
Abstract
Abstract
Cloning by somatic cell nuclear transfer (SCNT) is characterized by low efficiency and the occurrence of developmental abnormalities, which are rather poorly studied phenomena in goats. This study aimed at comparing overall SCNT efficiency in goats by using in vitro-matured (IVM) or in vivo-matured oocytes and fibroblast donor cells (mock transfected, transgenic, or wild type), also characterizing symptoms of the Abnormal Offspring Syndrome (AOS) in development, comparing results with pregnancies produced by artificial insemination (AI) and in vivo-derived (IVD) embryos. The SCNT group had lower pregnancy rate (18.3%, 11/60), total number of concepti (20.0%, 12/60), term births (3.3%, 2/60), and live births (1.7%, 1/60) than both the IVD (77.8%, 7/9; 155.5%, 14/9; 122.2%, 11/9; 88.8%, 8/9) and the AI (71.4%, 10/14; 121.4%, 17/14; 100%, 14/14; 78.5%, 11/14) groups, respectively (p < 0.05). No SCNT pregnancies reached term using IVM oocytes, but in vivo-matured oocytes resulted in two term transgenic cloned kids. The proportion fetal membrane (FM) weight/birth weight reflected an increase in FM size and cotyledonary enlargement in clones, for disproportionally bigger newborns in relation to cotyledonary numbers. Overall, goat cloning showed losses and abnormality patterns similar to the AOS in cloned cattle and sheep, which have not been previously well recognized in goats.
Introduction
C
Most problems associated with cloning procedures using somatic cells are still poorly understood (Palmieri et al., 2008; Smith et al., 2012). This scenario may be even more challenging when cloning is applied for the generation of transgenic animals, as transgenic donor cells are generally at higher cell culture passages, a factor known to cause genetic and epigenetic instability (Giraldo et al., 2006; Humpherys et al., 2001; Powell et al., 2004). Thus, the low cloning efficiency and the unpredictable occurrence of abnormalities throughout development have limited the widespread use of cloning as a tool for multiple applications (Campbell et al., 1996; Feng et al., 2015; Reggio et al., 2001; Wells et al., 1997; Yekta et al., 2013; Zou et al., 2001).
The collection of symptoms and abnormalities associated with cloning by SCNT has been named Abnormal Offpring Syndrome (AOS) by Farin et al. (2006), and it is a rather well-studied phenomenon in cattle and sheep (Chen et al., 2013; Palmieri et al., 2008; Smith et al., 2012), but not much in goats (Rutigliano et al., 2015). Faulty nuclear reprogramming involving the inefficient or insufficient modulation of epigenetic marks in the somatic nucleus is still recognized as one of the main causes of the low efficiency of cloning by SCNT and the appearance of the AOS, virtually for all animal species cloned to date (Campbell et al., 2007; Niemann et al., 2008; Smith et al., 2012). In such a context, oocyte quality and nucleus donor cell plasticity may determine the epigenetic reprogramming capacity after cloning, which has been the focus of continuous scientific investigations (Behboodi et al., 2004; Franciosi et al., 2012; Humpherys et al., 2001).
Cytoplasmic factors present in MII oocytes are expected to epigenetically reshape and remodel the donor cell nucleus, which must be temporally and quantitatively adjusted to properly reprogram the somatic nucleus (Campbell et al., 2001; Oback and Wells, 2003). On the other hand, the karyoplast type and origin, along with in vitro cell manipulation procedures, such as transgene transfection and cell selection, may influence the degree at which the somatic cell genome will endure in the epigenetic remodeling and reprogramming process after cloning (Batchelder et al., 2005; Oback and Wells, 2007; Powell et al., 2004).
Oocytes and donor cells exert leading roles in cloning success or demise, which emphasize the importance of protocols for the production of more competent cytoplasts for cloning using karyoplasts with improved epigenetic plasticity (Niemann et al., 2008; Powell et al., 2004). Thus, this study aimed at comparing the effects of the cytoplast source (in vivo- vs. in vitro-matured [IVM] oocytes) and karyoplast type from the same genetic background (wild-type control cells, transfected but diagnosed as nontransgenic cells, and transfected and detected as transgenic cells) on the overall goat SCNT cloning efficiency, comparing in vivo developmental outcome and the appearance of symptoms for the AOS in the course of development and at term with concurrent in vivo-derived (IVD) pregnancies by artificial insemination (AI) and superovulation and embryo transfer. The experimental design intended to define effective cloning protocols for use in a transgenic goat program for the production of transgenic goats expressing recombinant proteins in milk.
Materials and Methods
Ethics approval
This study was approved by the Biosafety Committee of the University of Fortaleza, biosafety certification number 0294/10.
Reagents
Most reagents were purchased from Sigma-Aldrich Chemical Company, unless stated otherwise.
Animals, and animal care and handling
This study was carried out using Nubian-crossed goats with a mean body condition score of 2.7, ranging from 2.0 to 3.5 (1–5 scale), and a mean body weight of 43.3 kg, ranging from 30 to 65 kg. Animals were kept in paddocks with free access to shaded areas for a minimum of 90 days before the onset of the experiments, receiving mineral salt and water ad libitum and a balanced diet (elephant grass, alfalfa hay, corn, and soybean to obtain 14%–16% crude protein and 65%–75% of total digestible nutrients). The study was conducted in the Brazilian Northeast region (03° 43′ 02" S and 38° 32′ 35" W), in a tropical wet and dry climate with an average annual rainfall around 1600 mm associated with long periods of drought.
Transgene expression vector
A 1.6-kb human glucocerebrosidase (hGCase) coding sequence was in vitro synthesized and cloned into the XhoI site of the pBC1 Milk Expression Vector (Life Technologies) under the goat β-casein promoter. The 1.8-kb neomycin resistance gene cassette (neoR) under the SV40 promoter was amplified from the pEGFP-N1 vector (Clontech Laboratories), subcloned into the pCR®-Blunt (Invitrogen), and inserted between the AatII-PvuI sites into the pBC-hGCase vector, replacing the ampicillin resistance gene and resulting in the pBC-NeoR-hGCase expression vector (Fig. 1).
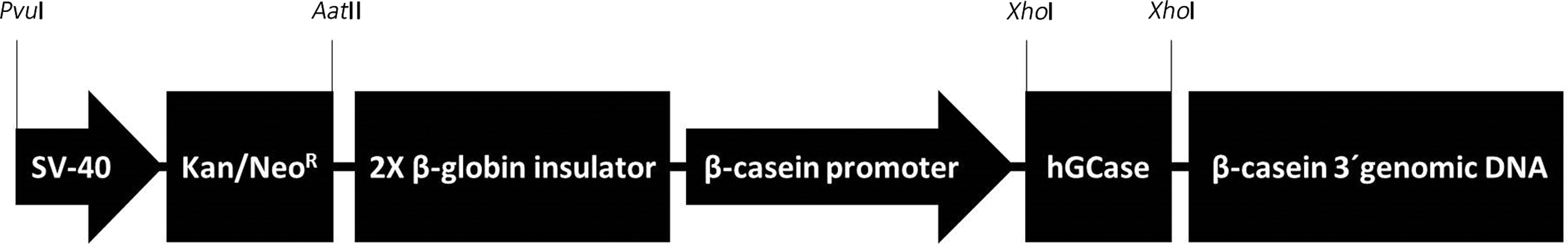
Schematic representation of the milk expression vector pBC-NeoR-hGCase, containing the 1.8-kb neomycin resistance gene cassette (Kana/NeoR) under the SV-40 promoter inserted between PvuI-AatII sites, and the 1.6-kb human glucocerebrosiase (hGCase) coding sequence (NM_001005741.2) cloned in the XhoI restriction site under the goat β-casein promoter, followed by regulatory exon regions.
Primary goat cell cultures, cell transfection, and establishment of transgenic cell lineages
Primary goat cell cultures. An ear biopsy from a 7-month-old goat female was used for the establishment of primary fibroblast cell cultures and expansion in TCM 199 medium +15% fetal bovine serum (FBS; Gibco®, Life Technologies) and 1% antibiotic-antimycotic solution. Cells at passage 2 and at medium to high confluence were frozen (Gerger et al., 2010).
Cell transfection and cell selection. Cells were transfected with the pBC-NeoR-hGCase transgene at passage 3 by electroporation (Nucleofector Amaxa®, program U12; Lonza), following the manufacturer's recommendation. Transfected fibroblast cells were seeded in 90-mm culture plates (50 × 103 per plate), in culture medium supplemented with 400 μg/mL G418 (Lonza) for 10–12 days. G418-resistant cell colonies were analyzed by polymerase chain reaction for transgene integration (data not shown).
Syngeneic cell lines used for cloning by SCNT. Two positive transgenic cell lines were used for cloning by SCNT: Cell A (CA), used at passages 10–13; and Cell B (CB), used at passage 8. In addition, two cell cultures from the same donor animal (syngeneic) were used for cloning as controls: wild-type cells (CW), which were control fibroblast cells that were neither transfected/selected nor transgenic, at passage 3 (cells that were used as the base for the other cell types in this study); and negative (or sham) control cells (CT), at passage 8, which were fibroblast cells that were transfected but resulted negative (nontransgenic) after antibiotic selection (bystander effect, Chen et al., 2002; Lai et al., 2002). Cell cycle synchronization to G0/G1 was attained after high confluence (80%–95%) followed by serum starvation (0.5% FBS) for 24 hours before SCNT (Rutigliano et al., 2015).
Source of goat oocytes for cloning procedures
For IVM oocytes, immature cumulus-oocyte complexes (COCs) were obtained either postmortem from cycling does according to Reggio et al. (2001) or in vivo by laparoscopic ovum pick up (LOPU) according to Baldassarre et al. (2003), with minor modifications. Viable COCs were IVM for 21 hours following Reggio et al. (2001) and Feltrin et al. (2014). Conversely, in vivo-matured COCs were obtained from follicle-stimulating hormone (FSH)-stimulated mature does according to Menchaca and Rubianes (2007), after a 12-day-long progesterone exposure. Donors were slaughtered for the postmortem ovary collection, for transport to the laboratory in Dulbecco's phosphate buffered saline (DPBS) at 25°C for 1 hour. Oocytes were retrieved by follicular aspiration with a 10-mL syringe coupled to a 21-G needle. In vivo-matured oocytes were immediately used for cloning procedures. The mean number of >3-mm antral follicles, and total, viable and matured goat COCs was recorded per goat female per maturation process (in vivo vs. in vitro).
Cloning by nuclear transfer procedures (SCNT group)
Goat cloning followed modified procedures by Reggio et al. (2001) and Feltrin et al. (2014). IVM and in vivo-matured COCs were denuded by pipetting in TCM-Hank's medium supplemented with 2 mg/mL hyaluronidase. Matured oocytes (MII) were selected by the presence of the first polar body in TCM-Hank's medium supplemented with 1% FBS, 1% antibiotic-antimycotic solution, and 0.2 mM sodium pyruvate (manipulation medium or MM). Oocytes were incubated for 20–30 minutes in 10 μg/mL cytochalasin B (CCB) and 5 μg/mL Hoechst 33342 in MM, in groups of eight oocytes per manipulation round. Then, oocytes were enucleated individually in 10 μL drops under mineral oil, by micromanipulation (TransferMan®; Eppendorf®), under minimal exposure to ultraviolet light. Embryo reconstruction was performed in MM +10 μg/mL CCB. At this step, 10–15 μm small and round donor cells with smooth membranes were selected to be injected into the perivitelline space of enucleated cytoplasts.
Reconstructed structures were washed in fusion medium (0.3 M mannitol, 0.1 mM MgSO4 7H2O, 0.05 mM CaCl2.2H2O, 0.5 mM Hepes, and 0.1% polyvinyl alcohol (PVA)), and they were manually aligned in a 200-μm fusion chamber coupled to an electrofusion apparatus (Multiporator®; Eppendorf) to be subjected to membrane fusion by a single 1.0-kV/cm DC pulse for 45 μsecond. Fusion rate was evaluated within the first hour, and non-fused structures were subjected to a second fusion round. The fused pairs were incubated in G-1® medium (Vitrolife) supplemented with 10 μg/mL CCB for 1.5 hours, subsequently activated in 5 μM ionomycin in G-1 medium for 5 minutes, followed by incubation in 2 mM 6-DMAP in G-1 medium for 4 hours. Then, SCNT embryos were cultured in vitro overnight in G-1 medium for about 10 hours at 5% CO2, 38.5°C and saturated humidity. Embryos were transported to the embryo transfer site in tubes with G-1 medium, using a portable incubator (Minitube®) at 38.5°C until the transfers into synchronous recipients, as described next.
AI and in vivo embryo production (IVD) procedures
Two embryo control groups with contemporaneous females from the same herd were used to evaluate the developmental potential of cloned embryos: (a) AI; (b) superovulation followed by AI, embryo collection, and transfer at the one-cell stage. The synchronization protocol for artificial insemination (AI group) used progesterone intravaginal inserts (Eazi-Breed CIDR®) on day 0, and administration of 0.75 mg D-cloprostenol intramuscular (IM), 150 IU eCG (Folligon®; MSD Saúde Animal) via IM, and removal of vaginal inserts on day 10. The cervical AI was performed 48 hours after the removal of the vaginal inserts. The in vivo embryo production (in vivo derived, or IVD group) was accomplished following Menchaca and Rubianes. (2007), with female donors subjected to two intrauterine AIs by laparoscopy, 40 and 52 hours after the removal of the vaginal inserts. Then, females were slaughtered at a local abattoir to collect one-cell stage embryos 12 hours after the last AI.
Transfer of in vivo- (IVD) or nuclear transfer- (SCNT) derived embryos
Female recipients were subjected to estrus synchronization and ovulation by the use of vaginal progesterone inserts (CIDR; Pfizer, Inc.) on day 0, followed by the administration of 0.75 mg D-cloprostenol (Prolise®; Tecnopec) and 150 IU eCG (Folligon; Intervet/Shering-Plough Animal Health), vaginal insert removal on day 10, and 50 μg gonadorelin (Gestran®; Tecnopec) on day 11, 36 hours after the insert removal. The hormonal protocol was devised for recipients to ovulate between 12 and 24 hours after fertilization or chemical activation for IVD or NT embryos, respectively. Embryo transfer procedure was based on Baldassarre et al. (2003), with minor modifications, by ventromedial surgical laparotomy between 54 and 57 hours after progesterone insert removal. After assessing the presence of ≥6 mm diameter pre-ovulatory ovarian follicle(s), the uterus was exteriorized and embryos were transferred in DPBS +5 mg/mL bovine serum albumin to the oviduct ipsilateral to the ovary bearing the largest follicle or fresh ovulation. A total of 4–8 in vivo- (IVD) and 10–18 SCNT-derived embryos were transferred per recipient.
Pregnancy diagnosis, gestational monitoring, parturition, and neonatal care
Pregnancies were diagnosed by transrectal ultrasonography on the 23rd day after embryo transfer, and they were reassessed on days 30, 37, and 44 of gestation. Then, transabdominal ultrasonographic exams were performed weekly on the SCNT group and monthly on the AI and IVD groups. When embryo death was detected in SCNT animals by lack of a heartbeat, female recipients were euthanized for the removal of the conceptus for verification of cloning and transgenesis by molecular analysis.
Pregnant females from the IVD group delivered spontaneously at term, whereas parturition in animals from the AI and SCNT groups was hormonally induced according to a protocol for goats (Chavatte-Palmer et al., 2013). Ten animals from the AI groups were subjected to pre-induction between days 140 and 144 of gestation, induction 4 days later, for the vaginal delivery to occur between days 144 and 147 of pregnancy, which served as controls for the effect of the induction protocol on birth and on parameters of neonatal physiology.
In the SCNT group, one-term delivery of twin clones was accomplished after pre-induction on day 141 (6 am), induction on day 145 (9 am), and elective C-section on day 146 (9 am). On birth, neonates were placed on sternal recumbency and dried out while aspirating excess fluids from the airways and mouth. After weighing, each neonate received doses of colostrum for a total of 8%–12% of the body weight within the first 12 hours ex utero. Neonatal care for the SCNT newborns was more intensive, following recommendations by Chavatte-Palmer et al. (2013). Specific measures included controlled room temperature (30°C), nasal oxygen supplementation during the first hours after birth, and frequent evaluation of vital parameters (body temperature, pulse, respiration rate, mucosal color, and capillary refilling time) during the immediate postnatal period. Blood glucose and total plasma protein were measured every 60 minutes for the first 3 hours, every 3 hours for the remaining of the first day, and every 6–8 hours for the following 9 days. Urinalysis was also performed whenever possible (Uridiag®; Daydiag). Blood and urine assessments of SCNT newborns were compared with those of their IVD counterparts.
Collection and analysis of fetal membranes
Fetal membranes (FM) were collected after vaginal release in the AI and SCNT groups. For two animals in the SCNT group, FM were collected after euthanasia due to fetal death detected by ultrasonography on days 109 and 130 of gestation. FM were photographed, inspected for anomalies, and assessed for morphological and morphometric parameters. Weight, length, width, and total area of the membranes were assessed along with the number, length, width, and individual and total cotyledonary surface areas (SAs). Measures of the SA of membranes and cotyledons were estimated using the equation for determining the area of an ellipsis (length/2 × width/2 × π).
Data analysis
Each dataset was tested for normality using the Kolmogorov–Smirnov test, and when needed, data were normalized by log transformation. Oocyte viability and maturation, fusion after embryonic reconstruction, pregnancy, embryonic/fetal death, and birth rates were compared between groups by Chi-square or Fisher's exact tests. The mean number of total matured and viable oocytes, birth weights (BWs), FM weights and SAs, and cotyledonary areas were compared by analysis of variance, using the general linear model (GLM) procedure, with pairwise comparisons by the Tukey test (Minitab®). A simple correlation test was used to evaluate associations between variables. The level of significance was 5%.
Results
Oocyte retrieval, maturation, and cloning by nuclear transfer
The mean numbers of >3-mm follicles and of retrieved, viable, and matured oocytes, on a per animal basis, according to the maturation process, are shown in Table 1. A total of 4997 in vivo- (n = 1130) and in vitro- (n = 3867) retrieved oocytes were obtained from 191 donor does, rendering 556 (49%) and 1419 (37%) matured (MII) oocytes, respectively, with both maturation processes and resulting in a similar mean number of matured oocytes per donor female (Table 1).
Different superscripts in the column differ, for p < 0.05.
By ANOVA; †By chi-square test.
ANOVA, analysis of variance; COC, cumulus-oocyte complexes.
The goat cloning efficiency using IVM or in vivo-matured oocytes is presented in Table 2, with no differences observed between oocyte sources for the compared parameters. A total of 1189 couplets were reconstructed, resulting in 929 (78.1%) fused reconstructed embryos. Although no differences were observed between the source of oocytes (in vivo vs. in vitro maturation) for fusion and pregnancy rates, a higher number of losses occurred when using IVM oocytes, with no pregnancies reaching term. On the other hand, a trend for a lower pregnancy outcome occurred when using IVD oocytes (p = 0.077). Nevertheless, the use of in vivo-matured oocytes resulted in the birth of two twin cloned transgenic offspring, from which one survived and the other died as a consequence of a congenital heart defect (next).
Same superscripts in the column do not differ, for p < 0.05.
By chi-square test.
The overall efficiency in the AI, IVD, or SCNT groups is depicted in Table 3 and Figure 2a, and the temporal distribution of pregnancy losses in the AI, IVD, and SCNT groups is depicted in Figure 2b, with pre- and post-natal losses and live birth obtained in the SCNT group, by cell type, presented in Figure 3. Pregnancy and birth rates in the AI and IVD groups were high and within normal and expected physiologic ranges, with low pregnancy and neonatal losses. Pregnancy rates in the SCNT group varied widely depending on the donor cell line, with a high rate of gestational losses and low birth outcome. Overall, the SCNT group had lower pregnancy rate (18.3%, 11/60), total number of concepti (20.0%, 12/60), term births (3.3%, 2/60), and live births (1.7%, 1/60) than the IVD (77.8%, 7/9; 155.5%, 14/9; 122.2%, 11/9; and 88.8%, 8/9) and the AI (71.4%, 10/14; 121.4%, 17/14; 100%, 14/14; and 78.5%, 11/14) groups, respectively, which were similar to one another. Gestational losses were exceedingly high in the SCNT group (90.9%, 10/11) in comparison to the IVD and AI groups, which did not have any loss. The perinatal death within the first 24 hours after birth was 50% (1/2) versus 37.5% (3/8) and 27.3% (3/11) in the SCNT compared with the IVD and AI groups, respectively, with both being similar to each other. The proportions of pregnancies and of live births per number of transferred embryos were approximately 10- and 100-fold lower in the SCNT group (1:78 and 1:858) than in the IVD group (1:8 and 1:7), respectively.

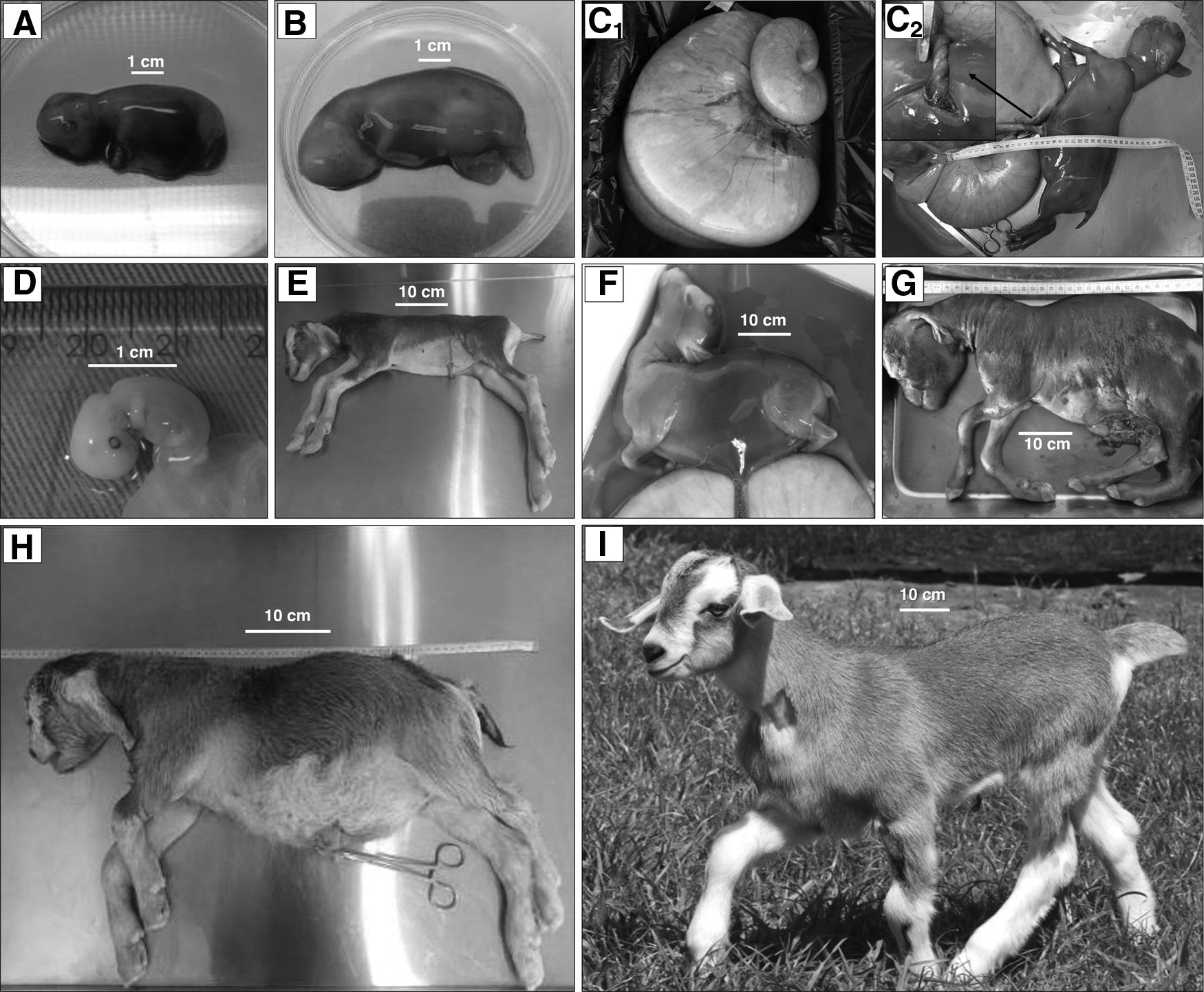
Embryonic, fetal, and post-natal losses and healthy newborn obtained in the SCNT group after reconstruction with distinctively manipulated syngeneic fibroblast donor cells.
Different superscripts in the column differ, for p < 0.05.
By chi-square or Fisher's exact test.
AI, artificial insemination; CA, Cell A; CB, Cell B; IVD, in vivo-derived; SCNT, somatic cell nuclear transfer.
Pregnancy outcome, pre-natal monitoring, conceptus abnormalities, and pregnancy losses
The timeline for embryonic, fetal, and post-natal losses until the birth of a viable cloned offspring pregnancy was evaluated in the SCNT group based on the different donor cell types, as shown in Figures 2b and 3. The SCNT group was characterized by a low overall efficiency to produce viable offspring due to high embryonic and fetal losses. Cloning using cell type CA resulted in losses observed during all gestational periods, detected on days 26, 37, and 130, with only one pregnancy reaching term that yielded offspring. However, cloning using cell type CB did not result in offspring, being usually associated with earlier losses, with 80% not reaching the last trimester of pregnancy (conceptus deaths detected on days 30, 32, 43, and 58).
One female carrying a conceptus derived from an embryo reconstructed with an IVM oocyte and a cell type CB was diagnosed with fetal hydrops at mid-pregnancy. After regular monitoring, fetal death was detected by transabdominal ultrasonography on day 109, with the animal subjected to euthanasia for the collection and gross morphological evaluation of the pregnant tract and contents. After removal, weighing, and a careful dissection of the pregnant tract (Fig. 3C1), a total of 26 L of allantoic fluid was obtained from the 30-kg whole tract, with an osmolarity of 279 mOsm/kg, and only 300 mL of amniotic fluid recovered, at 299 mOsm/kg osmolarity. The conceptus had 88 placentomes, and after removal of all uterine contents, the empty thin-walled uterus weighed 1.3 kg. The non-viable fetus weighed 1.8 kg, with generalized edema. A severe torsion of the umbilical cord was observed near the fetal abdominal wall (Fig. 3C2), which may have been the main cause of fetal death and the edema.
When using cell types CW or CT, pregnancies reached the last trimester of pregnancy, but fetal deaths were detected on days 100 and 135, respectively, with no pregnancy reaching term. Perinatal losses were high in the SCNT group, although no statistical difference was observed between groups. All retrieved concepti were confirmed as clones and/or transgenic after molecular analysis.
Neonatal care
One pregnant female reached term carrying viable twin cloned concepti produced from in vivo-matured oocytes and cell type CA. One viable cloned female was born weighing 4 kg (Fig. 3I). At birth and during the first 24 hours of life, the animal had physical and clinical parameters within normal-range values for the species and within values observed in the AI group. However, the animal developed gastrointestinal, thermoregulatory, and metabolic disturbances in the next few days. Recurrent intestinal constipation, accompanied by hyperthermia above 40°C, occurred between the 2nd and the 15th day of life. The animal was treated with enema, mineral oil PO, flunixin meglumine (Banamine®; MSD Saúde Animal) and metamizole (D500®; Fort Dodge, Brazil) IM, and alcohol baths at 25% when needed. Hyperglycemia (>150 mg/dL) and glycosuria, observed on the fourth day after birth, gradually resolved on their own after a few days. After this period of instability, the animal grew healthy through adulthood.
The other twin female weighed 7 kg (Fig. 3H) and presented fetal anasarca, cardiac insufficiency, and respiratory arrest. Immediate emergency treatment consisted of oxygen therapy, cardiac massage, and 0.1 mg/kg IV bolus administration of epinephrine (Hydren®;Hypofarma), 50 mg endotracheal surfactant (Poractant alfa, Curosurf®; Chiesi), and 3 mg/kg aminophylline (Farmace) IM. Despite efforts, the animal died within 15 minutes after birth. Post-mortem findings included hypoplasia of the spiroid colon and aplasia of two tricuspid valve leaflets, causing a right-sided heart insufficiency and leading to right-atrium dilation, pericardial effusion, ascites containing 1 L of fluid, and hepatic congestion with round edges. Umbilical cord samples collected at birth from both twins confirmed them as clones and transgenic to hGCase.
Assessment of the FMs
Comparisons of FM morphologic and morphometric data between the SCNT and AI groups are presented in Tables 4 and 5. For the AI group, BWs and FM measurements were based on the delivery of singletons (AI:1), twins (AI:2), triplets (AI:3), or quintuplets (AI:5), with data representing the range for each parity subset or their associations (AI:1 + 2, AI:1 + 2 + 3, AI:1 + 2 + 3 + 5). The group AI:1 + 2 + 3 + 5 can be also interpreted as a control with normal mean values and range expected for fetal and placental measurements for the species. Considering the body weight at birth and the physical assessments performed in the FM of each parity subset after AI, an inverse relationship was observed between parity and (a) BW, (b) length, width, total SA, and weight of individual FM, (c) number and total SA of cotyledons in each FM, and (d) ratio between number of the cotyledons and BW (n/kg). On the other hand, the length, width, and individual cotyledonary SA and the proportion FM weight:BW (g/g) tended to increase as a function of parity in the AI group.
Different upper case superscripts within the same dataset in the row differ, for p < 0.05.
AI1, AI2, AI3, AI5: term AI pregnancies with singletons, twins, triplets, or quintuplets, respectively.
AI:1 + 2: AI subgroup, including data from term pregnancies with singletons and twins.
AI:1 + 2 + 3 + 5: AI subgroup, including data from term pregnancies with singletons, twins, triplets, and quintuplets.
Denotes the number of pregnancies for each of the groups and subgroups.
Denotes the number of FM evaluated in each of the groups and subgroups.
Abortion on day 130 of gestation.
BW, birth weight; FM, fetal membranes; SA, surface area.
Denotes the number of pregnancies for each of the groups and subgroups.
Denotes the number of FM evaluated in each of the groups and subgroups.
Abortion on day 130 of gestation.
BW, birth weight.
Newborn/fetal and placental measurements were significantly greater in SCNT pregnancies when compared with values from the AI:1 + 2 + 3 + 5 control pregnancies. As SCNT pregnancies carried only singleton or twin fetuses, BW in the SCNT group did not differ, but FM lengths, SA, and weights were statistically higher in the SCNT group than in the AI:1 + 2 subgroup. Thus, the proportion between the total FM weight and the BW (g/g), which was 1:8 in the SCNT groups and 1:10 and 1:9 in the AI:1 + 2 + 3 + 5 and AI:1 + 2 subgroups, respectively, reflected an increased FM mass in the SCNT group.
The SCNT group also showed larger values for length, width, and individual and total cotyledonary SA than the AI:1 + 2 + 3 + 5 and AI:1 + 2 subgroups. The individual or total cotyledonary SA in the SCNT group was 2.0- and 2.2- or 3.4- and 2.3-fold larger than in the AI:1 + 2 + 3 + 5 and AI:1 + 2 subgroups, respectively. Although the mean number of cotyledons present in FM of clones was lower than in the AI:1 + 2 subgroup, no differences were detected in the AI:1 + 2 + 3 + 5 subgroup. The ratio between cotyledonary numbers and BW (n of cotyledons/kg at birth) was ∼50% lower in the SCNT group (10.9) than in the AI:1 + 2 + 3 + 5 (20.5) and AI:1 + 2 (24.1) subgroups, denoting that cloned newborns were disproportionally bigger in relation to the number of cotyledons, when compared with all AI control groups and subgroups (Table 5).
A compensatory effect to the elevated fetal weight was observed considering the existing proportion between BW and total cotyledonary SA (kg/cm2), which was higher in the SCNT than in the AI:1 + 2 + 3 + 5 and AI:1 + 2 groups. Likewise, the proportion between total cotyledonary SA and total FM SA (cm2/cm2) in the SCNT and the AI groups supports the observation that disproportionally larger cotyledons were present in FM of cloned neonates (Table 5). Indeed, the distribution of cotyledons based on the individual SA shows a deviation to the right in the SCNT group, with a higher frequency of larger size for a lower number of cotyledons when compared with the AI group (Fig. 4a). A positive correlation (r = 0.997, p < 0.05) between individual cotyledonary SA and total FM-SA was observed in the SCNT group, whereas no such relationship was seen in the AI:1 + 2 + 3 + 5 and AI:1 + 2 subgroups. Conversely, a negative correlation (r = −0.999, p < 0.05) was detected between cotyledonary number and total SA in SCNT-derived placentas. This correlation was slightly positive (r = 0.617) in the AI:1 + 2 + 3 + 5 subgroup but was not observed in the AI:1 + 2 subgroup.
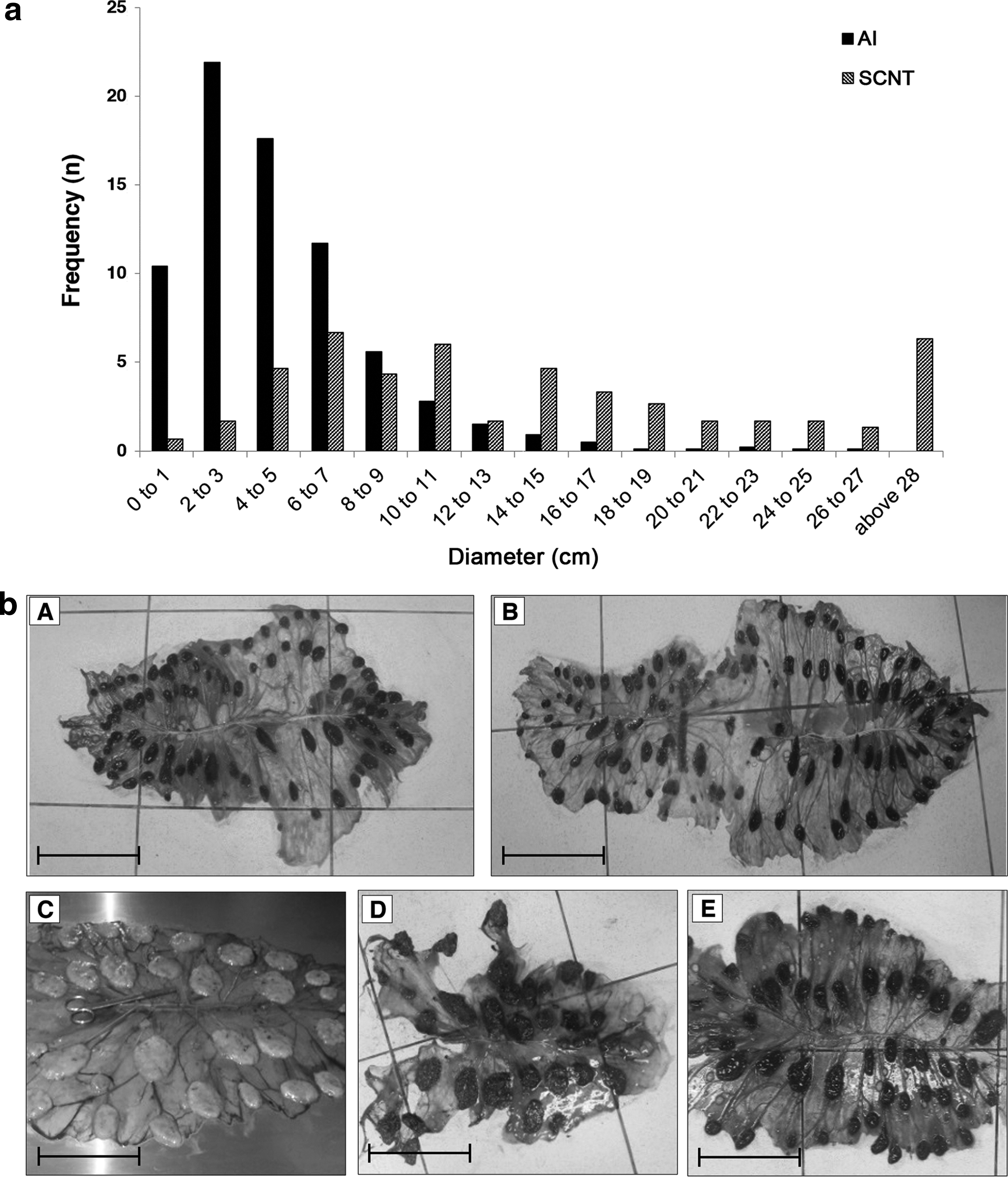
FM from two AI- and three SCNT-derived pregnancies are shown in Figure 4b. The FM and cotyledonary measures from the healthy clone closely resembled values observed in the AI group, whereas the greatest divergence from normality was seen for the conceptus that resulted in the deceased clone, which had larger values for BW, FM-SA and weight, and individual and total cotyledonary SA than the AI group and subgroups. The day-130 aborted cloned conceptus had values that fell in between the other two SCNT-derived cloned concepti.
Discussion
Even with the increasing diversity of species cloned to date, general efficiency of cloning by SCNT had not improved much since the birth of Dolly by Wilmut et al. (1997). Quality of oocytes and preparation of donor cells are positively correlated to the reprogramming competence of the recipient cytoplasm and to the plasticity of donor karyoplast, with both aspects still being at the spotlight of investigations (Franciosi et al., 2012; Niemann et al., 2008; Oback and Wells, 2007). Global cloning efficiency may be further hampered when cloning procedures are used as a tool for the production of transgenic animals, in which donor cells are genetically modified by the genome integration of exogenous DNA and in vitro cultured up to rather high passage levels (Giraldo et al., 2006; Zakhartchenko et al., 2001).
Faulty nuclear reprogramming is still considered one of the main reasons for the low SCNT efficiency (Dean et al., 2001; Lee et al., 2014; Niemann et al., 2008; Smith et al., 2012), with the appearance of pre- and post-natal abnormalities characterizing the AOS, commonly observed after in vitro embryo manipulations such as cloning by SCNT (Batchelder et al., 2007; Cibelli et al., 1998; Ricci et al., 2009; Zakhartchenko et al., 1999). In this study, the goat cloning efficiency was evaluated considering the source of recipient oocytes (cytoplast), types of donor cells (karyoplast), and cell and embryo in vitro manipulation, by accessing subsequent failures on development that could be related to the AOS in goats.
Cloning efficiency
The overall cloning efficiency was low in this study, especially mainly due to low pregnancy rates associated with high gestational losses. The control groups represented by cervical AI and oviductal transfer of IVD embryos ascertained the overall fertility status of the herd, the synchronization protocols and hormone treatments, and the rather innocuous effect of the embryo handling and surgical procedures on pregnancy and live birth outcomes. Those observations also demonstrated that the lower performance observed after cloning, characterized by low pregnancy rates, high gestational losses, and low delivery outcome of viable offspring were inherently due to the cloning procedure per se. The overall mean pregnancy rate in the SCNT group was 18.3% (11/60), which was lower than reported in a worldwide survey for goat cloning (30%–35%; Chavatte-Palmer et al., 2013) and in early studies (50%–78%; Baguisi et al., 1999; Behboodi et al., 2004; Hosseini et al., 2015a; Keefer et al., 2001). However, some studies reported similar lower rates (7%–22%) as this after using either IVM (Meng et al., 2013; Reggio et al., 2001; Yekta et al., 2013; Zhang et al., 2010) or in vivo-matured oocytes (Wan et al., 2012; Zou et al. 2001).
The efficiency of term births per transferred embryo (0.23% considering the mean values for all cloning groups, and 1.1% when analyzing the specific association between in vivo-matured oocytes and cell type CA that corresponded to the two births in this study) was in accordance with other reports (0%–1.3%; Feng et al., 2015; Lan et al., 2006; Meng et al., 2013; Ni et al., 2014; Yekta et al., 2013; Zhang et al., 2010; Zou et al., 2001), but it was still below values reported by early studies (2.5%–7.4%) using in vivo-matured (Zou et al., 2002) or IVM COCs (Baldassarre et al., 2003; Keefer et al., 2001). A rather low overall efficiency was also observed in a successful large-scale transgenic goat cloning program performed by GTC Biotherapeutics Corporation, as reported by Gavin et al. (2013), in which the efficiency was 0.89% and 0.39% after using fetal- or adult-derived fibroblast cells for cloning, respectively.
Our high rate of pregnancy losses (90.9%) was likely the major determinant of the low efficiency in this study, similar to other studies reporting losses ranging from 80% to 100% (Baguisi et al., 1999; Lan et al., 2006; Zhang et al., 2010). Despite previous reports indicating otherwise (Baldassarre et al., 2003; Keefer et al., 2001, 2002; Reggio et al., 2001; Rutigliano et al., 2015), goat cloning may result in similar excessive patterns of gestational losses (50%–80%) and other abnormalities as observed in sheep and cattle (Batchelder et al., 2007; Campbell et al., 1996; Schnieke et al., 1997; Wells et al., 1997; Wilmut et al., 1997).
Cytoplast source
The different oocyte sources resulted in different numbers of total and viable COCs, but they yielded similar mean numbers of matured oocytes (MII) between groups, regardless of the maturation process. As the oocyte competence pre-determines the nuclear and cytoplasmic maturation potential, in both in vivo and in vitro conditions (Junka et al., 2003; Sirard and Desrosier, 2007), results when using IVM oocytes were surprising, as pregnancy rates (10/42; 23.8%) were compatible with the literature (Gavin et al., 2013; Meng et al., 2013; Reggio et al., 2001; Wan et al., 2012; Yekta et al., 2013; Zhang et al., 2010; Zou et al., 2001), but these resulted in very high gestational losses (10/10). Despite the lack of success with IVM oocytes, their use in cloning has been successful, irrespective of whether COCs were collected after in vivo retrieval by LOPU (Baldassarre et al., 2003; Keefer et al., 2002) or postmortem from slaughterhouse ovaries (Feng et al., 2015; Hosseini et al., 2015a, 2015b; Nasr-Esfahani et al., 2011; Reggio et al., 2001; Rutigliano et al., 2015; Yuan et al., 2014).
Interestingly, in this study, from 18 transfers of in vivo-matured oocytes, only 1 pregnancy (5.6%) was achieved and reached term. In vivo-matured CCOs were shown as more competent in sheep (Wells et al., 1997), cattle (Behboodi et al., 2001), and goats (Behboodi et al., 2004); whereas transcriptional and epigenetic patterns have been altered in IVM oocytes (Franciosi et al., 2012; Wang et al., 2010), making in vivo-matured oocytes the recipient cytoplasts of choice for sheep (Schnieke et al., 1997; Wilmut et al., 1997) and goat (Baguisi et al., 1999; Zou et al., 2001) cloning. However, the use of the in vivo-matured goat oocytes for cloning has some limitations.
The individual oocyte variation among oocytes hinders the prediction of the best moment for oocyte retrieval, as the beginning and the progression of the in vivo maturation, determined by the endogenous plasma lutenizing hormone (LH) surge (Hyttel et al., 1997; Kruip et al., 1983) is variable, resulting in oocytes with different competence and/or potential for development after cloning. Not surprisingly, a morphologically heterogeneous population of in vivo-matured oocytes was observed, presumably at the metaphase II, late metaphase II, and telophase II stages, as also observed by others (Baguisi et al., 1999; Behboodi et al., 2001), with the in vivo maturation procedure resulting in a reduced number of highly competent oocytes for nuclear reprogramming, translated into low pregnancy rates and low birth outcome. Thus, in vivo-matured oocytes may represent a spectrum from aged oocytes to rather immature, cytoplasmically incompetent, or young MII oocytes that may be unresponsive to activation, reducing cloning efficiency.
The cytoplasm quality is crucial to cloning outcome, as all oocyte features are important for subsequent embryonic and fetal development, influencing the oocyte transcriptome and proteome (Huang et al., 2014). Chromatin remodeling and reprogramming factors (proteins, messenger RNAs [mRNAs], microRNAs, and molecular precursors) that accumulated during folliculogenesis are required for genome reprogramming and embryonic development until the activation of the embryonic genome (Alberio et al., 2001; Gao et al., 2007), and they must be both temporally and quantitatively appropriate to reprogram the somatic nucleus after cloning (Campbell et al., 2001; Oback and Wells, 2003).
Karyoplast type
Choice of the karyoplast is another main issue in animal cloning, accounting for great variations in the success rates (Campbell and Alberio, 2003; Campbell et al., 2007). By compiling the results obtained with the transgenic cells CA and CB, a pregnancy outcome halved results obtained using non-transgenic control cells CW and CT. It is worth noting that cells CA, CB, CW, and CT were derived from the same animal (syngeneic), thus eliminating the genotype variation. The processes of transfection, selection, and multiplication that are usually used in the production of transgenic cells may induce cellular differentiation, accumulation of chromosomal errors, and alteration of the epigenetic cell profile (Batchelder et al., 2005). The cellular differentiation is known to reduce the cellular ability for cloning, whereas genetic (Giraldo et al., 2006) and epigenetic (Humpherys et al., 2001) alterations are usually more elevated with longer culture periods.
Previous studies demonstrated that non-transgenic control cells have a trend to be more efficient in cloning by SCNT than the transgenic cells that are transfected and selected through selective drugs (Zakhartchenko et al., 2001). Moreover, cells collected from transgenic cloned fetuses or cloned animals, derived from nuclear transfer procedures using somatic cells that went through transfection and selection processes, may continue to propagate the lower competence when used for recloning (Zakhartchenko et al., 2001). Such outcomes may be due to the transfection and selection procedures, which entail aging associated with cell exposure to detrimental conditions, as well as to the random incorporation of exogenous DNA into the somatic genome, potentially affecting embryonic, fetal, and post-natal developments (Batchelder et al., 2005; Giraldo et al., 2006; Humpherys et al., 2001; Zakhartchenko et al., 2001). Interestingly, the worldwide survey for goat cloning by Chavatte-Palmer et al. (2013) did not demonstrate any differences in efficiency and outcome after cloning using either fetal or adult cells, and transgenic or wild-type cells.
In this study, the CT cell resulted in a good pregnancy rate (50%). Such a cell type resulted from a transfection associated to a non-effective selection process, a relatively common event called bystander effect, in which transgenic cells provide a paracrine or direct protection to the neighboring non-transgenic cells (Chen et al., 2002; Lai et al., 2002). In this case, the cell manipulation and passage level did not seem to affect pregnancy rates after cloning. The difficulties in performing comprehensive controlled studies and the low efficiency of the procedure, as seen in this study and in those by others (Baguisi et al., 1999; Beehboodi et al., 2004; Keefer et al., 2001; Ni et al., 2014; Yekta et al., 2013), challenge adequate comparisons among different donor cell types and recipient cytoplasts for SCNT.
In addition to the aforementioned cytoplast and karyoplast significance, the general SCNT procedures and embryo culture conditions are also crucial for successful cloning. In this sense, the use of G1 medium for a short in vitro culture period for cloned goat embryos, as in this study (10 hours), has already been effectively used by Rutigliano et al. (2015) for 12 hours before transfers, although other pieces of evidence by Hosseini et al. (2015b) contradict such findings. Those authors compared the use of SOF1/SOF2 and G1.2/G2.2 sequential media for embryo culture of goat cloned embryos up to the blastocyst stage, reporting that the use of G1.2/G2.2 could adversely affect pregnancy outcome (establishment of pregnancy, full-term development, and live birth). In our study and in the one by Rutigliano et al. (2015), only G1 medium was used for a short-term (10–12 hours) in vitro culture. Likely, the longer embryo culture period up to the blastocyst stage in G1.2/G2.2 medium has limited the establishment of pregnancies and births in the referred study (Hosseini et al., 2015b). Further studies should be performed evaluating the effects of culture media for a shorter or longer embryo culture period on in vivo embryo developmental outcome after cloned embryo transfer.
AOS in goats
The gestational losses of clones produced by SCNT have been associated to placental anomalies, metabolic disorders, and cardiopulmonary abnormalities (Batchelder et al., 2007; Cibelli et al., 1998; Ricci et al., 2009; Zakhartchenko et al., 1999). Placental dysfunctions, which are probably the primary cause of embryo and fetal losses of clones, are characterized by morphometric and morphologic alterations and disturbances, such as reduced vascularization, decreased binucleated cell number, placental edema, and hydrallantois (Batchelder et al., 2005, 2007; Constant et al., 2006; Ricci et al., 2009). Such findings are better characterized in cattle. Compared with other cloned ruminant species, cloned goats are considered less prone to abnormalities that are usually related to the AOS (Keefer et al., 2001, 2002; Reggio et al., 2001; Rutigliano et al., 2015). Still, limited information is available in the literature with systematic comparisons between control and cloned goats. Our findings in goats agree with those reported for other species in frequency and spectrum of abnormalities (Chavatte-Palmer et al., 2013), and they are likely linked to the cloning process per se.
Although placental morphological and functional analyses have been investigated in more detail in cattle, sheep, and mice, it has been insufficiently explored in goats. In the present study, the comparative morphologic and morphometric data derived from the FM and placentas from the SCNT and AI groups demonstrated significant deviations from normality in cloned specimens, in a similar pattern as seen in cattle (Batchelder et al., 2007; Constant et al., 2006; Ricci et al., 2009). To note, BW, total FM weight and SA, individual and total cotyledonary SA, and the proportions between number of cotyledons and BW, and between BW and cotyledonary SA were markedly altered in the SCNT group. Cloned offspring and FM were heavier and larger, and cotyledons were enlarged, in lower numbers, and with a distinct topographic distribution when compared with the AI group (Tables 4 and 5; Figs. 3 and 4). Nevertheless, the healthy offspring that survived had similar parameters compared with the control groups, whereas a wide range of variability was observed within the SCNT pregnancies. Even though some previous studies did not report placental morphological abnormalities after goat cloning (Keefer et al., 2001, 2002), the reduced number of cotyledons in SCNT-derived placentas was also associated with high gestational and perinatal losses in goats (Lan et al., 2006).
Much of the placental abnormalities that are intrinsic to the SCNT cloning procedures may be due to epigenetic alterations (Humpherys et al., 2001; Mann and Bartolomei, 2002; Smith et al., 2012) and to a faulty gene expression pattern, especially for imprinted genes (Humpherys et al., 2002). Nevertheless, the lack of information about reprogramming failures and the occurrence of abnormalities, added to the increasing interest in the goat model for transgenesis and gene edition, may motivate new studies that are aiming at elucidating and minimizing placental alterations after cloning in goats.
The appearance of a case of hydroallantois in a doe carrying a cloned conceptus in this study adds to the list of failures and phenotypes associated to AOS, which are also often associated with cloning by SCNT in cattle (Batchelder et al., 2007; Cibelli et al., 1998; Constant et al., 2006; Ricci et al., 2009; Zakhartchenko et al., 1999). The large volume (26 L) and osmolarity (279 mOsm/kg) of the allantoic fluid were indicative of hydrallantois. The mean fetal fluid volume in the pregnant goat ranges from 0.5 to 1.5 L, which may have up to a 10-fold increase in cases of fetal hydrops, and the allantoic fluid tends to be hyposmotic relative to plasma osmolarity (Braun, 2007). In fact, the presence of fetal hydrops in in vitro-derived pregnancies has been well documented in the literature (Batchelder et al., 2007; Cibelli et al., 1998; Constant et al., 2006; Ricci et al., 2009; Zakhartchenko et al., 1999), and they are considered one of the major causes of pregnancy loss in the last third of pregnancy (Constant et al., 2006; Edwards et al., 2003). For goats under conventional reproductive conditions, the fetal hydrops has been considered a rare phenomenon with few published reports (Jones and Fecteau, 1995; Morin et al., 1994; Selvaraju et al., 2010).
One of the two transgenic cloned newborns presented a temporary homeostatic disturbance that started in the second day of life ex utero and continued until the end of the second week, comprising dysfunctions in the gastrointestinal, thermoregulatory, and metabolic systems. The other offspring died soon after birth due to cardiorespiratory arrest caused by a cardiac malformation, despite the immediate neonatal support, including oxygen therapy, cardiopulmonary resuscitation, and the use of respiratory-stimulating drugs, as recommended by Meirelles et al. (2010) in cattle and by Chavatte-Palmer et al. (2013) in goats.
The clinical problems observed in the viable neonate have already been described in clones of other species (Chavatte-Palmer et al., 2013; Meirelles et al., 2010), and they are usually related to failures in tissues, organs, or systems that are not challenged or needed during the pre-natal life. Likewise, the post-natal loss in this study is in accordance with other reports, in which death rates during the hebdomadal period can be higher than 50% (Behboodi et al., 2004; Chavatte-Palmer et al., 2013; Keefer et al., 2001; Lan et al., 2006; Meng et al., 2013). As postulated by Barker's hypothesis, the balance of physiologic and metabolic responses in adult life seems to be pre-programed early in embryonic and fetal life (Barker, 1999). Moreover, there are pieces of evidence that nutritional and environmental challenges during the post-natal development exert a negative effect on the oocyte competence and animal fertility (Boland et al., 2001). Such a fetal programming phenomenon, or the fetal origin to adulthood diseases, also appears to impact development after cloning, considering the pre- and post-natal morphological, physiological, and metabolical alterations often seen in cloned concepti.
In summary, a lower cloning efficiency was characterized by a lower pregnancy rate, high gestational losses, and neonatal abnormalities, accompanied by placental anomalies that were compatible and comparable to patterns described in other species. Goat oocytes retrieved from distinct sources showed similar potential as cytoplasts for cloning procedures. However, the cytoplast competence to attain pregnancies and live births was conditioned to the source of oocytes, as IVM oocytes were competent to establish pregnancies, but none reached term. On the other hand, pregnancies after the transfer of cloned embryos using in vivo-matured oocytes was lower, but they resulted in a term pregnancy.
Moreover, the use of non-transgenic control cells yielded higher pregnancy rates than transgenic cells. Irrespective of the cell type or oocyte source, a high incidence of gestational losses was observed after cloning, likely due to failures in the cloning process per se, as described in cattle and sheep. FM derived from cloning presented morphologic and morphometric deviations from normal when compared with the AI control group, and disproportionally bigger newborns in relation to the number of cotyledons. A viable transgenic cloned goat was born from an in vivo-matured oocyte reconstructed with transgenic karyoplasts, being the first transgenic cloned goat born in the tropics of the world. Goat cloning by SCNT still faces technical challenges, but the technology remains of great relevance, especially for the production of GM animals.
Footnotes
Acknowledgments
This study was supported by Grant No. 03.10.0192.00 funded by FINEP/MCT/Brazil. L.T.M. was a Doctorate student at the RENORBIO Graduate Program, supported by a Scholarship granted by CAPES/Brazil.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.