
Abstract
Abstract
Insulin replacement is the current therapeutic option for type-1 diabetes. However, exogenous insulin cannot precisely represent the normal pattern of insulin secretion. Another therapeutic strategy is transplantation of pancreatic islets, but this is limited by immune rejection, intrinsic complications, and lack of donor availability. Stem cell therapy that results in the regeneration of insulin-producing cells represents an attractive choice. However, with advancing age, stem cells also undergo senescence, which leads to changes in the function of various cellular processes that result in a decrease in the regeneration potential of these aging stem cells. In this study, the effect of young and aging mesenchymal stem cells (MSCs) on the regeneration of pancreatic beta cells in streptozotocin (STZ)-induced type-1 diabetic mice was observed after hypoxic preconditioning. Hypoxia was chemically induced by 2, 4-dinitrophenol (DNP). Plasma insulin and glucose levels were measured at various time intervals, and pancreatic sections were analyzed histochemically. The effect of DNP was also analyzed on apoptosis of MSCs by flow cytometry and on gene expression of certain growth factors by quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). We observed that hypoxic preconditioning caused changes in the gene expression levels of growth factors in both young and aging MSCs. Young MSCs showed significant regeneration potential compared with the aging cells in vivo. However, hypoxic preconditioning was able to improve the regeneration potential of aging MSCs. It is concluded from the present study that the regeneration potential of aging MSCs into pancreatic β-cells can be enhanced by hypoxic preconditioning, which causes changes in the gene expression of certain growth factors.
Introduction
D
Insulin therapy is the standard treatment option for type-I diabetes. Different regimes of insulin injections are prescribed with careful monitoring of blood glucose levels. However, good metabolic control is not easy to maintain. Severe hypoglycemic episodes are frequently associated with insulin injections, because exogenously supplied insulin does not represent the normal physiology of insulin secretion (Jia et al., 2003). Another therapeutic option is the β-cell replacement using transplantation of either whole pancreas or islets (Jia et al., 2003; Jiping et al., 2006; Vija et al., 2009). Islet transplantation has a major disadvantage of having ethical concerns, immune rejection, as well as the quality and quantity of cells (Jia et al., 2003; Jiping et al., 2006).
In this perspective, regeneration of β-cells can be achieved by self-replication of β-cells or by differentiation of progenitor cells by using stem cell therapy. This therapy can also overcome the problems of autoimmunity and results in improved secretion of endogenous insulin (Vija et al., 2009). It has been demonstrated that endodermal-like cells can be produced from embryonic-like stem cells and can be subsequently differentiated into insulin-secreting β-cells both in vitro and in vivo (D'Amour et al., 2006). The understanding of a number of transcription factors and extracellular signals involved in the embryonic development of pancreas has led scientists to this remarkable achievement (Oliver-krasinski and Stoffers, 2008). However, embryonic-like stem cells have some limitations for their use in regenerative medicine. These cells can lead to teratoma formation and immune rejection, and they also have ethical concerns. On the other hand, mesenchymal stem cells (MSCs) can be an attractive therapeutic option due to their potential of differentiating into pancreatic insulin-secreting cells in vitro (Matsumoto et al., 2006; Oliver-krasinski and Stoffers, 2008; Vija et al., 2009).
It has been observed that with advancing age, stem cells are subject to insults at the molecular level from various extra- and intracellular sources, and, thus, these aging cells lose their trans-differentiation potential. An understanding of the intrinsic mechanism of age-related decrease in the regeneration potential of aging stem cells might result in the development of novel treatment strategies (Kasper et al., 2009).
The regeneration potential of aging MSCs can be enhanced by utilizing preconditioning strategies in which cells are exposed to non-cytotoxic stress before transplantation. Preconditioning leads to the stimulation of endogenous mechanisms, resulting in multiple responses, including activation of specific cell surface receptors by growth factors and cytokines (Takahashi et al., 2006; Wisel et al., 2009).
One of the promising preconditioning strategies is the treatment of MSCs with DNP, a metabolic inhibitor that depletes the energy recourses of cells. DNP uncouples oxidative phosphorylation from ATP production by facilitating proton flux back into the mitochondrial matrix (Loomis et al., 1948), thus inhibiting intracellular phosphorylation of glucose (Morgan and Street, 1959). DNP is shown to produce an anaerobic environment by a mechanism that is distinct from low-tension hypoxia. DNP-treated adhesion fibroblasts were shown to upregulate the expression of VEGF but not hypoxia-inducible factor (HIF1α), suggesting that it works through the HIF-independent pathway (Shavell et al., 2012). In the present study, we analyzed the effect of DNP on young and aging MSCs with reference to their differentiation potential into pancreatic beta cells after type-1 diabetes in mice. The effect of DNP was also explored in terms of its ability to induce the expression of certain growth factors. The present study would provide an insight into the hypoxic preconditioning strategy of aging MSCs as a possible future therapeutic approach against diabetes.
Materials and Methods
Animals
All animal procedures were carried out in accordance with the International guidelines for the care and use of laboratory animals after the approval of the local ethical committee. For all experiments, NMRI mice were divided into two groups according to age and weight. Group A consisted of 4- to 6-week-old mice (20–25 g; young), and group B comprised 32- to 40-week-old mice (35–40 g; aging).
Isolation and propagation of young and aging MSCs
Whole bone marrow was flushed from tibia and femur of NMRI mice and cultured in Dulbecco's modified Eagle's medium (GIBCO) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. Culture was maintained at 37°C in a humidified condition having 5% CO2. Non-adherent hematopoietic cells were removed from adherent MSCs. On reaching ∼70% confluence, the cells were sub-cultured with 0.25% trypsin-EDTA. MSCs of passages 1–2 were used throughout this study.
Characterization of young and aging MSCs by flow cytometry
Cultured MSCs were analyzed for the presence of surface markers CD90, CD44, CD117 and negative marker, CD34. Cell dissociation solution was added to each flask and incubated at 37°C for 15–20 minutes. Dissociated cells were centrifuged at 400 g for 5 minutes. Four hundred microliters of FACS solution (1% BSA, 1 mM EDTA, 0.1% sodium azide in PBS) was added to the pelleted cells. One hundred microliters each of this cell suspension containing 1 × 106 cells was added to 5 mL FACS tubes. Five microliters of blocking solution (1% BSA in PBS) was added to each tube and incubated for 2 minutes at room temperature. The cells were incubated separately with specific primary antibodies in 1:40 dilution on ice for 30 minutes. Cells were washed three times with 2 mL FACS solution, vortexed, and centrifuged at 400 g for 5 minutes at room temperature. Alexa fluor 546 goat anti-mouse secondary antibody (1:500) was then added to each tube, and tubes were kept on ice for 30 minutes. Supernatant was removed, and 500 μL FACS solution was added to each tube. Cells were counted in a flow cytometer and analyzed by CellQuest Pro software (FACSCalibur™; Becton, Dickinson and Co.).
Analysis of growth factors in young and aging MSCs in response to hypoxia
Hypoxia induction
Optimization of DNP concentration is described in detail elsewhere (Ali et al., 2015; Haneef et al., 2014). Briefly, a range of 0.1–1 mM concentration of DNP was used for different periods. The concentration that showed minimum cytotoxicity and less effect on cell morphology was selected for further study. Young and aging MSCs were treated separately with an optimized concentration of 0.5 mM DNP for 10 minutes. The cell culture medium containing DNP was replaced by normal medium, and cells were re-oxygenated for 24 hours by incubating them at 37°C in a humidified incubator with 5% CO2. Untreated cells were used as control. Normal and hypoxic cells were used in the subsequent experiments for analysis of apoptosis by flow cytometry and growth factor expression analysis by qRT-polymerase chain reaction (PCR).
Analysis of growth factors by qRT-PCR
Total RNA from control and treated groups were isolated using the Trizol method according to the manufacturer's protocol. The quality and purity of the isolated total RNA was quantified with a spectrophotometer at 260 and 280 nm. A260: A280 ratio of 1.9–2.0 was considered acceptable for further experiments. RNA was reverse transcribed into cDNA using the Revert Aid First Strand cDNA kit according to the manufacturer's instructions. cDNA was amplified by qRT-PCR using the Go Taq qPCR master mix as recommended by the manufacturer. The primer sequences, PCR annealing temperatures, and expected product sizes are listed in Table 1. Mice GAPDH gene was used as an internal standard. The RT-PCR reactions were subjected to amplification by PCR using the program: (1) denaturation: 96°C for 30 seconds; (b) amplification: 45 cycles of denaturation at 96°C, annealing at 58°C, and extension at 72°C for 30 seconds each; and (c) final extension: 72°C for 10 minutes. Fold change in gene expression pattern was calculated from delta cycle threshold (ΔCt) value and plotted.
Apoptosis assay by flow cytometry
Young and aging MSCs with or without DNP treatment were trypsinized, and the dissociated cell suspension was centrifuged at 120 g for 5 minutes. The cells were washed twice by 1 × PBS and centrifuged at the same speed. The supernatant was decanted, and the pelleted cells were resuspended in 400 μL of buffer containing PBS, serum, and sodium azide. One microliter of propidium iodide (PI) was added to the cells, mixed gently, and incubated for 1 minute in the dark. The PI fluorescence using the FL-2 channel was determined with the flow cytometer.
Effect of hypoxia on the regeneration potential of young and aging MSCs
Type-1 diabetic model
Type-1 diabetes was induced in male NMRI mice aged 4–6 weeks as previously described (Breyer et al., 2005), with some modifications. Streptozotocin (STZ; 50 mg/kg) was dissolved in citrate buffer, pH 4.5 and injected intraperitoneally (i/p) as multiple low doses for 5 consecutive days. Control mice were injected with citrate buffer i/p. Plasma insulin and glucose levels were measured at various time intervals Blood was drawn from the tail vein both before and after the induction of diabetes as well as from the control animals. The animals whose blood glucose level was above 16.67 mM at day 15 were considered type-1 diabetic.
Cell transplantation and tracking
For cell tracking, young and aging MSCs were labeled before transplantation with cell membrane labeling dye, PKH26 (Sigma) according to the manufacturer's protocol. Briefly, 1 × 106 cells/100 μL were injected in the diabetic mice through tail vein at day 10 after the completion of STZ treatment. Control mice were injected with normal saline. Mice were sacrificed by an overdose (200 μL; i/p) of pentothal sodium (Capitelli et al., 2014) at day 10 post-transplantation. The pancreas was dissected. Optimal Cutting Temperature (OCT) compound is used as an embedding medium for frozen tissue samples before cryosectioning. OCT-containing tissue was frozen in isopropanol precooled in liquid nitrogen. Eight- to 16 μm sections were cut using cryotome (Thermo Electron Corporation). The sections were visualized by fluorescence microscopy.
Histochemical analysis
Histological sections were stained as described earlier (Hafizur et al., 2011). Briefly, the pancreas was dissected, kept overnight in 4% neutral buffered formalin, washed, dehydrated, and embedded in paraffin wax. Six-micrometer sections were cut using microtome and transferred to polylysine-coated slides. Antigen retrieval and rehydration was performed as previously described (Hafizur et al., 2011). Sections were stained with eosin and hematoxylin. Stained slides were analyzed by microscopy.
Statistical analysis
Data are presented as mean ± standard error of mean (SEM). Statistical analysis was performed by using one-way ANOVA and Bonferroni's or Tukey's post hoc test for multiple comparisons. Values of p < 0.05 were considered statistically significant.
Results
Young and aging MSCs
Passage 1 cultures of young and aging MSCs showed a heterogeneous population of spindle and fibroblast-shaped cells. Aging MSCs were slightly bigger, having less spindle morphology with a more flattened shape as compared with young MSCs (Fig. 1).
BM-MSCs at Passage 1 (P1) isolated from
Characterization of young and aging MSCs by flow cytometry
Flow cytometric analysis of young and aging bone marrow-derived MSCs showed that CD90- and CD44-positive cells were more in young as compared with aging cells, whereas c-kit or CD117-positive cells were slightly more positive in case of aging cells. CD34, a negative marker for MSCs, was negative in the majority of these cells (Fig. 2).
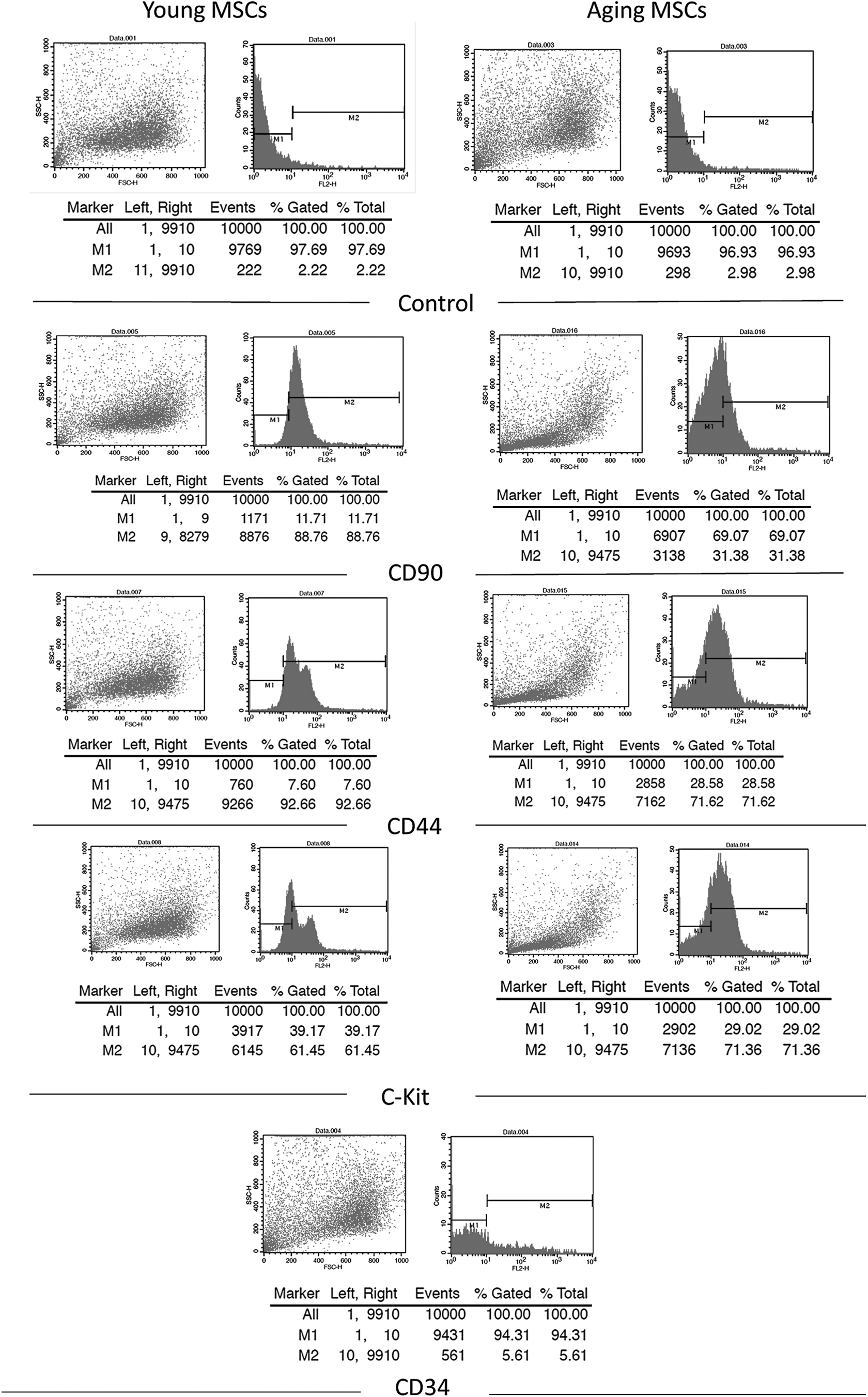
Flow cytometric analysis of young and aging MSCs showing positive staining for CD90, CD44, and CD117 and negative staining for CD34. Alexa fluor goat anti-mouse 546 secondary antibody was used for detection. FSC was selected as the threshold parameter and the threshold was set to a value of 52, which eliminates small debris. FSC, forward scatter; MSCs, mesenchymal stem cells.
Analysis of growth factors in young and aging MSCs in response to hypoxia
Hypoxia induction
For hypoxia induction, an optimized concentration (0.5 mM) of DNP was selected. MSCs were treated with DNP for 10 minutes. At this concentration, cells were slightly shrunken but regained normal morphology after re-oxygenation; whereas at higher concentrations, almost all cells were dead (Fig. 3).
Young and aging MSCs showing
Expression levels of growth factors
We analyzed the mRNA expression levels of angiopoietin-1 (Ang-1), fibroblast growth factor (FGF), hepatocyte growth factor (HGF), transforming growth factor β-1 (TGFβ-1), and insulin-like growth factor (IGF) by qRT-PCR in normal and hypoxic MSCs after 24 hours of re-oxygenation. A comparison between young and aging cells shows significantly higher (*p < 0.01) expression levels of angiopoietin-1 and significantly lower (*p < 0.01) expression levels of FGF and IGF in aging MSCs as compared with young normal MSCs. Hypoxic preconditioning caused changes in the gene expression levels of IGF, FGF, and TGFβ-1. IGF levels were significantly decreased (*p < 0.001) in both young and aging hypoxic MSCs, whereas FGF level was significantly decreased (*p < 0.01) in aging hypoxic MSCs. A non-significant increase in TGFβ-1 expression was observed in aging hypoxic MSCs as compared with normal cells (Fig. 4).
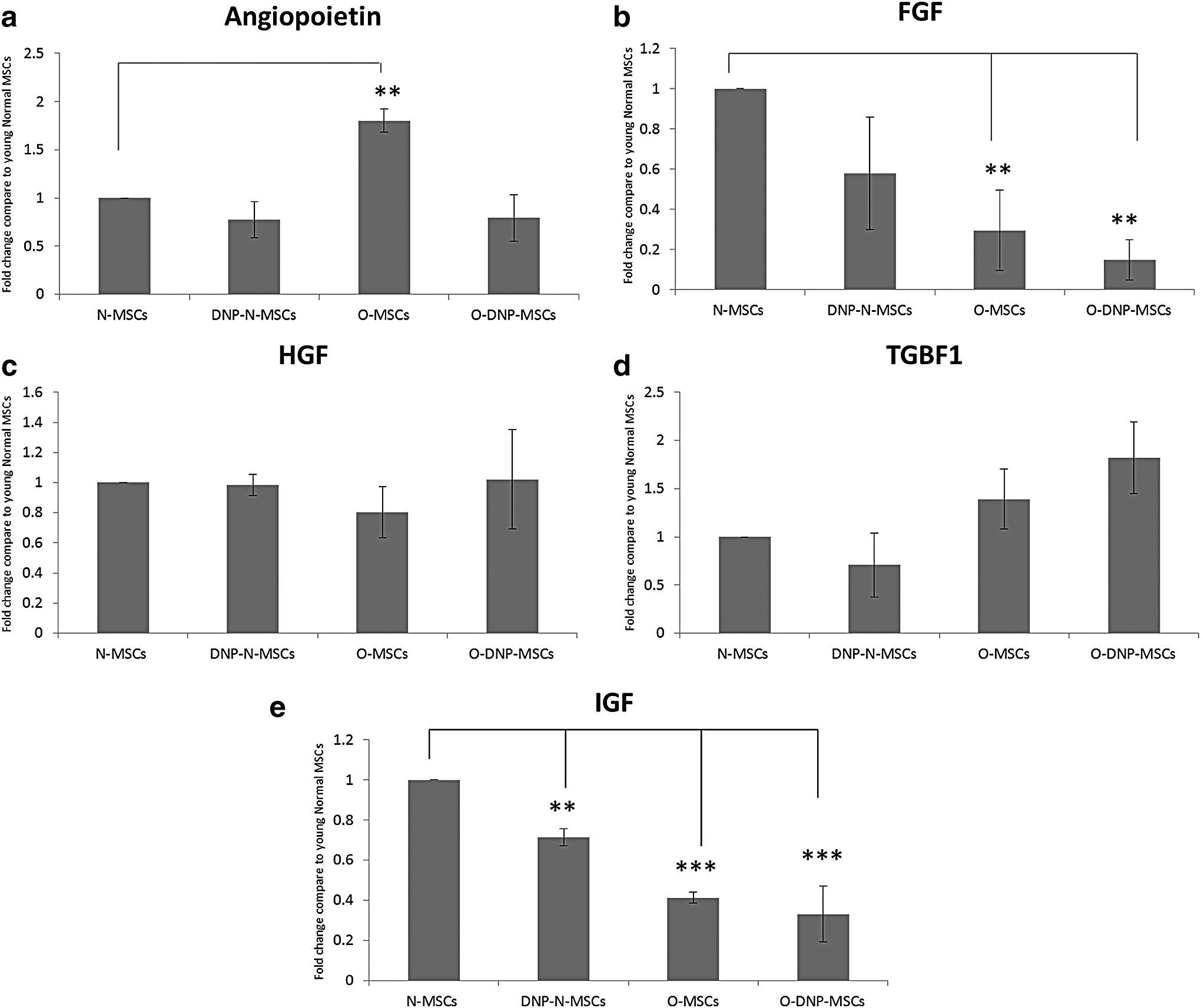
Bar diagrams showing fold changes in expression by quantitative polymerase chain reaction analysis of
Apoptosis assay by flow cytometry
The number of apoptotic cells is slightly more in case of aging MSCs as compared with normal young MSCs. However, there seems to be no significant difference in case of both normal and aging cells. No effect of DNP is seen as well (Fig. 5).
Flow cytometric analysis of apoptosis in normoxic and hypoxic young and aging MSCs by PI. Hypoxia was induced by 0.5 mM DNP for 10 minutes, and cells were re-oxygenated for 24 hours. Both normoxic
Effect of hypoxia on regeneration potential of young and aging MSCs
Elevation of fasting blood glucose concentration was evident at day 7. STZ-treated mice were found to have a blood glucose concentration >16.67 mM on the 15th day. Destructive diabetes was also confirmed by the histological study of pancreas of STZ-treated mice as compared with control (Fig. 6). A significant decrease in the size of pancreatic islets was observed in the treated pancreatic tissue section (Fig. 6a–g). Fewer fluorescently labeled aging MSCs were detected in the frozen pancreatic sections of the treated mice pancreas as compared with that in the case of young MSCs (Fig. 6h, i). Insulin immunostaining was performed in normal, diabetic, and MSCs-treated groups. It is found that diabetic animals that received normal and DNP-treated MSCs preserved islets from STZ-induced degeneration (Fig. 6j–o). The fasting blood glucose levels were monitored at 2-day intervals up to 10 days. An examination of fasting blood glucose levels of mice treated with young MSCs showed a decrease in glucose levels as compared with diseased control (Fig. 7).
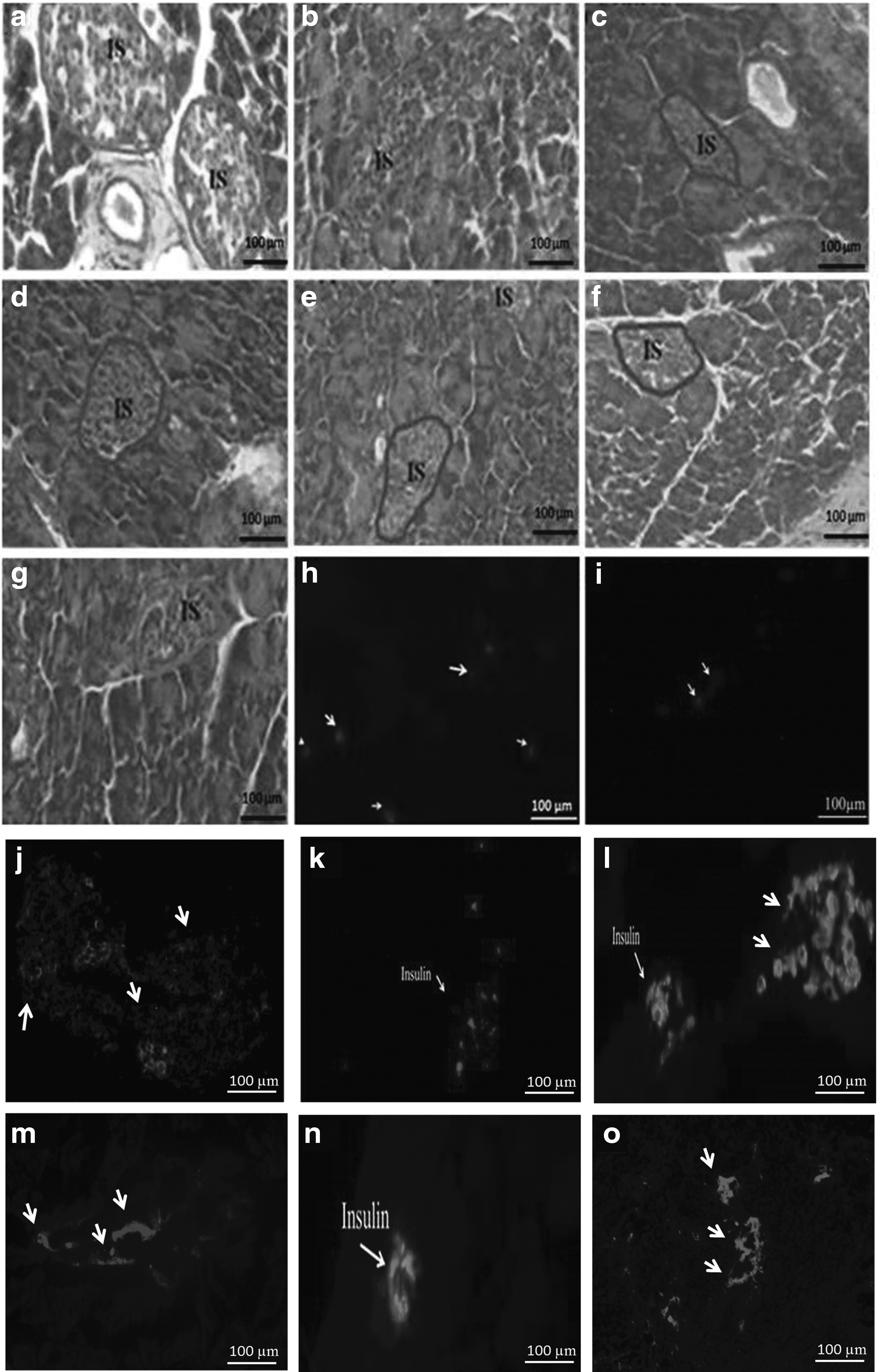
H/E staining of
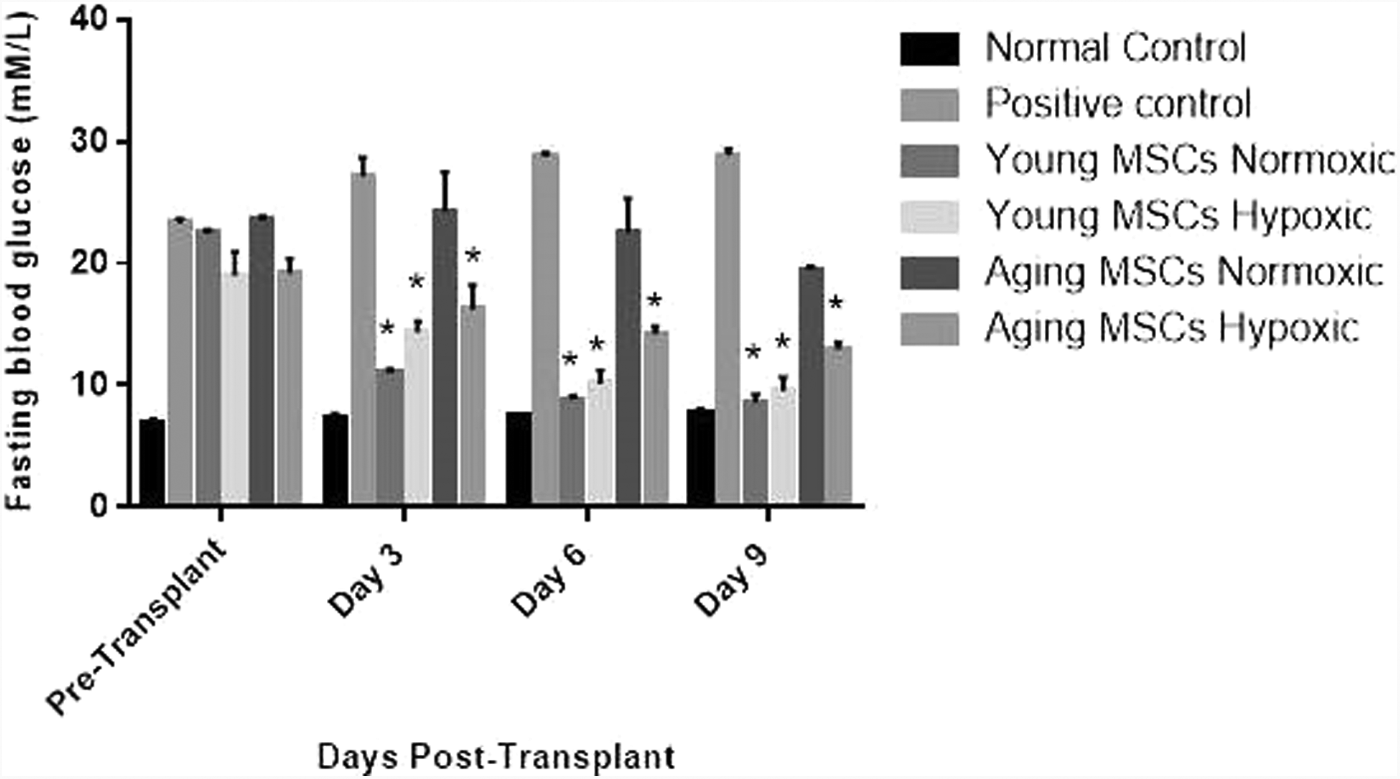
Bar diagrams showing fasting blood glucose levels (mM) in normal, diabetic control, and treated mice. Data represent mean ± SEM for n = 4. One-way ANOVA followed by Bonferroni's post hoc test was performed with level of confidence *p < 0.05. SEM, standard error of mean.
In case of transplantation of aging MSCs, normal glucose levels were not restored in treated mice as compared with control. We found a significant decrease in insulin concentration up to day 16 compared with the normal group. A significant increase in insulin concentration was observed in all the treated groups as compared with diseased control except in case of the normal aging MSCs (Fig. 8).
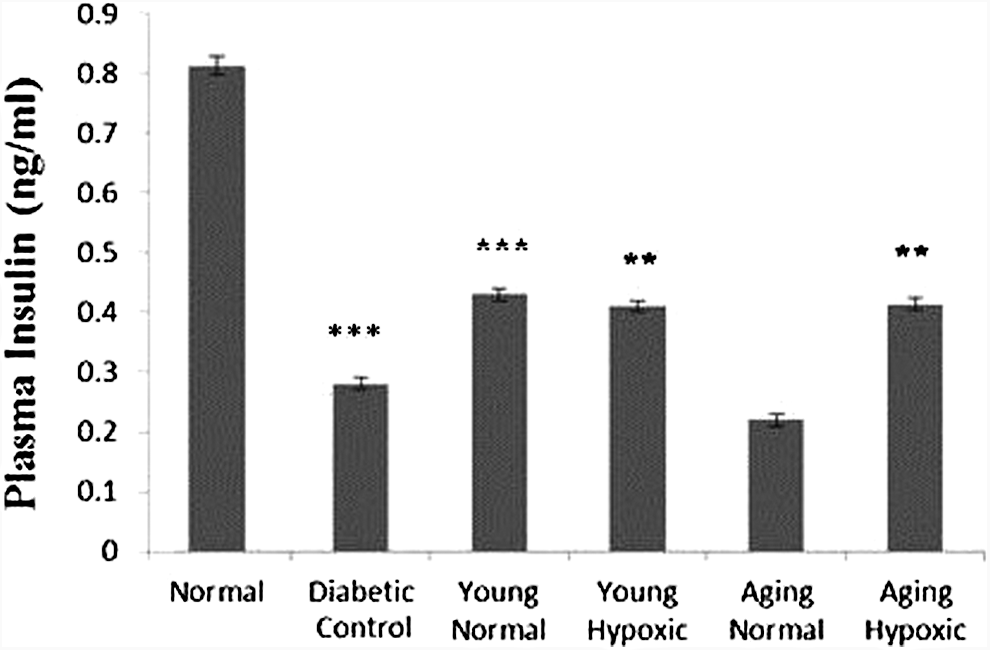
Bar diagram showing blood insulin levels (ng/mL) in normal, diabetic control, and treated mice. Data represent mean ± SEM for n = 4. One-way ANOVA followed by Bonferroni's post hoc test was performed with level of confidence p < 0.05 as statistically significant and **p < 0.01, ***p < 0.001.
Discussion
Pancreatic or islet transplantation can be a reliable source of insulin in type-1 diabetes, but this therapeutic approach is limited by the immune rejection and scarcity of organ donors (Wang et al., 2013). An alternative therapeutic option is stem cell therapy. In this perspective, bone marrow MSCs are the preferred cell type because of their easy access and culturing. However, biological age-related changes occur in MSCs, which result in the decrease in their therapeutic potential and viability (Shen et al., 2011; Yu et al., 2001). As stem cells age, their proliferation rate is also reduced. The number of MSCs in culture is also decreased in older donors as compared with young donors (Baxter et al., 2004; Zhou et al., 2008). To improve the regeneration potential of aging stem cells, one can use various preconditioning strategies.
Preconditioning of stem cells with hypoxic shock, growth factors, and anti-aging compounds signifies an effective approach to enhance proliferation, survival, and differentiation of MSCs (Khan et al., 2013; Mohsin et al., 2011). Among these, the role of a low-oxygen microenvironment is quite significant. Hypoxic preconditioning not only helps stem cells to survive in the injured area but also modulates the expression of various tissue-protective cytokines and growth factors (Muscari et al., 2013).
In this study, we report the effect of hypoxic preconditioning on young and aging MSCs on the expression levels of certain growth factor genes, as well as on the islet cell regeneration in vivo after transplanting these cells into the STZ-induced diabetic mice. We induced chemical hypoxia (Jovanović et al., 2009) using an optimized dose of DNP in young and aging MSCs as previously done by our group for rat bone marrow MSCs (Ali et al., 2015; Haneef et al., 2014), and we analyzed selective genes after 24 hours of re-oxygenation. This includes FGF, angiopoietin-1 (Ang-1), HGF, IGF, and TGFβ-1. When grown in a low-oxygen environment, MSCs stimulate the secretion of these growth factors (Crisostomo et al., 2008; Rochefort et al., 2006). Given their roles in beta-cell regeneration and in a variety of other cellular processes, it can be expected that these growth factors will contribute to the improved regeneration potential of preconditioned MSCs.
The FGF superfamily consists of structurally related polypeptides having diverse functions, including development, metabolism, wound healing, tissue repair, and angiogenesis. FGFs are involved in processes, including cell proliferation, migration, differentiation, adhesion, and survival (Laestander and Engström, 2014).
Ang-1 is a ligand for the Tie2 receptor, which may be involved in endothelial development (Davis et al., 1996). It causes a variety of effects such as inhibition of cell permeability and inflammation, activation of endothelial cell migration, vessel growth, and angiogenesis (Saharinen et al., 2010). Protective effects of Ang1/Tie2 signaling on cell apoptosis have been reported for many cell types, including endothelial cells, neurons, and cardiomyocytes (Kwak et al., 1999; Valable et al., 2003). It stimulates a number of intracellular signaling pathways, notably the PI3K/Akt pathway (Kontos et al., 1998), that play a significant role in the regulation of cell survival/apoptosis, differentiation, and proliferation (Fayard et al., 2005). The involvement of this pathway in the protection of MSCs against serum deprivation and hypoxia-induced apoptosis has been well established (Liu et al., 2008).
HGF has been identified in many tissues where it promotes angiogenesis and cell survival (Kitamura et al., 2007; Sulpice et al., 2009). It induces secretion of VEGF through activation of its receptor c-Met (Dong et al., 2001; Wojta et al., 1999). IGF are important mediators of cell growth, differentiation, and transformation (Delafontaine et al., 2004). HGF and IGF have been shown to prevent apoptosis and to stimulate the proliferation of pancreatic beta cells when secreted by MSCs (Dai et al., 2003).
TGFβ-1 is a member of a large family of cytokines that is involved in cell differentiation, proliferation, adhesion, and migration. Proteins of TGF-β family are expressed throughout embryogenesis (Heine et al., 1987). They are also involved in the process of angiogenesis by activation of serine/therionine kinase receptors (Chen et al., 1998). TGF-β is shown to be a vital trophic factor promoting islet survival, function, and regeneration (Han et al., 2011).
The ability of bone marrow-derived MSCs to secrete a number of cytokines and growth factors (Besse et al., 2000) decreases as the stem cells age (Khan et al., 2011). The loss of progenitor cell function with aging is, in part, mediated by an imbalance between factors that promote growth, migration, and survival, and factors enhancing telomere destruction and oxidative stress (Gonzalez et al., 2008). We have observed a decrease in certain growth factor expression in aging cells. This may result in a change in the differentiation potential of these aging stem cells along with many other associated properties.
To analyze the effect of hypoxia on the apoptosis of young and aging MSCs, their survival was measured by PI cellular uptake using flow cytometry. A slight increase in the number of apoptotic cells in aging MSCs was observed, but this change is non-significant. DNP seems to have no effect on both young and aging cells, which is expected as we used an optimized concentration of DNP and further re-oxygenation helped the cells to recover. Aging itself results in a decrease in the viability of bone marrow-derived MSCs; their therapeutic potential also declines with age (Shen et al., 2011; Yu et al., 2001). A number of cellular events that occur during the process of aging are responsible for these differences. When aging cells experience cellular insults, these cells undergo irreversible cellular changes due to their decreased potential to secrete cell survival factors.
We next investigated the role of hypoxic preconditioning in the pancreatic regeneration potential of young and aging MSCs. It has been documented that MSCs not only regenerate mesenchymal tissues, such as adipocytes, chondrocytes, and osteoblasts, but can also be trans-differentiated into other lineage-specific cells when they experience an appropriate microenvironment. This implies their efficacy in cell therapy applications (Wu et al., 2001). Our histological results showed that young and aging hypoxic MSCs as well as normoxic young MSCs showed significantly higher beta-cell regeneration potential compared with the normoxic aging MSCs after transplantation into diabetic mice. This is also evident by an increase in the blood plasma insulin concentration in animals that received DNP-treated cells. We also observed more insulin-positive cells in immunostained tissue sections of DNP-treated MSCs. Whether MSCs were able to directly differentiate into pancreatic beta cells or release paracrine factors to stimulate differentiation cannot be ascertained.
Transplantation of undifferentiated MSCs into diabetic mice supports the hypothesis that MSCs might differentiate toward insulin-producing β cells, or they might stimulate progenitor cell proliferation and differentiation. Intravenous administration of MSCs increased β-cell mass and decreased fasting blood glucose level in STZ-treated type-1 diabetic mice (Ezquer et al., 2008). However, whether MSCs can directly replace the loss of insulin-producing β cells is still uncertain. MSCs may not directly regenerate the pancreatic tissue by in vivo differentiation and migration, but they induce endogenous pancreatic tissue repair (Hess et al., 2003). It was also shown that new pancreatic β-cells are derived from the proliferation of pre-existing β cells (Choi et al., 2003; Dor et al., 2004). Emerging evidence now suggests that MSCs support islet function in an indirect manner by promoting the proliferation of pre-existing β-cells and angiogenesis, or in other words serving as a “trophic mediator.”
There are many other factors that need to be explored that can improve the differentiation potential of aging stem cells by adjustments in the levels of growth factors. By using various preconditioning strategies, such as DNP-induced chemical hypoxia, the cellular processes linked with differentiation of aging cells can be provoked, leading to enhanced differentiation potential of these cells. In conclusion, our results demonstrate that hypoxic preconditioning of the aging MSCs augments survival, migratory ability, and therapeutic potential in STZ-induced type-1 diabetic mice and, therefore, can result in better regeneration of pancreatic cells.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.