
Abstract
Abstract
Somatic cell nuclear transfer (SCNT) requires large numbers of matured oocytes. In vitro–matured (IVM) oocytes have been used in SCNT in many animals. We investigated the use of IVM oocytes in Rex rabbit SCNT using Rex rabbit ovaries obtained from a local abattoir. The meiotic ability of oocytes isolated from follicles of different diameters was studied. Rex rabbit SCNT was optimized for denucleation, activation, and donor cell synchronization. Rex rabbit oocytes grew to the largest diameter (110 μm) when the follicle diameter was 1.0 mm. Oocytes isolated from <0.5-mm follicles lacked the ability to resume meiosis. More than 90% of these oocytes remained in the germinal vesicle (GV) stage after in vitro culture (IVC) for 18 h. Oocytes isolated from >0.7-mm follicles acquired maturation ability. More than 90% of these oocytes matured after IVC for 18 h. The developmental potential of oocytes isolated from >1-mm follicles was greater than that of oocytes isolated from 0.7- to 1.0-mm follicles. The highest activation rates for IVM Rex rabbit oocytes were seen after treatment with 2.5 μM ionomycin for 5 min followed by 2 mM 6-dimethylaminopurine (6-DMAP) and 5 μg/mL cycloheximide (CHX) for 1 h. Ionomycin induced the chromatin of IVM oocytes to protrude from the oocyte surface, promoting denucleation. Fetal fibroblast cells (FFCs) and cumulus cells (CCs) were more suitable for Rex rabbit SCNT than skin fibroblast cells (SFCs) (blastocyst rate was 35.6 ± 2.2% and 38.0 ± 6.0% vs. 19.7 ± 3.1%). The best fusion condition was a 2DC interval for 1 sec, 1.6 kV/cm voltages, and 40 μsec duration in 0.28 M mannitol. In conclusion, the in vitro maturation of Rex rabbit oocytes and SCNT procedures were studied systematically and optimized in this study.
Introduction
S
Matured oocytes are necessary for cloning and serve as recipients in the reprogramming of the somatic cell nucleus. Oocytes matured in vivo have long been used (Campbell et al., 1996), but their high costs and low efficiency limit their use in SCNT (Long et al., 2014). The cheaper and more convenient oocytes matured in vitro from slaughterhouse-discarded ovaries are often used for SCNT. Using this method, viable cloned offspring have been produced in several species (Choi et al., 2015; Dieci et al., 2013; Lee et al., 2013; Lee et al., 2015; Lin et al., 2015; Pedersen et al., 2014; Sugimoto et al., 2014; Sun et al., 2015). However, to our knowledge, no rabbit offspring have been cloned from in vitro–matured (IVM) oocytes.
Optimal conditions for using the IVM oocytes need to be determined. Different oocytes isolated from different size follicles have different maturation abilities (Sui et al., 2005). Increased fertilization, cleavage, and embryonic development were significantly (p < 0.05) higher in cumulus–oocyte complexes (COCs) aspirated from large follicles, followed by those isolated from medium and small follicles (Khatir et al., 2007). Rabbit oocytes from >1-mm follicles have been cultured for 16 h at 38°C to obtain matured oocytes (Arias-Alvarez et al., 2010). Rabbit oocytes have also been cultured for 24–30 h to evaluate nuclear and ooplasmic maturations (Tao et al., 2008). However, optimal follicle diameters and culture times have yet to be determined for IVM rabbit oocytes.
Nuclear transfer includes many steps. Matured oocytes have been denucleated with Hoechst 33342 in many rabbit nuclear transfer studies (Mitalipov et al., 1999; Yin et al., 2000). Yin et al. denucleated matured oocytes with ionomycin–demecolcine, which caused the nucleus to protrude from the oocyte surface (Yin et al., 2002a). Using this method, Yin et al. obtained the first cloned rabbit. However, it has yet to be determined if this denucleation method can be used for IVM oocytes.
Activation is a key process in nuclear transfer. Most activation steps have been performed using an electric pulse and culturing with 5 mg/mL cycloheximide (CHX) and 2 mM 6-dimethylaminopurine (6-DMAP) for 1 h (Tian et al., 2012; Yang et al., 2013; Yin et al., 2013). Activation via ionomycin was used successfully to clone and bring live sheep to term (Lan et al., 2008). The use of ionomycin in rabbit activation has not been studied systematically. As previously reported, nuclear transferred (NT) embryos reconstructed in fresh cumulus cells (CCs) have better developmental potential than those reconstructed with fetal fibroblast cells (FFCs) in vitro (Tian et al., 2012). However, skin fibroblast cells (SFCs) are more readily obtained. The most suitable cell for rabbit IVM oocyte nuclear transfer remains unclear.
In this study, we optimized the process of SCNT with IVM oocytes. The oocytes were obtained from the ovaries of slaughtered Rex rabbits. The meiosis and maturation abilities of oocytes from different-diameter follicles were studied in detail. We activated oocytes with ionomycin instead of an electric pulse and selected a suitable donor cell type and proper cell cycle synchronization for nuclear transfer with IVM Rex rabbit oocytes. We further studied the effect of oocyte age on rabbit nuclear transfer.
Materials and Methods
Animals and chemicals
Ovaries were obtained from adult Rex rabbits in the Youyi slaughterhouse (Tai'an City, Shandong Province). Rex rabbits were slaughtered using humane slaughter techniques. Ovaries were kept in 25–30°C saline solutions and transported to the laboratory within 2 h of slaughter. All chemicals used for this study were purchased from Sigma unless otherwise stated. The Institutional Animal Care and Use Committee of Shandong Agricultural University approved this study.
Collection of Rex rabbit oocytes
Follicles were measured by a micrometer under a stereomicroscope. They were separated into the following groups according to diameter: <0.5 mm, 0.7–1.0 mm, >1.0–1.5 mm, and >1.5 mm. COCs were released by rupturing follicles in warm Dulbecco's phosphate-buffered saline (D-PBS).
In vitro culture of oocytes and meiosis analysis
COCs collected from the different follicle groups were washed three times in D-PBS before culturing in Tissue Culture Medium-199 (TCM-199; Gibco) supplemented with 10% (vol/vol) fetal calf serum (FCS; Gibco), 1 μg/mL 17β-estradiol, 24.2 mg/L sodium pyruvate, 0.05 IU/mL follicle-stimulating hormone (FSH), 0.05 IU/mL luteinizing hormone (LH), and 10 ng/mL epidermal growth factor (EGF). Twenty COCs were cultured in 100 μL of culture medium, covered with mineral oil, and placed in a constant temperature incubator at 38.5°C and 5% CO2. Cultured oocytes were denuded with a fine-caliber pipette in D-PBS containing 0.1% hyaluronidase. Meiosis was analyzed as previously described (Han et al., 2008). In brief, denuded oocytes were fixed in ethanol:acetic acid (3:1, vol/vol) for 24 h and then stained in 1% aceto-orcein and examined under a phase-contrast microscope.
Parthenogenetic activation
Metaphase II (MII) oocytes were activated by our modified method after 18 h IVC. In brief, the denuded MII oocytes were incubated in different concentrations of ionomycin for 5 min and then placed into culture medium containing 2 mM 6-DMAP and 5 μg/mL CHX or 2 mM 6-DMAP alone for 1 or 2 h, according to the experimental design. Three concentrations of ionomycin were used: 1.25 μM, 2.5 μM, and 5 μM.
Recipient cytoplasm preparation
Cytoplasmic protrusion was induced in denuded MII oocytes with 0.6 μg/mL demecolcine for 40 min. Chromosomes in the cytoplasmic protrusion (Fig. 1) and the first polar body were removed by an enucleating pipette. The micromanipulation liquid was D-PBS containing 7.5 mg/mL cytochalasin B.
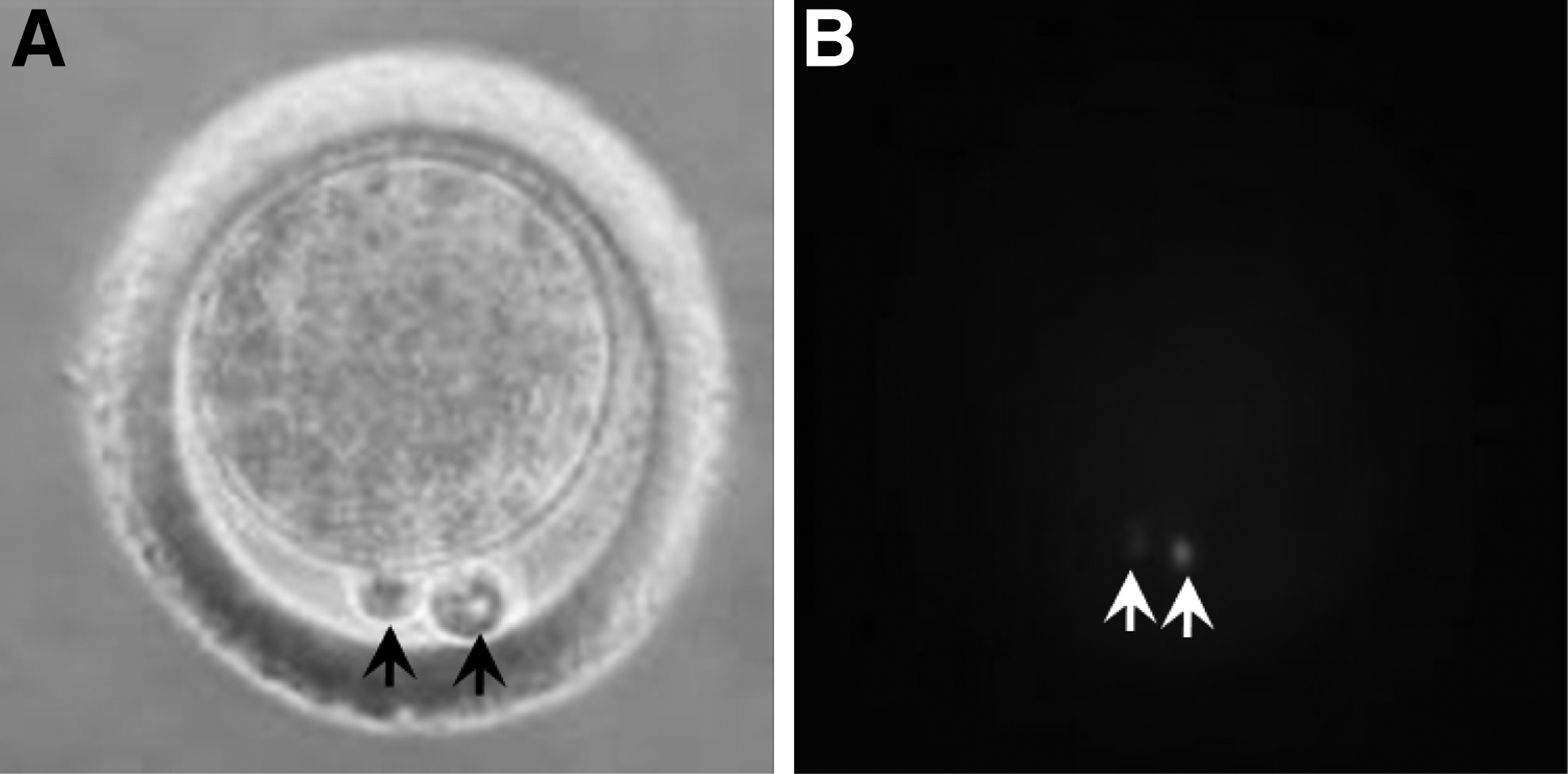
Chromosomes were in the cytoplasmic protrusions after demecolcine induction. (
Preparation of donor somatic cells
Three types of donor somatic cells were used in this research. Skin fibroblast cells were obtained from ear skin tissue, fetal fibroblast cells were obtained from fetuses at 17 days of pregnancy, and CCs were obtained from follicles and were used directly as donor cells. All fetuses and tissues were obtained from adult Rex rabbits in the Youyi slaughterhouse. SFCs and FFCs were obtained by similar methods. In brief, the tissue was first washed several times and then cut into 1-mm3 fragments in Dulbecco's Modified Eagle's Medium (DMEM) with 10% FBS. Cells were digested for 30 min (SFCs) or 3–5 min (FFCs) in trypsin-EDTA solution at 38.5°C. Cell suspensions were obtained after terminating digestion. Cells were cultured in DMEM supplemented with 10% FBS at 38.5°C under 5% CO2 in humidified air. The cell cycle was regulated both by serum starvation and contact inhibition. For serum starvation, cells were cultured in DMEM with 0.5% FCS for 1–3 days. For contact inhibition, cells were cultured for 1–3 days after the cells became 95% confluent.
Nuclear transfer, fusion, and embryo culture
Donor cells were digested into single cells before nuclear transfer. A single donor cell was injected into the perivitelline space of the recipient cell's cytoplasm. The micromanipulation liquid was the same as the enucleation liquid. Reconstructed embryos were treated with various voltages in different concentrations of mannite. Fused embryos were stimulated with 2.5 μM ionomycin for 5 min, followed by 2 mM 6-DMAP and 5 μg/mL CHX for 1 h.
Statistical analysis
We repeated each treatment at least three times. Data were compared using one-way analysis of variance (ANOVA) after being transformed via least significant difference (LSD). p < 0.05 was considered significant.
Results
Diameters of oocytes from different size follicles
Oocyte diameters increased with follicle size and reached a maximum diameter (133.1 ± 0.7 μm) when the follicles were larger than 1 mm. Among the four groups, the oocyte diameter from the <0.5-mm group was the smallest at approximately 90 μm, but the zona pellucida (ZP) was the thickest. As the follicle diameter increased, the diameter of the oocyte also gradually increased, but the ZP became thinner. When the follicle diameter was 1.0 mm, oocytes attained their largest diameter (over 110 μm) and the ZP thickness stabilized at approximately 10 μm (Table 1).
Oocyte diameter does not include zona pellucida (ZP).
The same letters within the same column indicates no significant difference, i.e., p > 0.05.
Meiosis in oocytes collected from different-diameter follicles
Oocytes isolated from the <0.5-mm group could not resume meiosis, but oocytes isolated from the 0.7- to 1.0-mm, >1.0- to 1.5-mm, and >1.5-mm groups resumed meiosis and matured. Meiosis is shown in Figure 2A. Despite having similar maturation abilities, the oocytes from the three larger follicle groups had different maturation speeds. According to the method presented by Sirard et al. (1989), we calculated areas under the curve, as shown in Figure 2B. The percentage of each area was calculated relative to the total area, which represents the duration of meiosis. The oocytes from the 0.7- to 1.0-mm group matured more slowly than the oocytes from the >1.0- to 1.5-mm, and >1.5-mm groups (Fig. 2).
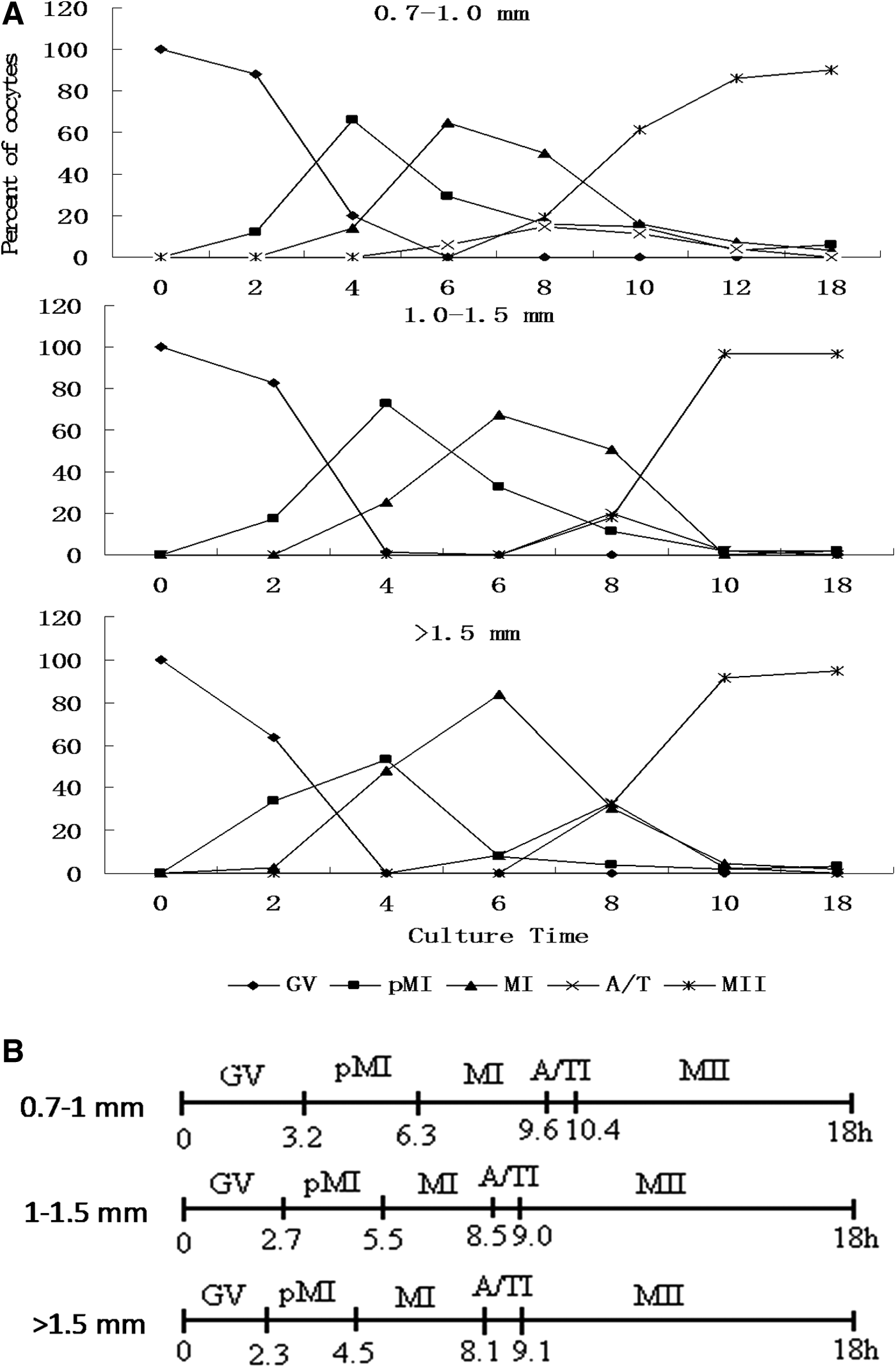
Meiosis in Rex rabbit oocytes isolated from differently sized follicles. (
Optimizing the Rex rabbit oocyte chemical activation method
Because the oocytes collected from >1.0-mm follicles were fully grown, we carried out the following experiments using these oocytes. Initially, oocytes were treated with 2.5 μM ionomycin for 5 min followed by 2 mM 6-DMAP for 2 h. We adjusted this method as follows.
First, we adjusted the method to minimize the activity of maturation-promoting factor/cytostatic factor (MPF/CSF). As shown in Table 2, oocytes matured in vitro for 18 h were activated with 2.5 μM ionomycin for 5 min and were then subjected to four different treatment conditions. Oocytes that were restrained in 6-DMAP and 5 μg/mL CHX for 1 h attained a 53% blastocyst rate, significantly higher than the other three groups (p < 0.05). There were no significant differences in the cleavage rate between the four groups.
The same letters within the same column indicates no significant difference, i.e., p > 0.05.
6-DMAP, 6-dimethylaminopurine; CHX, cycloheximide.
Three concentrations of ionomycin were used to induce calcium oscillations. At 1.25 mM, ionomycin did not activate the oocytes effectively. The cleavage rate was only 49.4% and the blastocyst rate was only 12.5%. When the ionomycin concentration increased to 2.5 mM, the cleavage rate increased to 82.0% and the blastocyst rate increased to 55.1%. No significant effects were observed when the ionomycin concentration was increased to 5 mM (Table 3).
The same letters within the same column indicates no significant difference, i.e., p > 0.05.
6-DMAP, 6-dimethylaminopurine; CHX, cycloheximide.
Demecolcine-assisted enucleation method
We next designed experiments to study the effects of chemical-assisted enucleation with demecolcine on Rex rabbit SCNT. Treatments with 0.6 μg/mL demecolcine for 1 h induced the oocytes to form cytoplasmic protrusions containing chromatin. The efficiency was observed to be affected by oocyte age. As shown in Table 4, younger oocytes had the lowest protrusion formation rates. Only 32.6% of young oocytes formed cytoplasmic protrusions when induced after 12 h of in vitro maturation. However, the protrusion formation rate increased to 86.7% when treatment was delayed to 14 h and to 90.3% in oocytes matured in vitro for 18 h.
The same letters within the same column indicates no significant difference, i.e., p > 0.05.
Donor somatic cell type and cycle affect the efficiency of reprogramming in IVM oocytes
The donor somatic cell type and cycle can affect the reprogramming efficiency of oocyte cytoplasm–transferred nuclei in the SCNT experiments. We used three types of somatic cells as nuclear donor cells—CCs, SFCs from the ear, and FFCs. As shown in Table 5, no significant differences were observed between the development of blastocysts in CCs and FFCs (∼36%).
The same letters in the same column indicates no significant difference, i.e., p > 0.05.
SFCs, skin fibroblast cells; FFCs, fetal fibroblast cells; CCs, cumulus cells.
However, the development of blastocysts was significantly lower in SFCs (19.7%). We next compared the effects of serum starvation and contact inhibition on cell cycle synchronization in Rex rabbit CCs (Table 6). Serum starvation synchronized CCs more rapidly.
Optimizing the fusion conditions of CCs with IVM oocytes
Fusion was performed with various voltages for different concentrations of mannite. When oocyte cell fusion was performed in 0.3 M mannitol, a 40-μsec double electrical pulse of 1.2 kV/cm intervals 1 sec resulted in a 60% fusion rate. When the voltage was increased to 1.5 kV/cm, the fusion rate increased to 75%. However, when the voltage was increased to 1.6 kV/cm, 75% of the oocytes died. When fusion was performed in 0.28 M mannitol, the fusion rate reached 85% with 1.5 kV/cm (Table 7).
The same letters within the same column indicates no significant difference, i.e., p > 0.05.
Effect of oocyte age
Oocytes cultured for 12, 14, or 18 h were used to investigate the relationship of culture time and the development of reconstructed embryos. For oocytes cultured for 12 h, the cleavage rate of the reconstructed embryo was 74.5%, the blastocyst rate was 30.1%, and the hatching rate was 70.8% (Fig. 3). For oocytes matured in vitro for 14 and 18 h, the cleavage rates reached 83.2% and 89.7%, respectively. There were no significant differences among the three groups (Table 8). The blastocyst and hatching rates also did not significantly differ among the three groups.
Hatched blastocyst cultured for 6 days.
The same letters in the same column indicates no significant difference, i.e., p > 0.05.
6-DMAP, 6-dimethylaminopurine; CHX, cycloheximide.
Discussion
Research on rabbit cloning from embryo cells began in 1975, and cloned blastocysts were obtained. However, no cloned offspring were born until 1988. Since then, Stice and colleagues cloned six rabbits using eight-cell embryo nuclei as nuclear donors. In 2002, a French research group successfully cloned rabbits from CCs (Yin et al., 2002a). Although they produced viable offspring, the recipient oocytes were collected in vivo by superovulation. One major objective of our research is to use oocytes matured in vitro as cytoplasmic recipients to produce cloned rabbits.
We first established an in vitro maturation system for Rex rabbit oocytes. Ovaries from the slaughterhouse were not qualitatively homogeneous. We compared the diameters and developmental abilities of oocytes collected from different follicles. The oocytes from <0.5-mm follicles lacked the ability to resume meiosis. Approximately 90% of these oocytes were in the germinal vesicle (GV) stage after 18 h of in vitro culture. Approximately 50% of the GV oocytes obtained from 0.5- to 0.7-mm follicles matured after 18 h of in vitro culture. This conclusion concurs with the results of Smith et al. (1978). When the follicles grew larger than 0.7 mm, the oocytes acquired complete maturation ability. More than 90% of oocytes were able to mature after 18 h of in vitro culture. Similar observations were seen in sheep. Sheep oocytes obtained from 0.5- to 0.8-mm follicles could resume meiosis. The 1- to 1.8-mm follicular oocytes achieved the metaphase I (MI) stage. Oocytes obtained from >2-mm follicles can achieve MII (Ma et al., 2003).
Although oocytes obtained from the >0.7-mm follicles were able to mature, their maturation speeds differed greatly. The 0.7- to 1.0-mm follicular oocytes achieved maximum maturation with 12 h of in vitro maturation, and the >1.0-mm follicular oocytes only needed 10 h to achieve maximum maturity. Yin and his colleagues indicated that 89% of oocytes can mature after 10 h of in vitro maturation (Yin et al., 2002a), consistent with our findings. Yin and colleagues also reported that 78% of oocytes matured after 8 h of in vitro maturation (Yin et al., 2002a). However, our results showed that only 17.7% of oocytes obtained from the >1.5-mm follicles could mature after 8 h of in vitro maturation, and only 32.4% of oocytes obtained from the >1.0- to 1.5-mm follicles could mature after 8 h of in vitro maturation. These contradictory results are likely due to the different oocyte sources. Oocytes were obtained from superovulated rabbits in Yin et al.'s experiment, but our oocytes were obtained from abattoir ovaries. Additionally, hormone stimulation may affect rabbit oocyte maturation speed and quality. Early research has shown that gonadotropin influences the maturation competence of MII bovine oocytes (Wang et al., 2014).
In MII oocytes pretreated with ionomycin–demecolcine, chromatin aggregated within bulges on the oocyte surfaces. These bulges were easily identified and removed by pipetting with a glass needle. By using this method, a cloned pig was born and a cloned rabbit fetus was obtained (Yin et al., 2002a, 2002b). We attempted to induce the bulge with demecolcine alone in IVM oocytes. Our results showed that demecolcine was not effective in oocytes cultured for 12 h in vitro. Only one-third of the oocytes formed bulges. However, with oocytes undergoing >15 h of in vitro maturation, demecolcine induced >80% oocytes to form bulges. We denucleated 100% of oocytes with this method. The angle of deviation between the bulge and the first polar body was greater than 30° in 65% of oocytes (data not shown). This may be one reason for low rabbit cloning efficiency.
When we employed denucleated IVM oocytes as cytoplasm, the cleavage rate did not differ significantly among cloned embryos that used ear SFCs, FFCs, or CCs as donor cells. However, the blastocyst rates of the FFC and CC groups were significantly higher than that of the SFC group. In a previous study, NT embryos reconstructed with fresh CCs (CC-embryos) had better developmental potential than those reconstructed with fetal fibroblasts (FF-embryos) (Tian et al., 2012). Oocytes obtained from in vivo or in vitro maturation may also be affected by the efficiency of the reprogramming. We synchronized the donor cells using contact inhibition and serum starvation. Flow cytometry results showed that the G0/G1 ratio was higher in the serum starvation group.
Although activation with ionomycin has been widely used in many animals and has resulted in successful cloned animal births (Lan et al., 2008), there is no existing systematic study on how to use ionomycin to activate rabbit oocytes. Hirabayashi and his colleagues activated rabbit oocytes injected with round spermatids with ionomycin, and rabbits were successfully born (Hirabayashi et al., 2009). We used 1.25 μM, 2.5 μM, and 5 μM ionomycin to activate oocytes. Ionomycin was not able to activate Rex rabbit IVM oocytes effectively at 1.25 μM. However, 2.5 μM and 5 μM ionomycin were both effective. More than 50% of oocytes developed into blastocysts, and more than 60% of blastocysts hatched. This method improved on previous studies (Mitalipov et al., 1999). We incubated activated embryos in TCM-199 containing 5 μg/mL CHX and 2 mM 6-DMAP for different times. The blastocyst rate was significantly higher in the 1-h incubation group than in the 2-h group (53.0% vs. 28.8%, p < 0.05). This may be because 72% of the microtubules were in interphase in nuclear transfer embryos treated with 5 μg/mL CHX and 2 mM 6-DMAP for 1 h (Chesne et al., 2002). Incubations with both 5 μg/mL CHX and 2 mM 6-DMAP provided better results than incubations with 2 mM 6-DMAP alone
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant nos. 31201789, 31272444, 31372273, and 31101715); the funders had important role in study design, data collection, and analysis. The Natural Science Fund Project in Anhui province (grant no. 1508085QC59) and the major project of Biology Discipline Construction in Anhui province (2014) had the role in decision to publish or preparation of the manuscript.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.