
Abstract
Abstract
The functions of POU class 5 transcription factor 1 (Oct-4) and caudal-type homeobox 2 (Cdx2) in the differentiation of the murine inner cell mass (ICM) and trophectoderm (TE) have been described in detail. However, little is known about the roles of OCT-4 and CDX2 in preimplantation bovine embryos. To elucidate their functions during early development in bovine embryos, we performed OCT-4 and CDX2 downregulation using RNA interference. We injected OCT-4- or CDX2-specific short interfering RNAs (siRNAs) into bovine zygotes. The rate of blastocyst development of OCT-4-downregulated embryos was lower compared with uninjected or control siRNA-injected embryos. Gene expression analysis revealed decreased CDX2 and fibroblast growth factor 4 expression in OCT-4-downregulated embryos. CDX2-downregulated embryos developed to the blastocyst stage; however, in most cases, blastocoel formation was delayed. Gene expression analysis revealed decreased GATA3 expression and elevated NANOG expression in CDX2-downregulated embryos. In conclusion, OCT-4 and CDX2 are essential for early development and gene expression involved in differentiation of ICM and TE lineages in bovine embryos.
Introduction
T
Previously, we demonstrated differences in the expression levels of several genes, including OCT-4 and CDX2, between cells of ICM and TE lineages in bovine and porcine embryos (Fujii et al., 2010, 2013). The levels of OCT-4 mRNA in ICM of bovine and porcine blastocysts were higher than those in TE. In contrast, the levels of CDX2 expression in bovine and porcine TE lineages were higher than those in ICM lineages. We concluded that OCT-4 and CDX2 may control the differentiation of ICM and TE in bovine and porcine embryos. However, OCT-4 expression was detected in both ICM and TE in cattle and pigs, even at the expanded blastocyst stage (Fujii et al., 2010, 2013; Hall et al., 2009; Kirchhof et al., 2000; Kuijk et al., 2008; Sakurai et al., 2013; van Eijk et al., 1999). Furthermore, porcine embryos in which OCT-4 was knocked down developed to the morula stage, but failed to form the blastocoel (Sakurai et al., 2013). Although these observations suggest a distinct role for OCT-4 during early development in bovine and porcine embryos, little is known about the functions of OCT-4 and CDX2 during the embryonic development of domestic animals.
Both ICM and TE lineages are functionalized and controlled by gene regulatory networks governed by key transcriptional regulators (Rossant and Tam, 2009). In murine embryos, Nanog and fibroblast growth factor 4 (Fgf4) are known targets of Oct-4 (Ambrosetti et al., 1997; Rodda et al., 2005; Yuan et al., 1995). These genes have a region bound by Oct-4, called the Oct–Sox enhancer, and are coexpressed with Oct-4 in ICM and epiblast (Ma et al., 1992; Mitsui et al., 2003; Niswander and Martin, 1992).
Furthermore, exclusive Oct-4 and Cdx2 expression patterns in murine embryos were generated by mutual repression between Oct-4 and Cdx2 (Niwa et al., 2005). However, there are some contradictory reports about the role of CDX2 regulating OCT-4 in cattle. Schiffmacher and Keefer (2013) reported that CDX2 overexpression in bovine TE-derived cells downregulated OCT-4 expression, suggesting that CDX2 negatively regulates OCT-4 in bovine embryos. However, others have shown that OCT-4 expression was unaffected by CDX2 downregulation in bovine embryos (Berg et al., 2011; Goissis and Cibelli, 2014). For domestic animals, in contrast to mice, limited information is available concerning the molecular mechanisms underlying differentiation of ICM and TE.
In this study, we investigated the effects of OCT-4 and CDX2 downregulation on the developmental competence and gene expression related to the differentiation of ICM and TE in bovine embryos with the objective elucidating the roles of OCT-4 and CDX2 during early development in these embryos.
Materials and Methods
In vitro fertilization and embryo culture
Bovine oocytes were obtained by aspirating ovaries collected at a local slaughterhouse, as previously described (Fujii et al., 2010). Cumulus–oocyte complexes (COCs) were matured in a drop of IVMD-101 medium (Research Institute for the Functional Peptides, Yamagata, Japan; Yamashita et al., 1999) and covered with mineral oil at 39°C in a humidified atmosphere of 5% CO2 in air for 22 hours. After in vitro maturation, COCs were transferred to a drop of in vitro fertilization (IVF)-100 medium (Research Institute for the Functional Peptides). Subsequently, frozen–thawed semen was centrifuged twice at 600 g for 5 minutes in IVF-100 medium, and the spermatozoa were added to the COCs at a final concentration of 5 × 106 sperm per milliliter. COCs and spermatozoa were incubated at 39°C in a humidified atmosphere of 5% CO2 in air for 6 hours.
After insemination, cumulus cells and excess spermatozoa were removed from presumptive zygotes by pipetting. Following microinjection of short interfering RNAs (siRNAs), the embryos were cultured in modified TALP (mTALP) medium (Bavister et al., 1983) with 1 mg/mL bovine serum albumin (BSA) at 39°C in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2. On Day 2 (IVF = Day 0), the embryos were transferred to mTALP with 3% (v/v) newborn calf serum (Life Technologies, Carlsbad, CA) and cultured at 39°C in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2 until Day 7 or 8. Embryonic development rates were assessed on Day 2 (2-cell≤), Day 4 (8-cell≤), Day 5 (morula and blastocyst≤), Day 6 (morula and blastocyst≤), Day 7 (blastocyst≤), and Day 8 (blastocyst≤ and expanded blastocyst≤).
Design of siRNAs and microinjection into embryos
The target sites for OCT-4 and CDX2 were selected from bovine sequences (GenBank Accession Numbers: OCT-4, AF022987.1; CDX2, NM_001206299.1). Specific siRNAs were designed using the online software BLOCK-iT™ RNAi Designer (Life Technologies; http://rnaidesigner.lifetechnologies.com/rnaiexpress/). Both sense and antisense RNA sequences for siRNAs were commercially synthesized (Table 1). After insemination, the denuded embryos were transferred to a drop of mTALP medium with 1 mg/mL BSA for microinjection. Approximately 10 pl of 50 μM specific siRNA duplexes was injected into the cytoplasm of each embryo using a Transjector 5246 (Eppendorf, Hamburg, Germany). Similarly, ∼10 pl of 20 μM nonsilencing siRNA (AllStars Negative Control siRNA; Qiagen, Hilden, Germany) was injected as a control. The embryos were washed after microinjection and cultured as described.
AS, antisense strand; F, forward; R, reverse; S, sense strand.
Determination of the relative abundance of gene transcripts
The basic protocol used to determine the relative abundance of gene transcripts was as previously described (Fujii et al., 2010; Sawai et al., 2012). On Day 5, morula-stage embryos were treated with 0.1% (w/v) protease in phosphate-buffered saline (PBS) containing 1% (w/v) polyvinylpyrrolidone for 5 minutes to remove the zona pellucida. Pools of five morulae or a blastocyst were added to 5 μL of lysis buffer (0.8%, v/v, Igepal [MP Biomedicals, LLC, Santa Ana, CA], 5 mM dithiothreitol [Life Technologies], and 1 U/μL RNasin [Promega Corp., Madison, WI]), snap-frozen in liquid nitrogen, and stored at −80°C. RNA samples were heated to 80°C for 5 minutes and then subjected to reverse transcription using the QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer's instructions. The reaction mixture was then diluted with diethylpyrocarbonate-treated water to obtain a final volume of 31 μL. Real-time quantitative PCR was performed using StepOne™ (Life Technologies), and the products were detected using SYBR Green included in the QuantiTect SYBR Green PCR Master Mix Kit (Qiagen).
The amplification program was as follows: preincubation at 95°C for 15 minutes to activate the HotStarTaq DNA Polymerase (Qiagen), followed by 45 cycles of denaturation at 94°C for 15 seconds, annealing of the primers at a range of temperatures (Table 1) for 30 seconds, and elongation at 72°C for 30 seconds. After completion of the last cycle, a melting curve was generated by starting fluorescence acquisition at 60°C and recording measurements in 0.3°C increments up to 95°C. A final quantification was performed using StepOne quantification software (Life Technologies). Expression of the target gene in each run was normalized to the internal standard Histone H2A.
Immunofluorescence staining
On Day 5, morula-stage embryos were fixed in 4% (w/v) paraformaldehyde in PBS (Wako Pure Chemical Industries, Ltd., Osaka, Japan) for 20 minutes at room temperature, and then washed twice for 10 minutes each in PBS containing 0.1% (v/v) Triton X-100 (TXPBS). Samples were subsequently permeabilized in 0.2% (v/v) Triton X-100 in PBS for 30 minutes, and then incubated in Image-iT™ FX Signal Enhancer (Life Technologies) for 30 minutes, followed by two 10-minute washes in TXPBS. Blocking was performed by incubation for 1.5 hours in 7% (v/v) goat serum (Life Technologies) in TXPBS for OCT-4 staining or in 0.5% (w/v) BSA and 1% (w/v) skim milk in TXPBS for CDX2 staining, followed by a 5-minute wash in TXPBS. Embryos were incubated with an anti-OCT-4 primary antibody (1:50; SC-9081; Santa Cruz Biotechnology, Inc., Dallas, TX) in PBS supplemented with 0.5% (w/v) BSA and 0.05% (v/v) Triton X-100 at 4°C overnight or an anti-CDX2 primary antibody (1:300; MU392A-UC; BioGenex, Fremont, CA) in PBS containing 0.5% (w/v) BSA and 0.05% (v/v) Triton X-100 at 37°C overnight (Nagatomo et al., 2013).
Next, the embryos were washed four times for 15 minutes in TXPBS. The embryos were then incubated with Alexa Fluor 488-conjugated goat anti-rabbit secondary antibody (A11034; Life Technologies) for OCT-4 staining or with Alexa Fluor 488-conjugated goat anti-mouse secondary antibody (A11029; Life Technologies) for CDX2 staining at a dilution of 1:400 at room temperature for 1 hour in PBS containing 0.5% (w/v) BSA and 0.05% (v/v) Triton X-100. The embryos were washed four times in TXPBS for 20 minutes and then mounted on to slides in a drop of VECTASHIELD Mounting Medium with DAPI (Vector Laboratories, Inc., Burlingame, CA). Fluorescent images were obtained using an inverted fluorescence microscope and a digital camera system (ECLIPSE Ti-U and DS-Fi2-L3; Nikon Corp., Tokyo, Japan).
Assessment of cell numbers
On Day 5, morula-stage embryos obtained by injection of OCT-4 or CDX2 siRNAs or a control siRNA were used for the assessment of the total cell numbers. Some morula-stage embryos were cultured up to Day 7; then, the total cell numbers of all embryos were counted. Embryos were fixed in 4% (w/v) paraformaldehyde in PBS for 20 minutes at room temperature. Samples were mounted on to slides in a drop of VECTASHIELD mounting medium with DAPI. Fluorescent images were obtained as described in the section above.
Statistical analysis
The percentage data for embryo development were subjected to an arcsine transformation. The transformed values were analyzed by one-way analysis of variance (ANOVA), followed by multiple pair-wise comparisons using the Tukey–Kramer method. With regard to OCT-4 downregulation, the levels of expression of OCT-4, CDX2, and FGF4 mRNA were analyzed using the Kruskal–Wallis test, followed by multiple pair-wise comparisons using Scheffé's method. The level of expression of NANOG mRNA was analyzed by one-way ANOVA, followed by multiple pair-wise comparisons using the Tukey–Kramer method. With respect to CDX2 downregulation, the levels of expression of CDX2, OCT-4, GATA3, and NANOG mRNA were analyzed by one-way ANOVA, followed by multiple pair-wise comparisons using the Tukey–Kramer method. The total cell numbers were analyzed using the F-test, followed by Student's t-test. p Values of <0.05 denoted a statistically significant difference.
Results
Effect of siRNA injection on OCT-4 expression
To determine the efficacy of OCT-4 downregulation by RNA interference, we evaluated the expression levels of OCT-4 mRNA and protein in uninjected, control siRNA-injected, and OCT-4-specific siRNA-injected morula-stage embryos on Day 5 (Fig. 1). The relative abundance of OCT-4 mRNA in the embryos injected with OCT-4 siRNA was significantly lower (p < 0.01) than that in the uninjected and control siRNA-injected embryos (Fig. 1A). The expression of OCT-4 protein was evaluated by immunofluorescence staining. In OCT-4 siRNA-injected embryos, it was difficult to detect a nuclear OCT-4 signal at the morula stage (Fig. 1B).

Validation of OCT-4 siRNA.
Effect of OCT-4 downregulation on the development of bovine embryos
We evaluated the in vitro developmental competence of OCT-4 siRNA-injected embryos (Table 2). No significant difference in the rate of development to the morula stage (Day 5) was observed between the experimental groups. On Day 6, the rate of development to the morula stage in the OCT-4 siRNA-injected embryos (18.2%) was significantly higher (p < 0.05) than that in the uninjected (6.6%) and control siRNA-injected embryos (7.8%). The rate of blastocyst development (Days 6 and 7) was significantly lower in the OCT-4 siRNA-injected embryos (12.3% and 16.3%, respectively) than in the uninjected (36.5%, p < 0.05, and 40.6%, p < 0.01, respectively) and control siRNA-injected embryos (31.2%, p < 0.05, and 36.1%, p < 0.01, respectively).
Experiments were replicated six times.
Percentages of the number of embryos cultured.
Values with different superscripts within each column differ significantly (p < 0.01 or 0.05).
In the OCT-4 siRNA-injected embryos, most embryos that developed to the morula stage on Day 5 maintained a morula-like morphology until Day 7 or degenerated without forming blastocoel (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/cell). In contrast, on Day 7, almost all blastocysts obtained from uninjected or control siRNA-injected embryos had expanded; some embryos had even reached the hatched stage (Supplementary Fig. S1).
As shown in Figure 2, no significant difference in the total cell numbers at the morula stage (Day 5) was observed between the control siRNA-injected (44.6 ± 4.0) and OCT-4 siRNA-injected embryos (36.0 ± 2.8). On Day 7, the total cell number of the OCT-4 siRNA-injected embryos (68.2 ± 10.0) was significantly lower (p < 0.01) than that of the control siRNA-injected embryos (134.2 ± 12.4; Fig. 2).
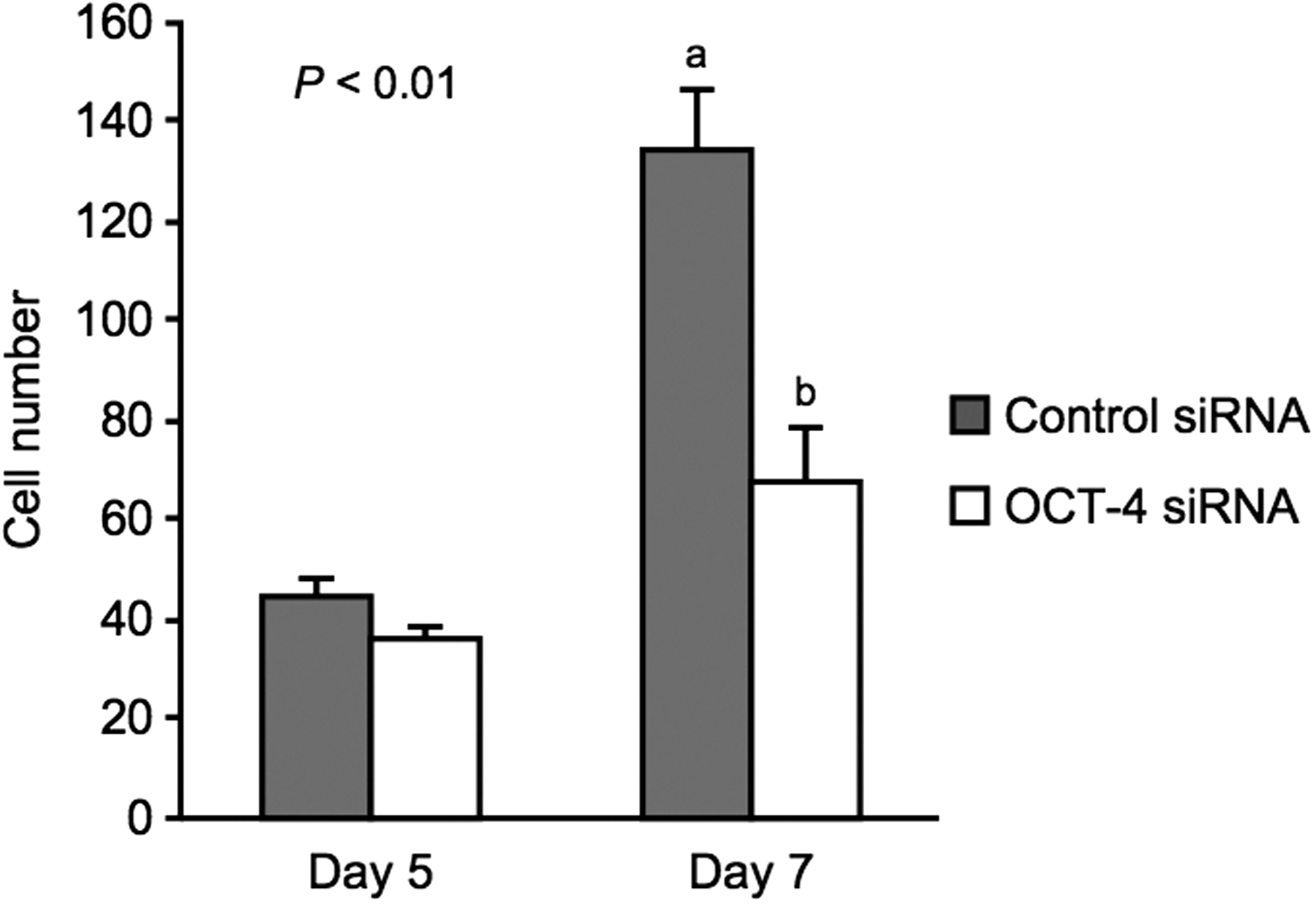
Effect of OCT-4 depletion on total cell numbers at the morula stage on Days 5 and 7. Total cell numbers (mean ± SEM) of morula-stage embryos following control siRNA (gray, n = 22) or OCT-4 siRNA injection (white, n = 20) on Day 5 were assessed. On Day 7, total cell numbers of embryos, evaluated as morula on Day 5, following control siRNA (n = 21) or OCT-4 siRNA injection (n = 18) were assessed. a,bDifferent characters indicate a significant difference (p < 0.01).
Quantitative PCR analysis of gene expression in OCT-4-downregulated embryos
To clarify the effect of OCT-4 downregulation on gene expression, we evaluated the levels of CDX2, FGF4, and NANOG mRNA at the morula stage. As shown in Figure 3A, the level of CDX2 mRNA in the OCT-4 siRNA-injected embryos was significantly lower (p < 0.05) than that in the uninjected and control siRNA-injected embryos. The embryos injected with OCT-4 siRNA showed a significant reduction (p < 0.01) in the expression of FGF4 (Fig. 3B). The relative abundance of NANOG mRNA in the OCT-4 siRNA-injected embryos tended to be lower (p = 0.09) than that in the uninjected and control siRNA-injected embryos (Fig. 3C).
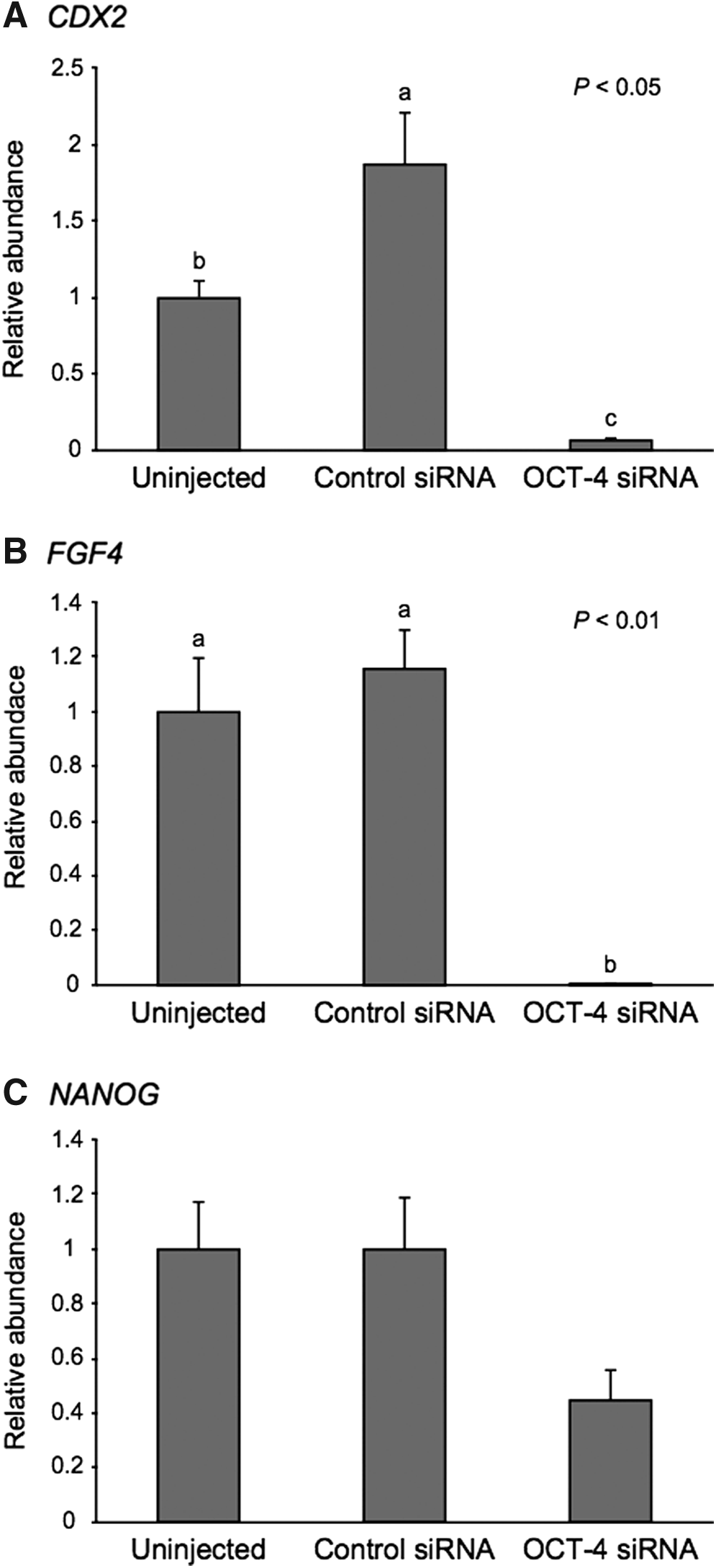
Relative abundance (mean ± SEM) of
Effect of siRNA injection on CDX2 expression
We evaluated the expression levels of CDX2 mRNA and protein in uninjected, control siRNA-injected, and CDX2-specific siRNA-injected embryos (Fig. 4). At the morula stage on Day 5, the relative abundance of CDX2 mRNA in the CDX2 siRNA-injected embryos was significantly lower (p < 0.01) than that in the uninjected and control siRNA-injected embryos (Fig. 4A). Subsequently, we evaluated the expression of CDX2 protein by immunofluorescence staining at the morula stage on Day 5, but we were unable to detect any CDX2 signal in the nuclei at that stage (data not shown), as previously reported (Goissis and Cibelli, 2014). Therefore, we evaluated the expression of CDX2 protein at the blastocyst stage on Day 6. In the CDX2 siRNA-injected embryos, it was difficult to detect a nuclear CDX2 signal (Fig. 4B); this difficulty remained on Day 7 in these embryos (Fig. 4C).
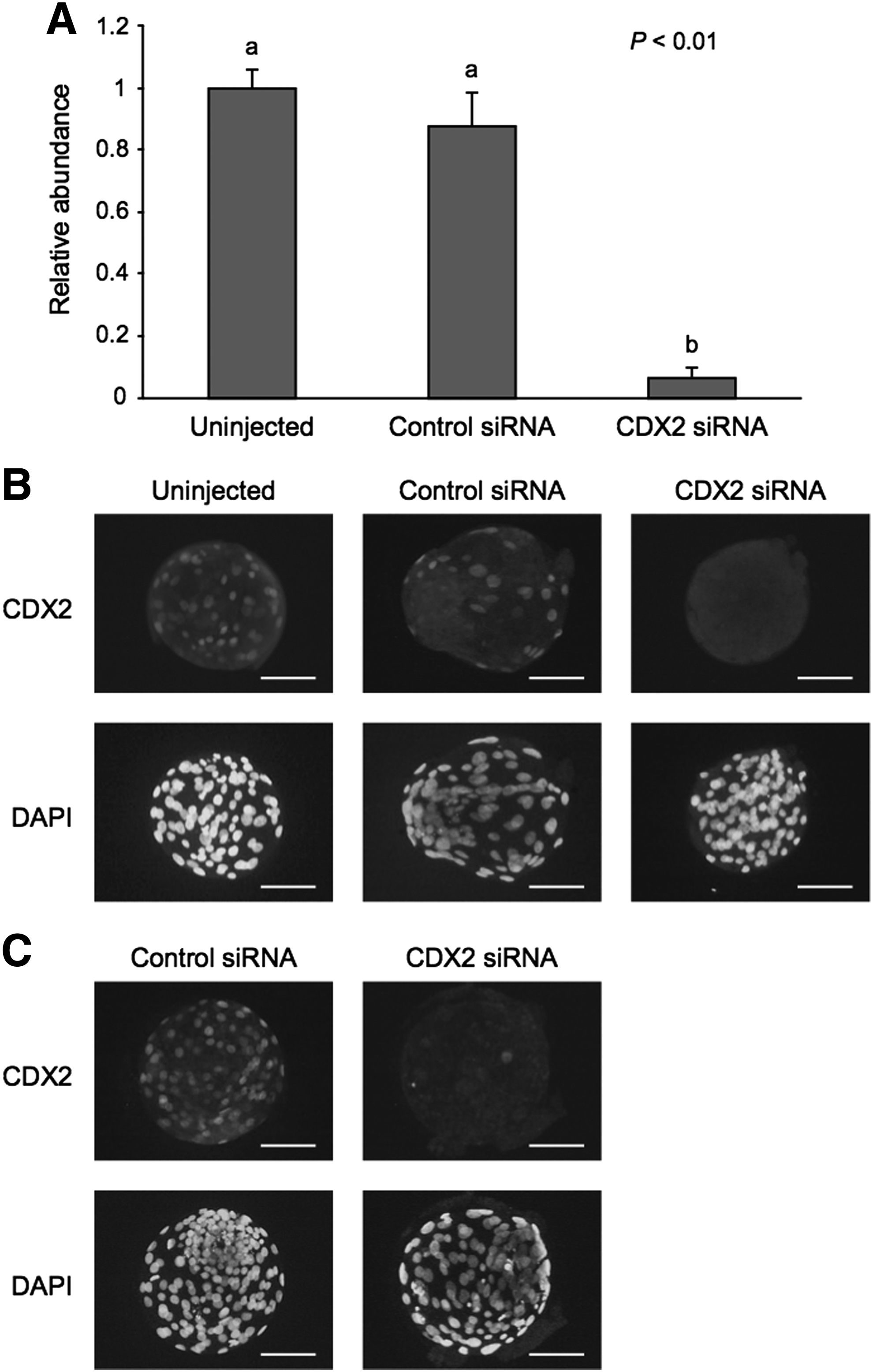
Validation of CDX2 siRNA.
Effect of CDX2 downregulation on the development of bovine embryos
Next, we evaluated the in vitro developmental competence of CDX2 siRNA-injected embryos (Table 3). No significant difference in the rate of development to the morula stage on Day 5 was observed between the experimental groups. On Day 6, the rate of blastocyst development in the CDX2 siRNA-injected embryos (13.1%) was significantly lower (p < 0.05) than that in the uninjected (36.6%) and control siRNA-injected embryos (34.4%). On Day 7, the rate of blastocyst development in the CDX2 siRNA-injected embryos remained low. On Day 8, there was no significant difference in the rate of blastocyst development between the three groups. However, the rate of expanded blastocyst development in the CDX2 siRNA-injected embryos (20.0%) was significantly lower (p < 0.05) than that in the control siRNA-injected embryos (33.9%; Table 3 and Supplementary Fig. S2).
Experiments were replicated five times.
Percentages of the number of embryos cultured.
Values with different superscripts within each column differ significantly (p < 0.05).
We assessed the effect of CDX2 downregulation on the total cell numbers in bovine embryos (Fig. 5). On Day 5, there was no significant difference in the total cell numbers at the morula stage between the control siRNA-injected (43.0 ± 3.5) and CDX2 siRNA-injected embryos (36.2 ± 2.3). On Day 7, the total cell number of the CDX2 siRNA-injected embryos (77.8 ± 9.1) was significantly lower (p < 0.01) compared with the control siRNA-injected embryos (125.1 ± 9.7).
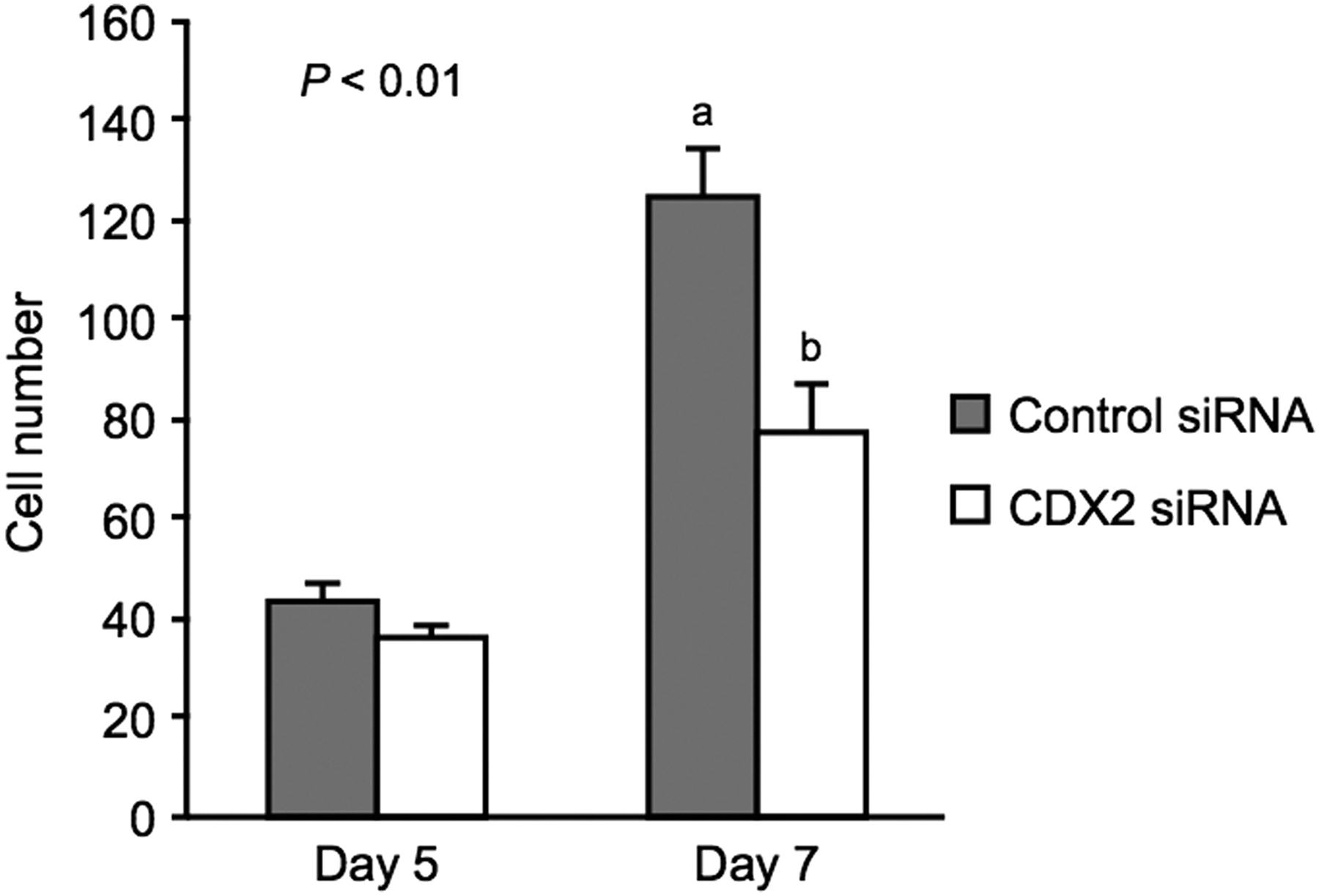
Effect of CDX2 depletion on total cell numbers at the morula stage on Days 5 and 7. Total cell numbers (mean ± SEM) of morula-stage embryos following control siRNA (gray, n = 20) or CDX2 siRNA injection (white, n = 20) on Day 5 were assessed. On Day 7, total cell numbers of embryos, evaluated as morula on Day 5, following control siRNA (n = 19) or CDX2 siRNA injection (n = 19) were assessed. a,bDifferent characters indicate a significant difference (p < 0.01).
Quantitative PCR analysis of gene expression in CDX2-downregulated embryos
To clarify the effect of CDX2 downregulation on gene expression, we evaluated the levels of OCT-4, GATA3, and NANOG mRNA at the morula stage. As shown in Figure 6A, the relative abundance of OCT-4 mRNA in the CDX2 siRNA-injected embryos was significantly lower (p < 0.05) than that in the uninjected embryos. The embryos injected with CDX2 siRNA showed a lower level (p < 0.05) of GATA3 mRNA than that in the uninjected and control siRNA-injected embryos (Fig. 6B). In contrast, the level of NANOG mRNA in the CDX2 siRNA-injected embryos was significantly higher (p < 0.05) than that in the uninjected and control siRNA-injected embryos (Fig. 6C).
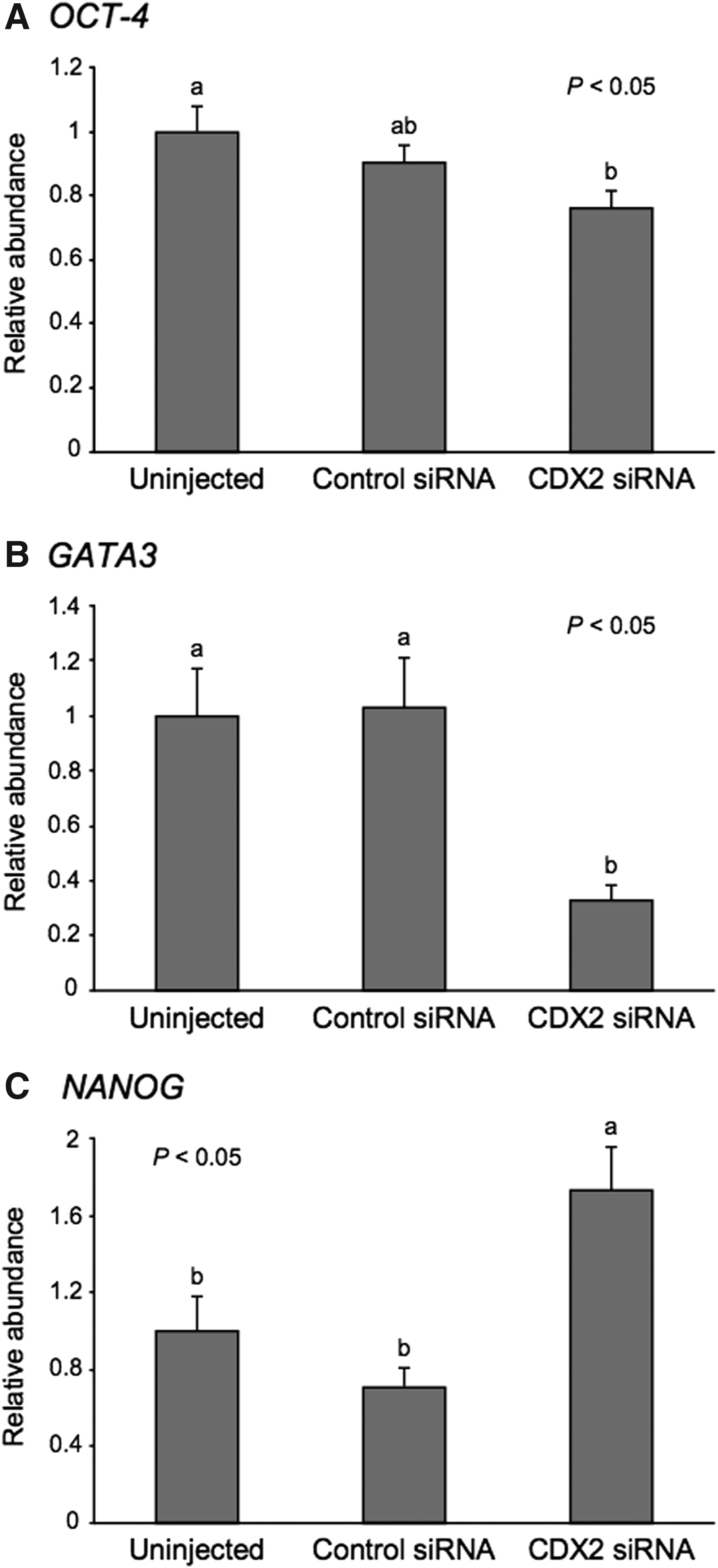
Relative abundance (mean ± SEM) of
Discussion
Several studies have shown that Oct-4 is essential for maintaining the pluripotency of embryonic stem cells and functions of ICM in mice. Oct-4 is known to regulate differentiation of ICM and direct embryonic stem cells to adopt the correct cell fate (Nichols et al., 1998; Niwa et al., 2000, 2002). In addition, Oct-4 is crucial for the induction of pluripotency in differentiated cells (Takahashi and Yamanaka, 2006). Conversely, Cdx2 promotes differentiation of the TE lineage by upregulating various TE-specific genes and downregulating Oct-4 and Nanog (Strumpf et al., 2005). Therefore, Oct-4 and Cdx2 are central regulators of differentiation of ICM and TE in murine embryos (Nichols et al., 1998; Niwa et al., 2005; Strumpf et al., 2005). In this study, we evaluated the roles of OCT-4 and CDX2 in preimplantation bovine embryos using RNA interference, and our findings suggest that these factors are critical for the normal differentiation of ICM and TE and sound blastocyst formation in cattle.
In this study, OCT-4 downregulation in bovine embryos had no effect on development to the morula stage, but blastocyst formation was perturbed. Moreover, cell proliferation was reduced during the morula-to-blastocyst stage in OCT-4-downregulated embryos. Although some OCT-4 siRNA-injected embryos developed to the blastocyst stage, normal levels of OCT-4 mRNA expression were detected in these blastocysts (data not shown). It is possible that these embryos were able to develop to the blastocyst stage because of a disappearing downregulating efficacy or incorrect injection of the OCT-4 siRNA, but we were unable to find a blastocyst with low OCT-4 expression.
As mentioned earlier, the effect of OCT-4 downregulation on development of bovine embryos was more severe compared with Oct-4 downregulation in murine embryos. Unlike murine embryos, OCT-4 expression in the bovine TE does not disappear, even at the fully expanded blastocyst stage (Kirchhof et al., 2000; van Eijk et al., 1999). OCT-4 may play a distinct role at the blastocyst stage in bovine embryos, for example, by maintaining proliferative TE cells in a differentiation-delayed state (Degrelle et al., 2005; Kurosaka et al., 2004). However, Nganvongpanit et al. (2006) reported that there was no significant difference in the rate of development to the blastocyst stage between OCT-4 double-stranded RNA (dsRNA)-injected and uninjected bovine embryos.
Although the reason why the developmental competence of OCT-4 dsRNA-injected embryos differed from that of siRNA-injected embryos is unclear, OCT-4 dsRNA-injected embryos required a longer culture period to form the blastocoel, and the number of ICM cells was significantly lower in these embryos (Nganvongpanit et al., 2006). We obtained a similar result following OCT-4 downregulation in porcine embryos (Sakurai et al., 2013). Porcine embryos express OCT-4 in TE and become elongated after hatching from the zona pellucida similarly to bovine embryos.
Although the reason underlying the arrest of blastocyst formation following OCT-4 downregulation is unclear, OCT-4 downregulation may affect the development of ICM as well as TE (Degrelle et al., 2005; Kurosaka et al., 2004). Indeed, we found that the expression of CDX2, which is a known TE-specific marker in murine embryos, was decreased in bovine embryos following OCT-4 downregulation. These results suggest that OCT-4 has common and/or distinct roles between species during blastocyst formation.
In murine embryos, Fgf4 is highly expressed in ICM and epiblast and activates the expression of fibroblast growth factor receptor 2 in the TE lineage (Allen et al., 2001; Schlessinger et al., 2000). It is established that the Fgf4 signaling pathway is required to maintain the proliferation of TE cells (Niswander and Martin, 1992; Rappolee et al., 1994; Tanaka et al., 1998). Conversely, Nanog, which is expressed in ICM, is essential for the maintenance of pluripotency in ICM and embryonic stem cells (Mitsui et al., 2003). In murine embryonic stem cells and embryonal carcinoma cells, the expression of Fgf4 and Nanog is regulated by binding of Oct-4 to the Oct–Sox enhancer (Ambrosetti et al., 1997; Kuroda et al., 2005; Rodda et al., 2005; Yuan et al., 1995). Consistent with a previously published report (Nganvongpanit et al., 2006), the level of FGF4 expression dramatically decreased in bovine embryos following OCT-4 downregulation.
Our results indicate that OCT-4 is involved in the regulation of FGF4 expression in preimplantation bovine embryos. In contrast, Nanog expression is increased by Fgf4 or Gata6 downregulation in murine embryos (Bessonnard et al., 2014; Kang et al., 2013; Yamanaka et al., 2010). In this study, NANOG expression tended to be lower, regardless of FGF4 suppression, in OCT-4-downregulated embryos. Although the levels of Fgf4 and Gata6 expression are decreased in Oct-4-deficient murine embryos, Nanog expression was hardly affected by the lack of Oct-4 (Frum et al., 2013; Le Bin et al., 2014). It is possible that multiple factors, including OCT-4, regulate NANOG expression in bovine embryos.
Reportedly, murine Cdx2-deficient embryos are able to form blastocysts, implying that Cdx2 is not essential for the initial specification of the TE lineage (Strumpf et al., 2005; Wu et al., 2010). However, Cdx2-deficient blastocysts fail to maintain blastocoel and epithelial integrity (Strumpf et al., 2005). In this study, blastocoel formation occurred in CDX2-downregulated bovine embryos. We suggest that CDX2 is dispensable for establishing TE in bovine embryos, as shown in previous reports (Berg et al., 2011; Goissis and Cibelli, 2014). Interestingly, CDX2 downregulation affected the timing of blastocoel formation and cell proliferation during the morula-to-blastocyst stage. CDX2 may function in development or differentiation at the morula-to-blastocyst stage in bovine embryos.
In murine embryos, Gata3 is exclusively expressed in TE and functions in the differentiation of TE by upregulating Cdx2 expression (Home et al., 2009; Ralston et al., 2010). Correspondingly, in bovine embryos, GATA3 was exclusively expressed in the TE lineage (Nagatomo et al., 2013). Therefore, GATA3 can be considered important for the differentiation of the TE lineage in bovine embryos. Conversely, NANOG, which is directly downregulated by Cdx2 in murine embryos (Chen et al., 2009; Strumpf et al., 2005), is undetectable in the TE lineage in bovine embryos (Kuijk et al., 2008; Nagatomo et al., 2013). The increase in NANOG expression following CDX2 downregulation suggests that CDX2 regulates NANOG expression in bovine as well as murine embryos.
In this study, GATA3 expression was reduced at the morula stage after CDX2 downregulation. Although it unclear whether CDX2 directly regulates GATA3 in bovine embryos, this decrease in GATA3 expression may have occurred because of the damaged integrity of the TE lineage caused by CDX2 downregulation. We examined the expression levels of GATA3 and NANOG mRNA at the blastocyst stage in CDX2-downregulated embryos; however, CDX2 downregulation did not affect these expression levels, although the level of CDX2 mRNA was significantly (p < 0.05) decreased (data not shown). Our results imply that CDX2 controls differentiation of TE in bovine embryos by directly or indirectly regulating various genes, such as GATA3 and NANOG, and that the role of CDX2 in promoting differentiation of TE may start from the morula stage. However, the role of CDX2 in the TE differentiation may be limited or compensated for by other genes, such as TEA domain transcription factor 4 (TEAD4), which is known to be the master regulator of the TE-specific transcriptional program in murine embryos (Nishioka et al., 2008; Yagi et al., 2007).
Oct-4 and Cdx2 mutually repress each other's expression in murine embryonic stem cells (Niwa et al., 2005). Moreover, Cdx2 knockout or knockdown causes ectopic expression and overexpression of Oct-4 in murine embryos (Ralston and Rossant, 2008; Strumpf et al., 2005; Wu et al., 2010). Therefore, we hypothesized that if reciprocal inhibition between OCT-4 and CDX2 occurs in bovine embryos, the expression of OCT-4 and CDX2 may be increased by the downregulation of their antagonist. Interestingly, OCT-4 downregulation in bovine embryos had the opposite effect; that is, CDX2 suppression was induced by OCT-4 siRNA injection.
These results imply a promotional relationship between OCT-4 and CDX2 in bovine embryos and reveal the consequence of OCT-4 expression in the bovine TE. Conversely, CDX2 downregulation did not cause OCT-4 overexpression in bovine embryos. Some investigators (Berg et al., 2011; Goissis and Cibelli, 2014) have reported that the levels of OCT-4 mRNA expression are normal in CDX2-downregulated bovine blastocysts and that the distal enhancer region of the OCT-4 gene differs between mice and cattle. Therefore, there may be no mechanism for OCT-4 repression by CDX2 in bovine embryos. Previously, we reported that OCT-4 expression in the bovine TE is decreased and disappears from the blastocyst to the elongation stage (Fujii et al., 2010), although the mechanism by which OCT-4 is repressed in the TE is unknown.
Further studies are necessary to elucidate this mechanism in bovine embryos. It remains unclear how CDX2 expression is regulated in bovine embryos. Dietrich and Hiiragi (2007) reported that the levels of Cdx2 expression initially show no correlation with the expression of Oct-4 in mice. Similarly, Ralston et al. (2010) reported that Oct-4 is not required for the initial repression of Cdx2, indicating that other mechanisms lead to the repression of Cdx2 at the morula-to-blastocyst stage. Several researchers have suggested that Tead4 induces the expression of Cdx2 in murine embryos (Nishioka et al., 2008, 2009; Yagi et al., 2007). TEAD4 expression has been reported in bovine embryos (Fujii et al., 2010); therefore, it is possible that TEAD4 regulates the expression of CDX2 in these embryos.
In conclusion, we showed that OCT-4 and CDX2 are essential for early development and gene expression involved in differentiation of ICM and TE lineages in bovine embryos. OCT-4 knockdown impaired development at the morula-to-blastocyst stage and gene expression related to the differentiation of ICM. Although CDX2-downregulated embryos were eventually able to establish TE, they exhibited reduced cell proliferation. Thus, CDX2 may function in the maintenance of TE integrity in bovine embryos. This study indicates the necessity of OCT-4 and CDX2 in preimplantation development and provides insight into the mechanisms underlying early lineage segregation in bovine embryos.
Footnotes
Acknowledgment
This work was supported by JSPS KAKENHI Grant Number 26292162.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.