
Abstract
Abstract
Directly regulating the translation of POU5F1, SOX2, KLF4, and miRNA-145 plays an important role in maintaining the pluripotency of stem cells and the development of early embryos. In the present study, the expression model of miRNA-145 on bovine somatic cell nuclear transfer (SCNT) and in vitro fertilized (IVF) embryos were investigated and compared. Results indicated that (1) the expression level of miRNA-145 was significantly higher in SCNT embryos than that in IVF embryos after the eight-cell stage; (2) miRNA-145 negatively regulated the POU5F1, SOX2, and KLF4 in bovine embryos; (3) decreasing the expression of miRNA-145 by the miRNA-145 inhibitor significantly enhanced the expression of these three genes and the blastocyst formation rate; it also increased the total cell number and inner cell mass ratio of the bovine day 7 SCNT embryos. In conclusion, decreasing miRNA-145 expression might be a feasible means to enhance SCNT efficiency in bovines.
Introduction
S
Recent reports demonstrated that POU5F1 and SOX2 are two key genes related to the development of embryos (Daniels et al., 2000; Farthing et al., 2008; Keramari et al., 2010; Khan et al., 2012; Pfeiffer et al., 2010; Xu et al., 2013). Low expression of POU5F1 was shown to induce development failure in bovine embryos (Beyhan et al., 2007; Kirchhof et al., 2000), SOX2 was reported to be a cofactor of POU5F1 (Ariel et al., 2003), and KLF4, another member of the Yamanaka factors, was important in maintaining the pluripotency of embryos (Niemann et al., 2008; Vigneault et al., 2009).
miRNAs are a class of noncoding RNAs that are 18–22 nt in length and bind to target messenger RNAs (mRNAs) for posttranscriptional regulation of gene expression in both plants and animals (Garcia and Miska, 2005; Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and Ambros, 2001). By binding to partially complementary target sequences in the 3′ untranslated regions of mRNA, miRNAs negatively regulate the target expression by degrading the target mRNAs or repressing the encoded proteins (Bartel, 2004; Filipowicz et al., 2008; Lim et al., 2005). In general, one miRNA can regulate multiple target mRNAs, and one mRNA can be repressed by multiple miRNAs; thus, miRNAs compose a complex regulation network (Stefani and Slack, 2008). POU5F1, SOX2, and KLF4 are critical for embryo development (Farthing et al., 2008; Niemann et al., 2008), and a recent study proved that miRNA-145 directly targets these three genes in human embryonic stem cells (hESCs; Xu et al., 2009). This makes miRNA-145 a likely candidate regulator of embryo development.
In this study, we hypothesized that miRNA-145 could regulate the translation of POU5F1, SOX2, and KLF4. We studied three bovine embryo groups to investigate the effects of miRNA-145 on embryo development: in vitro fertilized (IVF) embryos (IVF group), untreated SCNT embryos (NT group), and miRNA-treated embryos (M-NT/I-NT group).
Materials and Methods
Ethics statement
The entire experimental procedure was approved by the Animal Care and Use Committee of Northwest A&F University. The experiment was performed in accordance with animal welfare and ethics guidelines. Bovine ovaries were obtained from the slaughterhouse in Xi'an (China) and transported to our laboratory in a thermos filled with sterile normal saline at 15–20°C (Lewis et al., 2003). For the nuclear donor cell culture, a newborn female Holstein cow was obtained from the Keyuan (Yangling, China) cloning company.
Chemicals
Unless otherwise stated, all chemicals and reagents were purchased from Sigma-Aldrich. Disposable, sterile plasticware was purchased from Corning.
Collection of oocytes and in vitro maturation
Bovine ovaries were collected from the local slaughterhouse, stripped of connective tissue, and then washed four times using sterile normal saline. A 10 mL injector was used to aspirate the cumulus oocyte complexes (COCs) from 2–8 mm antral follicles. All collected oocytes were checked under a stereomicroscope; only oocytes with rounded form, including the cytoplasm and more than three compacted cumulus cell layers, were selected. In vitro maturation (IVM) was conducted in a maturation medium (TCM 199; Gibco) containing 10% fetal bovine serum (FBS), 1 μg/mL 17 B-estradiol, and 0.075 IU/mL human menopausal gonadotropin. The cultures were incubated in 95% humidified air with 5% CO2 at 38.5°C for 20 hours.
Somatic cell nuclear transfer
The nuclear donor cells obtained from the ear skin of a newborn female Holstein cow were cultured as previously described (Wang et al., 2011). The donor cells were maintained in the Dulbecco's modified Eagle's medium (Gibco) containing 10% FBS (Gibco). The cells were cultured in a serum-starved medium containing 0.5% FBS for 2 days before their use as nuclear donors for SCNT. SCNT was conducted as described previously (Wang et al., 2011; Zhang et al., 2015). After IVM for 20 hours, matured COCs were blown and suctioned briefly with a 1 mL pipette (Eppendorf) for 1 minute. Then, they were moved into phosphate-buffered saline (PBS) supplemented with 0.1% bovine testicular hyaluronidase for 3 minutes to remove the rounded cumulus cells. Only the oocytes with a first polar body and a uniform granulated cytoplasm were selected for SCNT. The selected oocytes were moved into a droplet of PBS containing 7.5 μg/mL cytochalasin B (CB) and 10% FBS. Enucleation was performed using a 20 μm inner diameter glass pipette by aspirating the first polar body and a small amount of the surrounding cytoplasm.
A single-donor cell was injected into the perivitelline space of the enucleated oocyte as an oocyte cell complex. After all oocytes were treated with this procedure within 30 minutes, all oocyte cell complexes were moved into the fusion medium and washed three times. Thereafter, they were moved into a droplet of the fusion medium. Oocyte cell complex fusion was performed with a pair of platinum electrodes connected to a micromanipulator, and a double electrical pulse of 35 V for 10 μseconds was used to fuse the oocyte cell complex. The reconstructed SCNT embryos were cultured in a modified synthetic oviductal fluid (mSOF) for 2 hours until activation. All SCNT embryos were activated in 5 μM ionomycin for 5 minutes, followed by 4 hours of exposure to 1.9 mM 6-dimethylpyridine in mSOF. After activation, SCNT embryos were cultured with mSOFaa as conducted in a previous study (Wang et al., 2011).
SCNT embryos disposed with miRNA-145 mimic and miRNA-145 inhibitor
A miRNA-145 mimic and miRNA-145 inhibitor were designed and synthesized by Guangzhou RiboBio (RiboBio). As described, the oocyte cell complexes were moved into the fusion medium and washed three times. Then, they were moved into a droplet of the fusion medium containing 40 μM miRNA-145 mimic or miRNA-145 inhibitor. Oocyte cell complex fusion was performed as described. Reconstructed SCNT embryos were cultured in mSOF containing 40 μM miRNA-145 mimic or miRNA-145 inhibitor for 2 hours until activation. After activation, miRNA-145 mimic disposed SCNT embryos (M-NT) and miRNA-145 inhibitor disposed SCNT embryos (I-NT) were cultured with the same procedure as that of the SCNT embryos.
In vitro fertilization
Procedures and methods were described previously (Wang et al., 2011). Briefly, 200 μL of frozen–thawed semen was placed at the bottom of a 15 mL tube with 5 mL Brackett and Oliphant (BO) medium containing 6 mg/mL bovine serum albumin (BSA) and 20 mg/mL heparin. Cultures were incubated in 95% humidified air with 5% CO2 at 38.5°C for 30 minutes. After incubation, 4 mL supernatant BO medium was transferred to another tube and centrifuged at 1000 × g for 10 minutes. The liquid supernatant was discarded and 50 μL sperm suspension was left at the bottom of the tube. The sperm suspension was mixed and added into a BO medium droplet with 20–25 COCs for fertilization. After IVF for 20 hours, IVF oocytes were washed two times in an mSOF medium to remove the cumulus cells and redundant sperms. Then, they were cultured in a droplet with mSOFaa, as described in the SCNT procedure.
Immunofluorescence stain for analysis of inner cell mass and trophectoderm
According to the manufacturer's protocol, the blastocysts were washed three times with PBS containing 0.2% polyvinyl pyrrolidone (PVP) at room temperature and fixed in an immunology staining fix solution (Beyotime) for 1 hour. Then, the cell membrane was penetrated with 0.2% Triton X-100 in PBS for 30 minutes and blocked overnight in PBS with 2% BSA at 4°C. Thereafter, the cells were incubated with the mouse anti-Cdx2 monoclonal antibody with a dilution of 1:100 at 4°C for 12 hours. After being washed three times, the specific interaction between the primary antibody and the target was revealed by TRITC-labeled goat anti-mouse IgG (Beyotime) with dilution folds of 1:500. The cell nucleus was visualized by DAPI (Beyotime). After these procedures, the blastocysts were mounted on glass slides to analyze the fluorescence.
Quantitative real-time polymerase chain reaction
Embryos at different developmental stages (eight-cell, morula, and blastocyst) from the four groups (IVF, SCNT, M-NT, and I-NT) were selected for quantitative real-time polymerase chain reaction (qRT-PCR). All embryos were randomly chosen and pooled for total RNA extraction. For each replication, 10 eight-cell embryos, 5 morula embryos, and 5 blastocyst embryos from the same batch were randomly chosen. Total RNA was isolated with the Cells-to-Signal Kit (Invitrogen) according to the manufacturer's protocol. The complementary DNA (cDNA) was achieved and amplified with the cDNA synthesis kit (TaKaRa) according to the manufacturer's protocol. miRNA-145, POU5F1, KLF4, and SOX2 were quantified with SYBR Premix Ex Taq 2 (TaKaRa) on a StepOne real-time PCR system (StepOne and StepOnePlus; ABI) according to the manufacturer's protocol. The Histone 2A (H2A) and U6 small nuclear RNA were used as internal controls (Su et al., 2011; Wang B. et al., 2014) (Table 4).
F, forward; R, reverse. The reverse primer was 10×miScript Universal Primer provided in miScript II RT kit (Qiagen).
Statistical analysis
Experiments were repeated at least three times. Each repetition was performed using oocytes matured on the same day to eliminate batch effects. All embryos and oocytes were randomly chosen for each treatment group. Data were analyzed by one-way analysis of variance and least significant difference tests using SPSS 13.0 (SPSS, Inc.). Difference was considered significant at p < 0.05.
Results
The relative expression levels of POU5F1, SOX2, and KLF4 were significantly lower in the SCNT group than in the IVF group.
To explore the differences between bovine IVF embryos and SCNT embryos, the in vitro development rates from the two-cell embryo to the blastocyst of the two groups were calculated. We found that cleavage rates of the IVF and SCNT embryos were similar and continued to the four-cell stage; a significant difference was not shown until the eight-cell stage (Table 1). Thus, we focused on the differences at the eight-cell stage, morula stage, and blastocyst stage of the IVF and SCNT embryos. The relative expression levels of the four genes (miRNA-145, POU5F1, KLF4, and SOX2) among the three stages were measured by qRT-PCR (Fig. 1). Compared with the IVF embryos, the POU5F1, KLF4, and SOX2 transcripts were significantly lower in the SCNT group; meanwhile, the relative expression levels of miRNA-145 in the SCNT group were significantly higher than that in the IVF group.
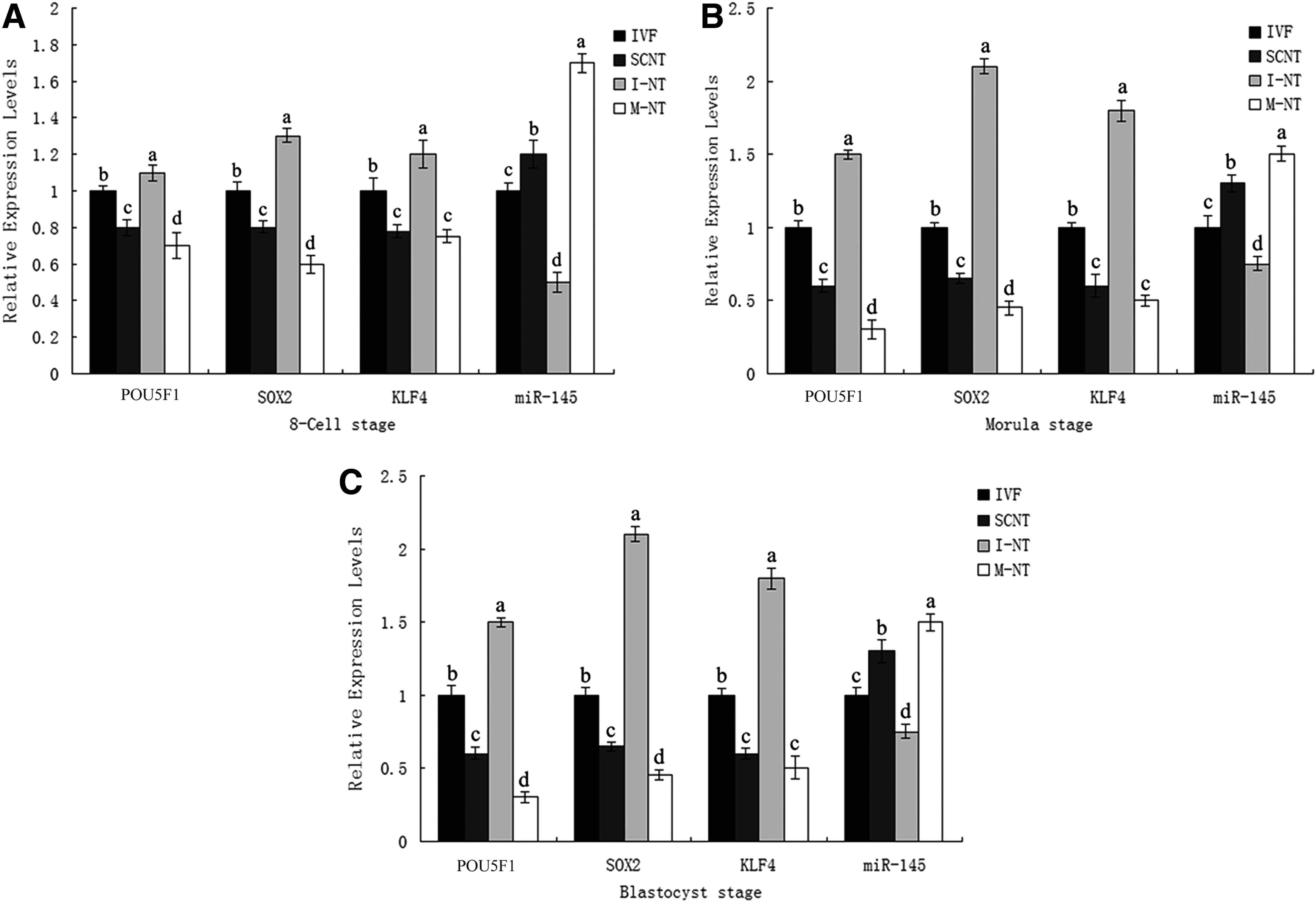
Relative expression levels of POU5F1, KLF4, SOX2, and miR-145 in different treated groups (IVF, SCNT, I-NT, M-NT) of bovine embryos. Values within one column graph that have different superscripts are significantly different from one another (p < 0.05).
Four replicates were performed in each treatment. 0 hour is the time the embryos were transferred to mSOFaa. Values within one column that have different superscripts are significantly different from one another (p < 0.05).
I-NT, miRNA-145 inhibitor-treated SCNT; IVF, in vitro fertilized; M-NT, miRNA-145 mimic-treated SCNT; SCNT, somatic cell nuclear transfer.
Effects of miRNA-145 on POU5F1, KLF4, and SOX2
To investigate the relationship between the miRNA-145 and the three genes (POU5F1, KLF4, and SOX2) in bovine embryos, SCNT embryos were separately disposed with the miRNA-145 mimic and inhibitor (Table 1). The miRNA-145 mimic disposed group had lower levels of expression of POU5F1, KLF4, and SOX2 than the SCNT group. By contrast, the miRNA-145 inhibitor disposed group had higher levels of expression of POU5F1, KLF4, and SOX2 than the SCNT group.
The miRNA-145 treatment significantly influenced the blastocyst rate of bovine SCNT embryos in vitro
To confirm the effect of miRNA-145 on the in vitro developmental capacity of bovine SCNT embryos, the SCNT embryos were separately disposed with the miRNA-145 mimic and inhibitor. In a preliminary test, the SCNT embryos disposed with 40 μM of miRNA-145 inhibitor significantly increased the blastocyst rate among other disposed groups (miRNA-145 inhibitor concentration at 0, 10, 20, 40, and 80 μM) (Table 2). Thus, we consider the 40 μM miRNA-145 inhibitor as the optimum for further research. For the corresponding treatment, SCNT embryos disposed with 40 μM of miRNA-145 mimic may also be used (Table 1).
Four replicates were performed in each treatment. 0 hour is the time embryos were transferred to mSOFaa. Values within one column that have different superscripts are significantly different from one another (p < 0.05).
Treatment with miRNA-145 affected the total cell numbers and inner cell mass ratio in blastocysts
To analyze the mechanism of the increased blastocyst rate by miRNA-145 inhibitor treatment, the total cell number and inner cell mass (ICM) ratio were measured via immunofluorescence assay. In the miRNA-145 inhibitor treatment group, the total cell number and the ICM ratio (ICM:trophectoderm [TE]) were significantly higher than those of the SCNT embryos. Conversely, in the miRNA-145 mimic treatment group, the total cell number and the ICM ratio in blastocysts were significantly lower than those of the SCNT embryos (Fig. 2 and Table 3).

Immunofluorescence of CDX2 and DAPI. Each blastocyst in the IVF, SCNT (NT), 40 μM of miR-145 inhibitor-treated SCNT (I-NT), and 40 μM of miR-145 mimic-treated SCNT (M-NT) groups was stained with CDX2 and DAPI.
No. of ICM cells + No. of TE cells = Total No. of cells.
The number of TE cell nuclei was counted using immunostaining for CDX2. The number of Total cells was counted using DAPI. The data are shown as mean ± SEM. Values within one column that have different superscripts are significantly different from one another (p < 0.05).
ICM, inner cell mass; SEM, standard error of the mean; TE, trophectoderm.
Discussion
For the bovine SCNT embryos, aberrant nuclear reprogramming is believed to be associated with the low efficiency of the development rate (Farin et al., 2006; Liu et al., 2015; Su et al., 2012; Yang et al., 2007). Recent studies showed that POU5F1, KLF4, and SOX2 were crucial genes during the reprogramming of bovine embryos. POU5F1 was reported to associate with maintaining the pluripotency of ICM in mouse embryos (Beyhan et al., 2007; Kirchhof et al., 2000). In bovine embryos, the expression of POU5F1 shows a similar pattern to its expression in mouse embryos, and lack of POU5F1 leads to development failure (Beyhan et al., 2007; Kirchhof et al., 2000). SOX2 was reported to cooperate with POU5F1 to establish the three cell lineages (Ariel et al., 2003; Huang et al., 2014). KLF4 exhibited the same function of maintaining the pluripotency of mouse and bovine embryos (Huang et al., 2014; Niemann et al., 2008; Vigneault et al., 2009). Several studies showed that the expression of POU5F1, KLF4, and SOX2 was lower in SCNT bovine embryos than in IVF bovine embryos (Huang et al., 2014).
In our study, we measured the expression differences of the three genes between the IVF and the SCNT bovine embryos based on the development stages. Results confirmed the previous conclusion that the expression of POU5F1, KLF4, and SOX2 was lower in the SCNT embryos than in the IVF embryos (Huang et al., 2014). This result indicated that the lower expression of these three genes in the SCNT embryos could be one reason for their lower blastocyst rate compared with that of the IVF embryos. In previous studies, the blastocyst rate of the SCNT embryos was improved with the increased expression of POU5F1 in bovines (Bortvin et al., 2003; Huang et al., 2014), and the blastocyst rate was increased with the enhanced expression levels of POU5F1, KLF4, and SOX2 in pigs (Huang et al., 2014). Those findings inspired us to further investigate increasing the efficiency of bovine SCNT embryos by modifying the expression of POU5F1, KLF4, and SOX2.
Previous studies reported that miRNA-145 directly targeted the POU5F1, KLF4, and SOX2 in hESCs (Xu et al., 2009). Upregulation with miRNA-145 would reduce the expression of these genes, and treatment of hESCs with a miRNA-145 inhibitor would improve their expression (Xu et al., 2009). We considered miRNA-145 as a candidate to modulate nuclear reprogramming in bovine embryos. Results showed that in bovine embryos, the expression of miRNA-145 was negatively related to the expression of POU5F1, KLF4, and SOX2. Furthermore, the miRNA-145 inhibitor could enhance the expression of POU5F1, KLF4, and SOX2, while the miRNA-145 mimic could reduce the expression of the three genes. Taken together, these results indicate that the miRNA-145 shows a similar regulation pattern on hESCs and bovine embryos.
Previous studies showed that the blastocyst rate of bovine SCNT embryos could be increased by improving the expression levels of POU5F1, KLF4, and SOX2 (Miyoshi et al., 2010; Su et al., 2011; Xu et al., 2012). In our study, when the expression of POU5F1, KLF4, and SOX2 by treatment was increased by the miRNA-145 inhibitor, the blastocyst rate of the bovine SCNT embryos was improved correspondingly. Meanwhile, treatment with the miRNA-145 mimic lowered the expression of POU5F1, KLF4, and SOX2 and reduced the blastocyst rate of the bovine SCNT embryos. This result confirmed former reports that the expression levels of POU5F1, KLF4, and SOX2 were vital to the blastocyst rate of bovine SCNT embryos (Beyhan et al., 2007; Kirchhof et al., 2000; Niemann et al., 2008; Vigneault et al., 2009).
In previous studies, POU5F1 was associated with maintaining the pluripotency of ICM (Beyhan et al., 2007; Kirchhof et al., 2000). Increased expression of POU5F1 would improve the ICM ratio of the bovine SCNT embryos (Miyoshi et al., 2010; Su et al., 2011; Xu et al., 2012). SOX2 cooperated with POU5F1 (Ariel et al., 2003; Huang et al., 2014), and KLF4 was crucial in maintaining the pluripotency of embryos (Huang et al., 2014; Niemann et al., 2008; Vigneault et al., 2009). The bovine SCNT embryos treated with Oxamflatin increased the total cell number and ICM ratio. Meanwhile, the expression levels of POU5F1 and SOX2 increased as well (Su et al., 2011). In pigs, the SCNT embryos treated with scriptaid showed an improved blastocyst rate with an increased expression of POU5F1 and KLF4 (Zhou et al., 2013). Previous studies indicated that the expression levels of POU5F1, KLF4, and SOX2 corresponded to the quality of embryo blastocysts and the ICM ratio. Those indications are also consistent with our observations.
In conclusion, our study demonstrated that the expression levels of POU5F1, KLF4, and SOX2 in bovines were critical to the nuclear reprogramming of embryos. Furthermore, the blastocyst rate of the SCNT embryos could be improved by enhancing the expression of these genes. Moreover, miRNA-145 was identified as a negative regulator for POU5F1, KLF4, and SOX2 in bovine embryos, and the bovine SCNT embryos treated with an miRNA-145 inhibitor expressed POU5F1, KLF4, and SOX2 more strongly. Moreover, treatment with 40 μM of the miRNA-145 inhibitor significantly improved the blastocyst rate of bovine SCNT embryos in vitro and increased the ICM ratio of the blastocyst embryos.
Footnotes
Acknowledgments
This study was supported by the National High Technology Research and Development Program of China (863 Program, 2011AA100303). The authors thank Younan Wang for transportation of the Holstein cow ovaries used in this study. They are also grateful to Dr. Hui Zhang for the technical assistance as well as to the members of the research group for technical support and helpful discussion.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.