modified synthetic oviduct fluid (mSOF) is an appropriate culture medium for in vitro production (IVP) of bovine embryos, and when mSOF is used as the culture medium, as high as ∼40% of the IVP bovine embryos can develop to blastocysts. However, the quality of the IVP embryos derived from mSOF is still unsatisfactory, and the in vivo developmental competence is obviously lower than that of in vivo–derived embryos. Thus, optimizing the culture medium is a key step for improving the quality of IVP embryos. Ingredient analysis showed that there are numerous growth factors in oviduct fluid. Moreover, some recipient proteins of these growth factors can also be detected in the preimplantation embryos. In addition, supplementing culture medium with some growth factors, such as plasmid-encoded granulocyte-macrophage colony-stimulating factor (pGM-CSF) (Kwak et al., 2012), epidermal growth factor (EGF) (Wani et al., 2012), and insulin-like growth factor-I (IGF-1) (Dhali et al., 2009), has obvious positive effects on the developmental potential of IVP embryos. These studies indicated that supplementing culture medium with growth factors might be a promising way to optimize culture medium.
The influence of IGF-1 on embryonic development is controversial. Numerous studies have shown that IGF-1 is an important growth factor for preimplantation embryo development, and it can improve the developmental competence of an IVF embryo by enhancing cell proliferation and mitogenesis and regulating apoptosis in various species (Lin et al. 2003; Makarevich and Markkula, 2002; Zhandi et al., 2009). However, some studies have reported that there was no obvious positive effect of IGF-1 on the in vitro and in vivo developmental competence of IVF embryos (Block and Hansen, 2007).
The widely acceptable explanation about these conflicting results is that the culture system and (or) embryonic status may be important factors influencing the effect of IGF-1. Such factors might exert positive effects in embryos undergoing adverse conditions, such as heat stress, a high oxygen environment, and suboptimal culture system, but no obvious effects on embryos in optimal condition (Block and Hansen, 2007). Compared to IVF embryos, most SCNT embryos have a weaker developmental status. In addition, previous studies have shown that numerous physical activities, including nutrition requirement, mRNA expression, and cellular differentiation, are different between IVF- and SCNT-derived embryos (Bauersachs et al., 2009; Niemann et al., 2002; Yamanaka et al., 2009). Although many investigations about the impact of IGF-1 on the development of IVF embryos have been carried out, there is little information about the effect of IGF-1 on the development of SCNT embryos. Thus, information about effects of IGF-1 on IVF embryos should be re-evaluated in SCNT embryos.
To improve the developmental competence of bovine SCNT embryos, the present study was carried out to explore the effects of exogenous IGF-1 on the developmental competence of both IVF and SCNT embryos. In addition, the expression patterns of IGF-1 and the IGF-1 receptor (IGF-1R) in both IVF- and SCNT-derived preimplantation embryos were assessed and compared.
Materials and Methods
Unless otherwise indicated, all reagents were purchased from Sigma (St. Louis, MO, USA). Procedures were approved by the Animal Care and Use Committee of Northwest A & F University and performed in accordance with animal welfare and ethics.
Oocytes collection and in vitro maturation
Bovine ovaries were collected from a local abattoir and transported to the laboratory in 0.9% sterile saline within 3–4 h at 15–20°C (Wang et al., 2011b). Ovaries were washed several times in prewarmed 0.9% sterile saline. The follicle liquid and cumulus–oocyte complexes (COCs) were retrieved from obvious follicles (2–8 mm in diameter) with a 10-mL syringe. Under the stereomicroscope, COCs surrounded with at least three intact layers of unexpanded cumulus cells and homogeneous cytoplasm were collected in phosphate-buffered saline (PBS) supplemented with 5% (vol/vol) fetal bovine serum (FBS). After three washes in Oocyte Maturation Medium [Tissue Culture Medium-199 (TCM-199) supplemented with 10% FBS, 1 μg/mL 17 β-estradiol, 0.075 IU/mL human menopausal gonadotropin, and 30 ng/mL EGF], about 100 COCs were placed in a 35-mm petri dish containing 4 mL of Oocyte Maturation Medium and incubated at 38.5°C in a humidified incubator of 5% CO2 in air for 20–22 h.
In vitro fertilization
Most cumulus cells of COCs were removed with a pipette, and only one or two layers of cumulus cells were left. COCs were rinsed three times in fertilization medium [BO medium supplemented with 6 mg/mL bovine serum albumin (BSA)], and transferred into 100-μL fertilization droplets (30 COCs/droplet) covered with mineral oil equilibrated for at least 8 h at 38.5°C in 5% CO2 in air. Bovine sperm stored in liquid nitrogen were thawed in water at 37°C and washed in BO medium (4 mL) via swimming up for 30 min. About 2 mL of liquid supernatant containing this sperm was collected in a 15-mL centrifuge tube, centrifuged at 1000 × g for 10 min, and resuspended in BO medium to a concentration of 2 × 106/mL. Sperm suspension solution was added to the fertilization droplets. The oocytes and sperm were incubated at 38.5°C in a humidified incubator of 5% CO2 in air for at least 8 h. All cumulus cells were denuded by treatment with 0.2% bovine testicular hyaluronidase in PBS.
Somatic cell nuclear transfer
SCNT was performed as described by Wang et al. (2011a). Briefly, after maturation, cumulus cells were removed by treatment with 0.2% bovine testicular hyaluronidase in PBS. Oocytes with a polar body were selected for SCNT. Oocytes were enucleated by aspirating the first polar body and a small amount of surrounding cytoplasm using a beveled glass pipette with 20-μm internal diameter in PBS supplemented with 7.5 μg/mL cytochalasin B and 10% FBS. Successfully enucleated oocytes were selected under a fluorescence microscope after staining with Hoechst 33342. A donor cell from generations 2–5 and cultured in serum-starved medium (0.5% FBS) for 2 days was injected into the perivitelline space of the successfully enucleated oocytes and then fused using two closely spaced electrical pulses of 35 V for 10 μsec. The reconstructed embryos were kept in mSOF containing 5 μg/mL cytochalasin B for 2 h before activation. The reconstructed embryos were then activated in 5 μM ionomycin for 3–5 min, followed by 4 h of exposure to 1.9 mM 6-dimethylaminopurine.
In vitro culture
SCNT embryos and IVF embryos were washed three times in mSOF with amino acids (mSOFaa) and finally cultured in 100-μL droplets (20 embryos/droplet) of mSOFaa supplemented with or without 100 ng of IGF-1 according to the experimental design. The culture condition was humidified atmosphere of 5% CO2 in air and 38.5°C. Medium was renewed at 72 h (day 3) by half-changing the medium. Development to two-cell embryos and blastocysts was monitored at 48 h and 168 h of culture, respectively (0 h being the time embryos were transferred to mSOFaa).
Immunofluorescence
Embryos were fixed in 4% (vol/vol) paraformaldehyde in PBS for 2 h at room temperature, permeabilized with 0.2% (vol/vol) Triton-X in PBS for 20 min at room temperature, and blocked in blocking liquid (cat. no. P0102, Beyotime, Shanghai, China) overnight at 4°C. Afterward, embryos were incubated overnight with the first anti-IGF-1R antibody from rabbit (cat. no. ab90657, Abcam, Cambridge, MA, USA) at a concentration of 2.5 μg/mL in primary dilution or anti-CDX2 mouse monoclonal antibody (CDX2 is a gene specifically expressed in TE). After incubation, embryos were washed in 0.1% PBS-polyvinyl alcohol (PVA), and treated with secondary antibodies of Alexa Fluor 555–labeled goat anti-rabbit immunoglobulin G (IgG; cat. no. A0452, Beyotime) for 2 h at room temperature. The secondary antibodies were diluted 1:500 (vol/vol) with Immunol Staining Secondary Antibody Dilution Solution (cat. no. P0108, Beyotime). After being washed three times in 0.1% PBS-PVA for 5 min each time, nuclear labeling of embryos was performed with 4′,6-diamidino-2-phenylindole hydrochloride (DAPI; Vysis Inc., Downers Grove, IL, USA) for 3 min and washing three times in 0.1% PBS-PVA for 5 min each. Finally, specimens were examined by epifluorescence using a Nikon Eclipse Ti-S microscope (Nikon, Tokyo, Japan). All images were captured using a Nikon DS-Ri1 digital camera and saved in TIFF format. The nuclei were identified by their blue fluorescence; IGF-1R or TE (CDX2) was red. The experiments were replicated three times. In each replication, 10–15 embryos per group were processed.
Quantitative real-time PCR
Thirty embryos of each developmental stage were used for quantitative real-time PCR (qPCR). The reactions were carried out in triplicate with the same pool of embryos, and experiments were replicated three times. The cDNA of oocytes and all stages of IVF and SCNT embryos cultured in mSOF were isolated under the Invitrogen SuperScript™ III Cells Direct cDNA Synthesis System according to the manufacturer's protocol. Matured oocytes and IVF or SCNT embryos at different stages, including one-cell, two-cell, four-cell, eight-cell, 16-cell, morula, and blastocyst, were used to collect mRNA. The qPCR was carried out according the guidance of Bustin et al. (2009). The amounts of IGF-1 and IGF-1R in the bovine embryos were calculated relatively. Two housekeeping genes, β-actin and Histone 2a.2 (H2a.2), were used as reference genes.
All qPCR reactions were performed in triplicate in a 20-μL reaction volume on the quantitative real-time PCR StepOne Plus System (ABI, Carlsbad, CA, USA) using 0.4 μL of ROX (50×), 10 μL of SYBR Green I (2×), 0.8 μL of F-primer, 0.8 μL of R-primer embryos, 2 μL of cDNA, and 6 μL of dH2O. The PCR protocol included 95°C 30 sec, followed by 40 cycles of 95°C 5 sec, 60°C 30 sec, and 72°C 30 sec. Primer sequences, annealing temperature, and the lengths of the amplified fragments of all transcripts are shown in Table 1.
H2A.2, histone 2A.2; IGF-1, insulin-like growth factor-1; IGF-1R, IGF-1 receptor.
Detection of apoptosis in blastocysts
The apoptotic index of day 7 blastocysts was examined by the Dead End™ Fluorometric TUNEL System (Promega, Madison, WI, USA) according to the manufacturer's protocol. Blastocysts were fixed in 4% paraformaldehyde for 2 h at room temperature and permeabilized in 0.5% Triton X-100 for 5 min. After being balanced in equid buffer for 5–8 min, they were incubated with [[[fluorescein isothiocyanate (FITC)-conjugated 2′-deoxyuridine, 5′-triphosphate (dUTP) (5 μL), terminal deoxynucleotidyl transferase (1 μL), and equid buffer (45 μL) at 37°C for 1 h in the dark (hereafter, all manipulations were performed in the dark). The tailing reaction was terminated in 2× saline sodium citrate (SSC; 0.15 mol/L sodium chloride, 0.015 mol/L sodium citrate) for 15 min.
The embryos were incubated in PBS containing 25 μg/mL RNase A and 25 μg/mL propidium iodide (PI) for 30 min, washed three times with PBS-PVA for 5 min, and mounted on slides for observation under a Nikon Eclipse Ti-S microscope (Nikon, Tokyo, Japan). All images were captured using a Nikon DS-Ri1 digital camera. The nuclei were identified by their red fluorescence, and nuclear DNA fragmentation labeled with FITC appeared green.
Embryo transfer
A healthy local cow at 2–5 years age was used as the SCNT embryo recipient. Two blastocysts were loaded in one 0.25-mL straw and transported from the laboratory to the experimental farm in HEPES-buffered TCM-199 supplemented with 10% FBS at 37°C within 1 h. The embryos were transferred nonsurgically into the uterine horn of the recipients on day 7 of their natural estrous cycle. Pregnancy was detected using rectal palpation/ultrasonography at 90 days of gestation.
Experimental design
In experiment 1, the metaphase II (MII) oocytes and embryos (one-cell, two-cell, four-cell, eight-cell, 16-cell, morula, and blastocyst) produced by IVF and SCNT were collected for qPCR, and expression levels of IGF-1 and IGF-1R were examined.
In experiment 2, the protein expression of IGF-1R was examined in bovine germinal vesicle (GV) oocytes, MII oocytes, and IVF and SCNT embryos at different developmental stages, including two-cell, four-cell, eight-cell, 16-cell, morula, and blastocyst.
In experiment 3, the effects of exogenous IGF-1 (100 ng/mL) (Bonilla et al., 2011) on the development of IVF and SCNT embryos were evaluated. The quality of embryos was assessed in terms of cleavage rate, blastocyst formation rate, the ratio of ICM to TE, apoptosis cell number in day 7 blastocysts, and in vivo development.
Statistical analysis
Experiments were repeated at least three times, and each replicate of the experiments was carried out using oocytes matured on the same day to remove any batch effect of oocytes. All embryos were allocated randomly to each treatment group. All data were tested by one-way repeated-measures analysis of variance (ANOVA) using SPSS 13.0 software. Differences were considered significant at p < 0.05.
Results
Expression patterns of IGF-1and IGF-1R in bovine oocytes, IVF, and SCNT embryos
As shown in Figure 1, the expression of IGF-1 was not detected in matured oocytes and IVF and SCNT embryos. The expression of IGF-1R was observed in MII oocytes and IVF and SCNT embryos throughout the preimplantation stages. In addition, no significant difference was observed in the expression level of IGF-1R at each developmental stage between SCNT and IVF embryos (p > 0.05).
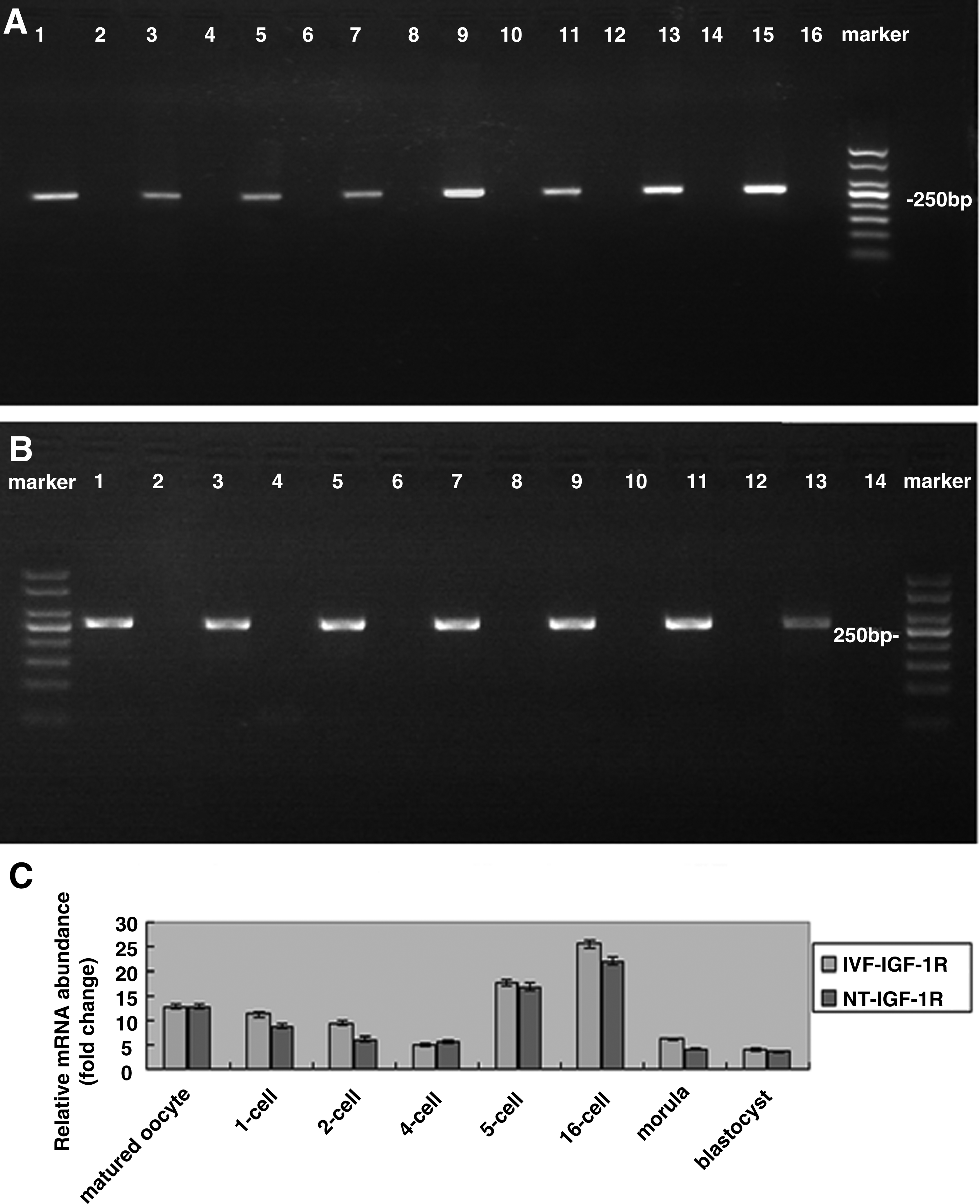
mRNA expression of IGF-1 and IGF-1R in bovine MII oocytes and preimplantation embryos produced by IVF and SCNT. (
As shown in Figure 2, the IGF-1R protein could not be observed in GV oocytes, but could be detected in the cumulus cells around GV oocytes and also could be detected in MII oocytes and all developmental stages of preimplantation embryos in both IVF and SCNT embryos.
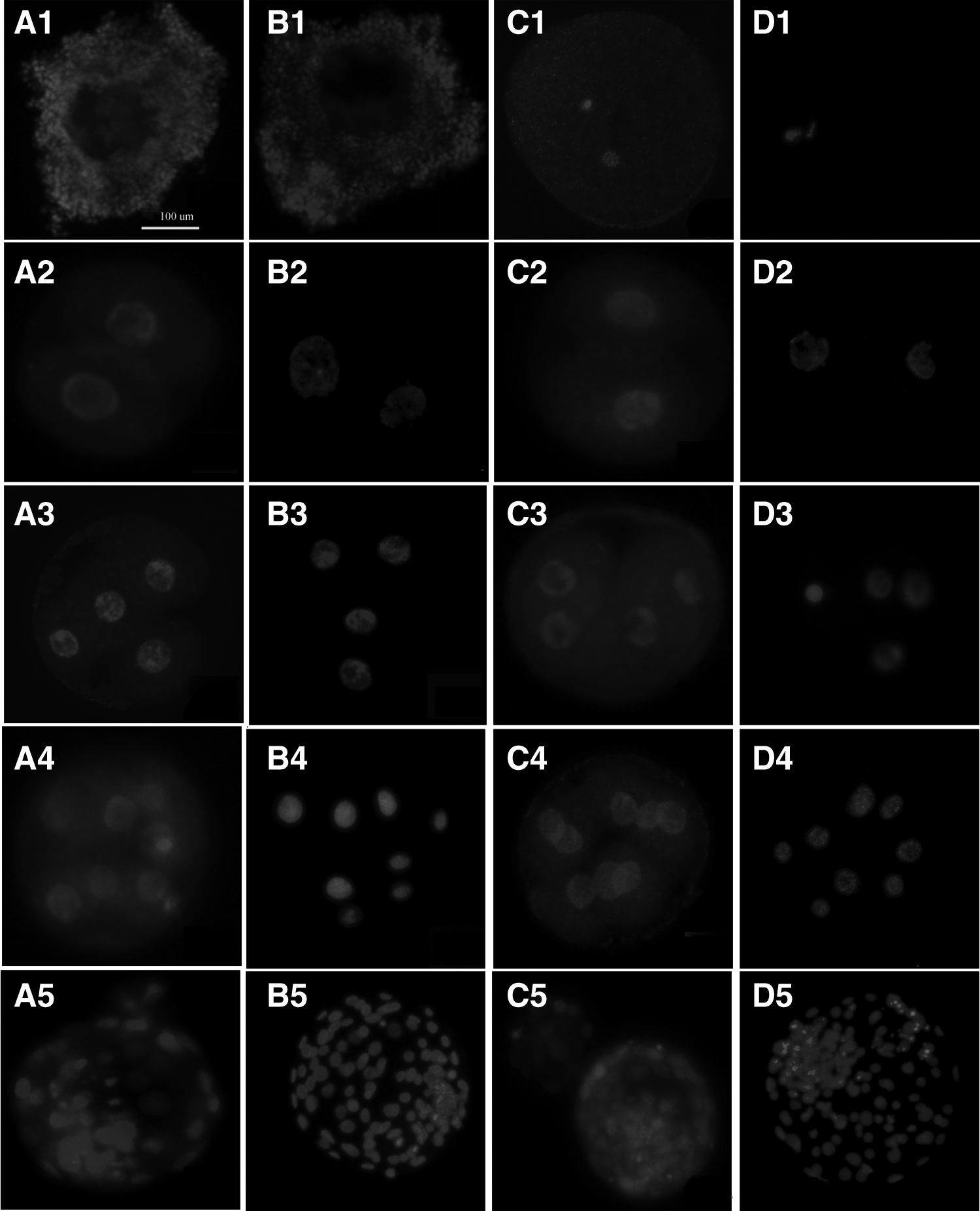
Expression of IGF-1R in bovine oocytes and preimplantation embryos produced by IVF and SCNT. (
Effects of exogenous IGF-1 on development of IVF and SCNT embryos
Exogenous IGF-1 had no obvious impact on the cleavage rate, blastocyst rate (Table 2), apoptotic index (Table 3 and Fig. 3), and allocation of ICM and TE (Table 4 and Fig. 4) of IVF embryos. However, exogenous IGF-1 could improve the developmental competence of SCNT embryos in terms of blastocyst developmental rate (31.3% vs. 43.2%, p < 0.05), total cell number (93.0 ± 9.9 vs. 101.0 ± 9.8, p < 0.05), ratio of in ICM to TE (0.29 ± 0.006 vs. 0.39 ± 0.005, p < 0.05), and apoptosis index in day 7 blastocysts (2.5 ± 0.22 vs. 8.7 ± 0.41, p < 0.05) compared with the control group. In addition, as shown in Table 2, although no statistical difference in pregnancy rate (34.4% vs. 41.5%, p > 0.05) and birth rate (14.3% vs. 26.4%, p > 0.05) and cloning efficiency (22.64% vs. 12.5%, p > 0.05) and birth weight (59.35 ± 1.91 kg vs. 60.04 ± 1.31 kg, p > 0.05) was observed between the IGF-1–supplemented group and the control group, there was an upward tendency in the means of both examined terms in IGF-1–supplemented group.
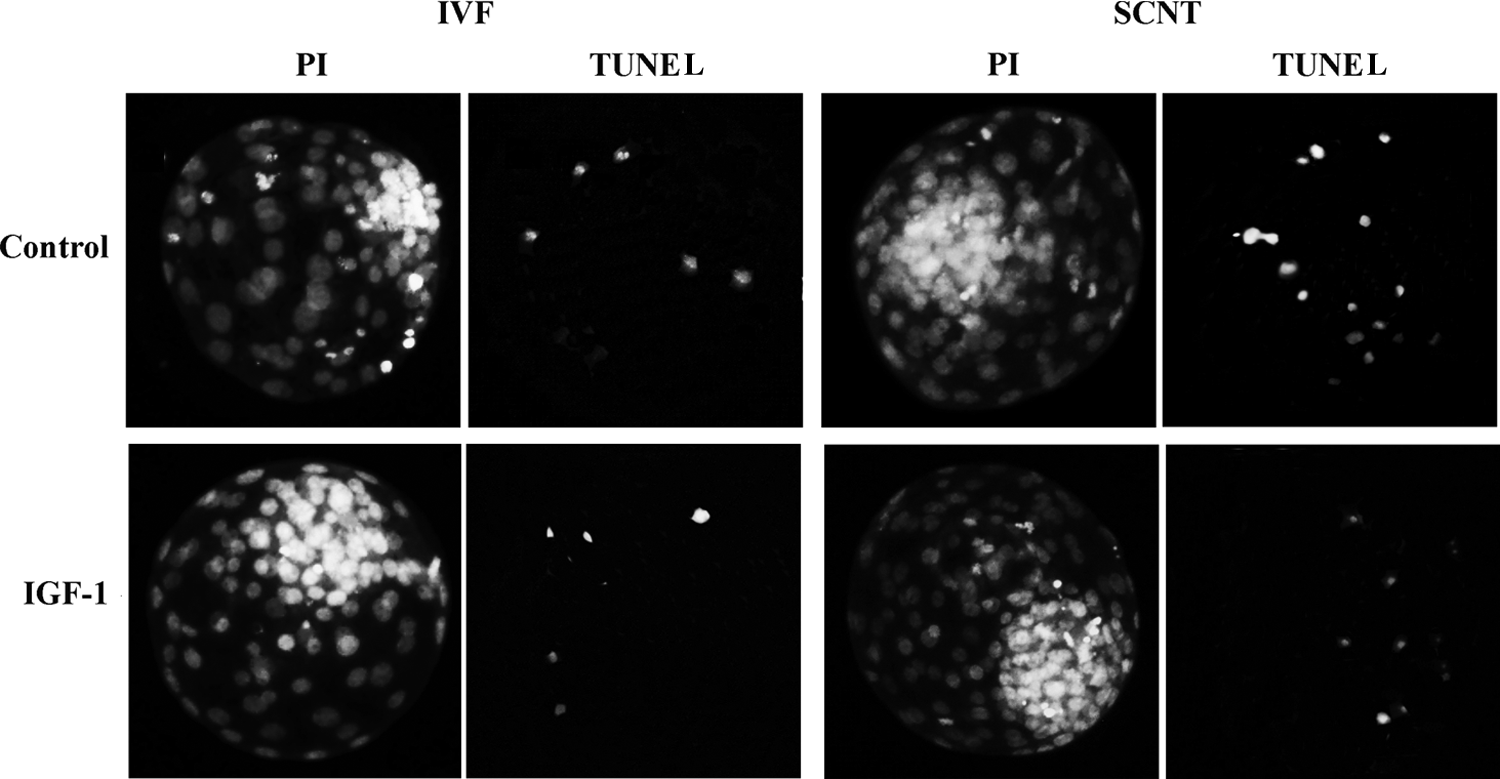
Incidence of apoptosis in blastocysts. The apoptotic blastomeres in day 7 blastocysts were detected by terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay. Each sample was counterstained with PI to detect DNA. Original magnification, 200×.
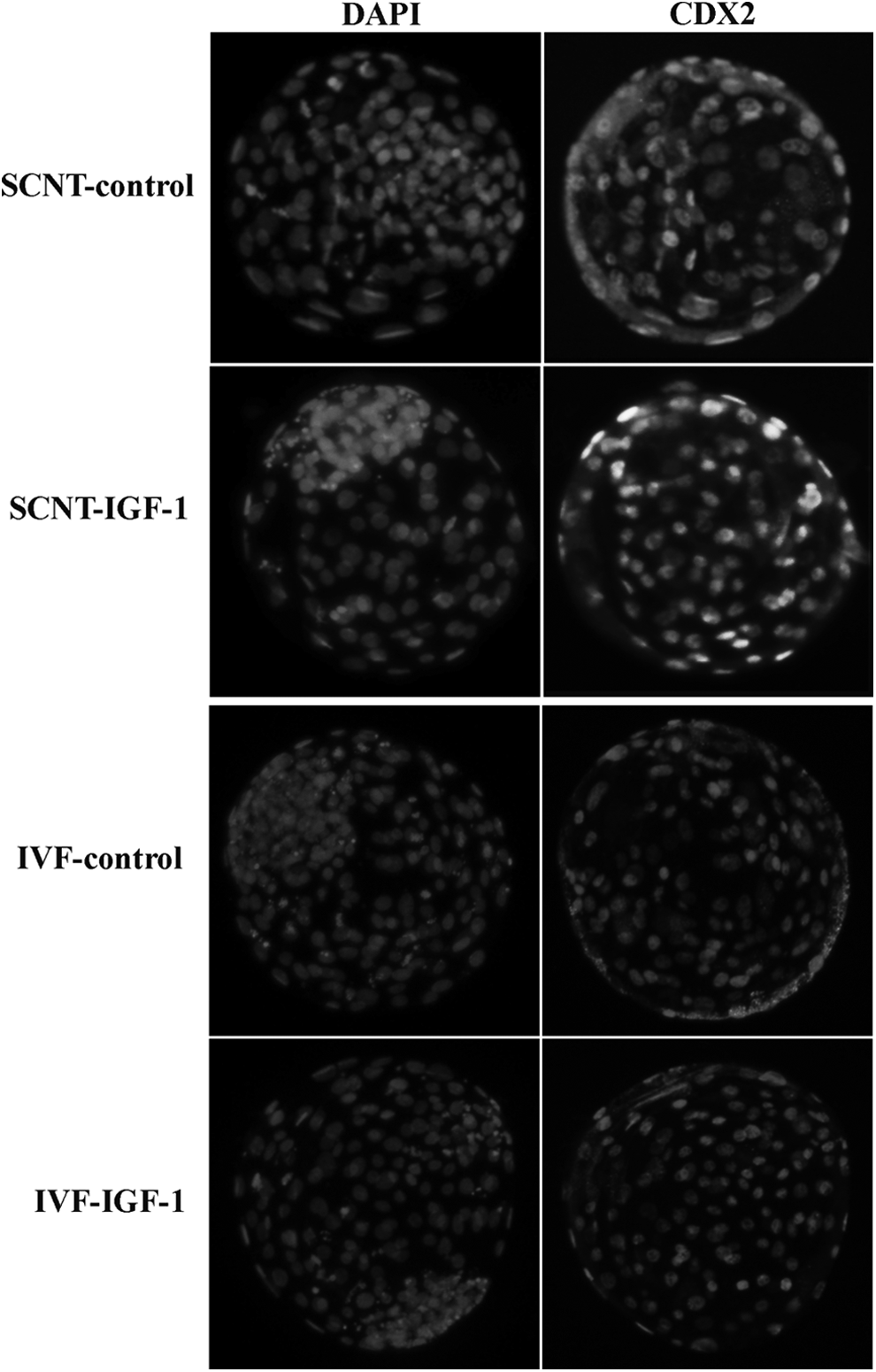
Immunostaining of TE and ICM. TE was stained with CDX2, an expression marker for TE. Nuclei were stained with DAPI. Original magnification, 200×.
For SCNT, the blastocyst rate is significantly higher in the IGF-1 group compared with the control.
For SCNT, the total blastomere number is significantly higher in the IGF-1 group compared with the control.
Pregnancy rate is the number of pregnancy recipients at 90 days after embryo transfer/number of recipient cows.
Kid born rate is the number of born cows/number of recipient cows.
Day 7 blastocysts were nonsurgically transferred (one embryo per recipient) to synchronized recipient cows.
Cloning efficiency is the number of surviving calves at 60 days after birth/number of recipient cows.
IGF-1, insulin-like growth factor-1; SCNT, somatic cell nuclear transfer; IVF, in vitro fertilization.
Apoptotic rate is apoptotic blastomeres/total blastomeres.
Within the same column, values with different superscripts are different (p < 0.05).
IGF-1, insulin-like growth factor-1; SCNT, somatic cell nuclear transfer; IVF, in vitro fertilization; SEM, standard error of the mean.
Within the same column, values with different superscripts are different (p < 0.05).
ICF-1, insulin-like growth factor-1; ICM, inner cell mass; TE, trophectoderm; SCNT, somatic cell nuclear transfer; IVF, in vitro fertilization; SEM, standard error of the mean.
Discussion
IGF-1 is an important gene in regulating embryo development, cell growth, and differentiation, and the expression patterns of IGF-1 and its family genes have been investigated thoroughly in IVF embryos. However, there was little information about the expression patterns of these genes in bovine SCNT embryos. The present study was conducted to investigate the expression patterns of IGF-1 and IGF-1R in bovine IVF and SCNT embryos. The results showed that IGF-1 is not expressed from the MII oocyte stage to the blastocyst stage in both IVF and SCNT embryos. However, IGF-1R is expressed throughout the preimplantation stages in both IVF and SCNT embryos. These results indicated that the development of preimplantation embryos might require exogenous IGF-1 from oviduct fluid. Accordingly, the effects of exogenous IGF-1 on the in vitro development of bovine IVF and SCNT embryos were assessed in the present study. The results showed that exogenous IGF-1 has no obvious positive impact on the developmental competence of IVF embryos, but can significantly improve the development competence of SCNT embryos.
In the present study, the IGF-1R could be detected in MII oocytes and all the developmental stages of IVF preimplantation embryos. This result was in agreement with earlier studies, which reported that mRNA expression of IGF-1R occurred throughout preimplantation development in IVF embryos (Bertolini et al., 2002; Lazzari et al., 2002; Wang et al., 2009; Watson et al., 1992; Yaseen et al., 2001; Yoshida et al., 1998). The expression levels of IGF-1R are changed dynamically in different developmental stages; they decrease significantly from MII oocytes to four-cell embryos and increase after embryonic genome activation (eight-cell stage). The results were coincident with those reported by Yaseen et al. (2001). This dynamic change could be explained by the fact that the maternally originated mRNAs of IGF-1R are utilized or degraded prior to the activation of the embryonic genome, and the transcripts of IGF-1R might switch on at the eight-cell stage. In addition, the present study found that the expression level of IGF-1R was similar at each developmental stage between SCNT and IVF embryos.
Consistent with previous studies, the expression of IGF-1R can be detected, yet the expression of IGF-1 can not be detected both in IVF and SCNT embryos throughout the preimplantation developmental stages (Moore et al., 2007; Wang et al., 2009; Watson et al., 1999; Winger et al., 1997; Warzych et al., 2007; Yaseen et al., 2001). This phenomenon indicated that: (1) The development of preimplantation embryos might require IGF-1. (2) Preimplantation embryos did not synthesize IGF-1 by themselves; they might absorb exogenous IGF-1 from oviduct and uterus fluid. Thus, supplementing culture medium with IGF-1 might be important for the development of in vitro–produced embryos.
Accumulated evidence has indicated that the effects of IGF-1 on the development of embryos is dependent on the embryonic status and (or) culture system (Block and Hansen, 2007). In the present study, no obviously positive effect on the development of IVF embryos was observed. This result might indicate that the culture condition and the embryonic status were good in the present study. However, positive effects of IGF-1 on the development of SCNT embryo were observed. The positive effects, including higher blastocyst formation rate, lower apoptosis index, and appropriate ICM and TE ratios, were consistent with previous findings in IVF embryos (Block et al., 2008; Makarevich and Markkula, 2002; Sirisathien and Brackett, 2003). These results indicate that SCNT embryos are in a poor developmental state when compared to IVF embryos, and thus might be more sensitive to IGF-1.
Conclusions
In summary, the present study showed that: (1) IGF-1R is expressed in both IVF and SCNT embryos throughout the preimplantation developmental stages, whereas IGF-1 is not expressed and (2) supplementing exogenous IGF-1 in the culture medium can improve the developmental competence of SCNT embryos.
Footnotes
Acknowledgments
The authors thank Wang You Nan for providing the Holstein cow ovaries used in this study. This work was carried out with support from grants of the Fundamental Research Funds for the Central Universities of China (grant no. QN2013030).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.