
Abstract
Abstract
Cell Stemness can be achieved by various reprogramming techniques namely, somatic cell nuclear transfer, cell fusion, cell extracts, and introduction of transcription factors from which induced pluripotent stem cells (iPSCs) are obtained. iPSCs are valuable cell sources for drug screening and human disease modeling. Alternatives to virus-based introduction of transcription factors include application of DNA-free methods and introduction of chemically defined culturing conditions. However, the possibility of tumor development is still a hurdle. By taking advantage of NTERA-2 cells, a human embryonal carcinoma cell line, we obtained partially differentiated cells and examined the dedifferentiation capacity of regenerative tissue from rabbit ears. Results indicated that treatment of partially differentiated NTERA-2 cells with the regenerating tissue-conditioned medium (CM) induced expression of key pluripotency markers as examined by real-time polymerase chain reaction, flow cytometry, and immunocytochemistry techniques. In this study, it is reported for the first time that the CM obtained from rabbit regenerating tissue contains dedifferentiation factors, taking cells back to the pluripotency. This system could be a simple and efficient way to reprogram the differentiated cells and generate iPSCs for clinical applications as this system is not accompanied by any viral vector, and reprograms the cells within 10 days of treatment. The results may convince the genomic experts to study the unknown signaling pathways involved in the dedifferentiation by regenerating tissue-CM to authenticate the reprogramming model.
Introduction
U
In 2006, a giant step was taken to impose direct reprogramming of somatic cells by transducing mouse fibroblasts with four exogenous transcription factors, Oct4 (O), Sox2 (S), Klf4 (K), and c-Myc (M). The resulting reprogrammed cells are referred as induced pluripotent stem cells (iPSCs) (Takahashi and Yamanaka, 2006). Despite the significant technical values of this approach, the iPSC technology suffers from a couple of drawbacks, for example, the recruitment of exogenous genetic factors (viral based constructs) for delivering the reprogramming factors in current available strategies. This brings the risk of cancer development, attributed to retroviral activation or insertional mutagenesis and also the presence of c-Myc and Klf4 genes in the reprogramming cocktail. Moreover, iPSC technology is accompanied with low efficiency and slow kinetics of reprogramming, which takes at least 2 to 3 weeks for emerging first reprogrammed cells in conventional protocols (Aasen et al., 2008; Aoi et al., 2008; Maherali et al., 2007; Okita et al., 2007, 2008; Okita et al., 2013; Park et al., 2008; Stadtfeld et al., 2008; Takahashi and Yamanaka, 2006; Takahashi et al., 2007; Wernig et al., 2007; Yu et al., 2007). Altogether, at the moment, these issues serve as barriers to applicability of this strategy in clinical practice.
To address these concerns, alternative methods have been applied for delivering reprogramming factors. It was demonstrated that iPSCs can be generated through nonintegrating or excisable gene delivery methods; although these approaches are safer than older risky ones, the risk of using genetic factors is still remaining (Okita et al., 2008, 2011; Somers et al., 2010; Stadtfeld et al., 2008; Woltjen et al., 2009; Zhou and Freed, 2009). Therefore, iPSCs have been derived by several DNA-free methods: direct delivery of key reprogramming proteins (Kim et al., 2009; Zhou et al., 2009), modified mRNAs (Warren et al., 2010), OSKM transgene delivery using Sendai virus (an RNA virus), and mature microRNA transfection (miR-200s, miR302s, and miR369s) (Miyoshi et al., 2011). Despite noticeable advantages, these methods are accompanied with either low efficiency of reprogramming (Kim et al., 2009; Miyoshi et al., 2011; Zhou et al., 2009) or technical challenges (Fusaki et al., 2009; Warren et al., 2010).
Among a variety of reprogramming approaches, induction of pluripotency, using defined chemical conditions, has gained ample attention for its potency in therapeutic applications. Hence, there are increasing efforts to identify chemical components with the power of somatic cell reprogramming or/and the ability to improve the efficiency of this process. As a result, several chemicals have been reported to either increase reprogramming efficiency or substitute for reprogramming factors (Huangfu et al., 2008; Ichida et al., 2009; Li et al., 2011; Lin et al., 2009; Marson et al., 2008; Shi et al., 2008; Silva et al., 2008; Zhu et al., 2010) (for a review, see [Nakhaei-Rad et al., 2012]).
Therefore, small-molecule screening studies should continue to identify crucial chemical components in derivation of iPSCs. In vitro studies have demonstrated the capacity of newt regenerating tissue extracts (McGann et al., 2001) and ES cell-conditioned medium (CM) (Grinnell and Bickenbach, 2007) for inducing dedifferentiation in somatic cells. Among mammals, scarless wound healing in rabbit ears seems to be a perfect pattern for tissue regeneration (Goss and Grimes, 1972, 1975; Grimes and Goss, 1970). Instead of using chemical components with the potential of reprogramming induction, we made an effort to choose natural components by employing the rabbit regenerating tissue-CM, which is supposed to initiate the regeneration mechanisms. In this simplified biological model, NTERA-2 (NT2) cells were first partially differentiated and then utilized for dedifferentiation screening purpose. We clarified that rabbit regenerating tissue-CM contains the factors that are able to turn on dedifferentiation program in this partially differentiated human cell line.
Materials and Methods
Cell culture before treatments
NTERA-2 cl. D1 cells (a generous gift from prof. Andrews from Sheffield University) were cultured in Dulbecco's Modified Eagle's Medium (DMEM, Gibco), supplemented with 10% fetal bovine serum (FBS, Gibco) at 37°C and 10% CO2. For prevention of differentiation, NT2 cells were passaged with a scraping approach using glass beads.
Preparation of conditioned medium from rabbit regenerating tissue
Two- to four-month-old male New Zealand white rabbits (Oryctolagus cuniculus) were purchased from Razi Vaccine and Serum Research Institute, Mashhad, Iran. Regeneration in rabbit ears was induced through making holes in their pinnas. To do so, rabbit ears were shaved and then anesthetized by lidocaine spray. Three holes were created in each pinna with the aid of a puncher, each 2.5 mm in diameter. Two days later, the regenerating tissues were removed as a ring-shaped tissue (by a second punch, each 4.5 mm in diameter), in the margins of the previous holes. The regenerating tissues were kept for 48 hours in DMEM, supplemented with 10% FBS and incubated at 37°C and 10% CO2. The medium was collected after 48 hours and passed through a 0.22-μm sterile filter. This medium is referred as rabbit regenerating tissue-CM in this work. As a control for nonregenerative tissue, rabbit nonregenerative tissue-CM was prepared from the back skin of rabbit, following the same procedure. The animals were treated and maintained according to the standard guidelines of Animal Care and Use Committee at Ferdowsi University of Mashhad.
Treatment of NT2 cells with retinoic acid and CM
Cell differentiation was induced by treatment of NT2 cells with a medium containing 10−5 M all-trans retinoic acid (RA) (Eastman-Kodak, Rochester, NY) for 72 hours (Andrews, 1984). NT2 cells were seeded and assayed in three different groups, naming untreated, NT2-RA, and NT2-RA-CM-treated cells. The first group of the cells was fed with DMEM-FBS, and the second and third groups were initially fed with DMEM-FBS, containing 10−5 M RA for 72 hours. After this period, cells were passaged and their media were changed by fresh DMEM-FBS and DMEM-FBS containing 1500 μL (75%) of the CM in the second and third groups, respectively, and incubated for another 3 days in the same conditions. In second passage, the media of all groups were replaced by the conventional medium of DMEM-FBS and incubated for three more days.
Semiquantitative RT-PCR and real-time polymerase chain reaction analyses
Total RNA was extracted using RNX-Plus solution (CinnaGen). RNA samples were verified by electrophoresis on a 1.2% agarose gel. RNA concentrations were quantified by a Nanodrop spectrophotometer. To exclude any genomic DNA contamination, RNA samples were treated by DNaseI enzyme following the manufacturer's protocol (Ambion). cDNA was then synthesized as follows: 1 μL of Oligo-dT primer (10 pmol) was added to 2 μg of total RNA and nuclease-free water (up to total volume of 12 μL). Next, the mixture was heated at 70°C for 10 minutes and kept on ice for 5 minutes. Then, the following components were added: 1 μL of M-MuLV Reverse Transcriptase (200 U/μL; Fermentas), 4 μL of 5X RT buffer, 1 μL of RiboLock RNase inhibitor (40 U/μL; Fermentas), and 2 μL of dNTPs mix (10 mM;, CinnaGen) to a final volume of 20 μL. Reaction mixture was incubated for 1 hour at 42°C and then 72°C for 10 minutes. For polymerase chain reaction (PCR), 0.25 μL cDNA was used as a template. 2X SYBR Green Master Mix (Pars Tous) was applied for relative real-time PCRs. Primer sequences for RT-PCR and real-time PCR are listed in Tables 1 and 2.
Flow cytometry
Flow cytometry analysis was performed as previously described (Mahmoudi et al., 2011). Briefly, cells were incubated for 45 minutes at 4°C with primary antibodies: anti-human Oct4 (1:200) (SC-5279; Santa cruz), anti-human SSEA-3 (MC631) (1:50), SSEA-4 (MC813-70) (1:50), TRA-1-60 (1:50), and TRA1-81 (1:50) (these antibodies were generous gifts from prof. P.W. Andrews, University of Sheffield). Following washing thrice in a wash buffer (phosphate-buffered saline [PBS] containing 5% FBS), cells were incubated for 30 minutes at 4°C with the secondary antibody, FITC-conjugated goat anti-mouse IgG (1:100), and we applied the incubated cells with secondary antibody alone as a negative control.
Immunofluorescence analysis
The control and treated cells were seeded on sterile cover slips and cultivated to reach 70%–80% confluency. They were then washed with the wash buffer (PBS containing 5% FBS), fixed with 4% paraformaldehyde/PBS for 20 minutes at 4°C, and incubated for 10 minutes in 0.2% digitonin (Sigma) for permeabilization. Cell staining was performed following 45 minutes incubation in the primary antibodies at room temperature: anti-human Oct4 (1:200), anti-human SSEA-3 (MC631) (1:50), SSEA-4 (MC813-70) (1:50), TRA-1-60 (1:50), and TRA-1-81 (1:50). The slides were then washed thrice and incubated with FITC-conjugated goat anti-mouse IgG (1:100). After the final washing, they were counter stained with DAPI (Invitrogen) to identify nuclei. Incubated cells with the secondary antibodies alone served as negative controls.
Results
NT2 cells are a cloned human embryonal carcinoma cell line, which are pluripotent cells and can be easily differentiated in response to RA treatment (Andrews, 1984; Andrews et al., 1984, 2005). Similar to ES cells, NT2 cells express the key pluripotency factors, including OCT4, SOX2, and NANOG (Sperger et al., 2003), and SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 cell surface antigens (Andrews et al., 1996; Matin et al., 2004; Niwa et al., 2000; Sperger et al., 2003). During differentiation process, the expression of these factors is affected and the modest decrease is obvious. To determine dedifferentiation ability of rabbit regenerating tissue-CM in differentiated cells, first we generated partially differentiated NT2 cells through treatment of these cells with 10−5M RA and then we examined the effects of the CM in partially differentiated NT2 cells, as illustrated in Figure 1.
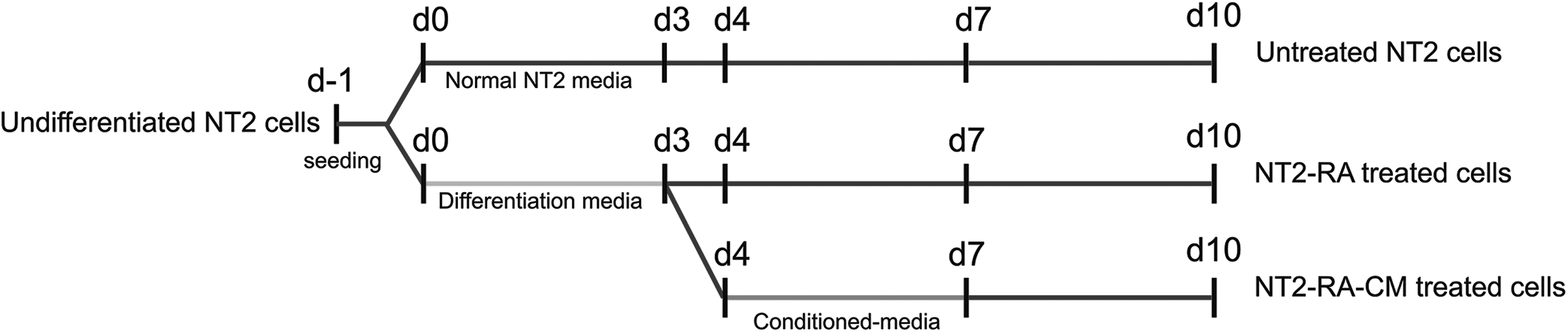
Schematic representations of the current protocol.
Differentiation/dedifferentiation of the treated NT2 cells
It has been shown that differentiation of NT2 cells by RA treatment can be evident by upregulation of SSEA-1 and downregulation of cell surface antigens correlated to pluripotency, including SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 (Draper et al., 2002; Matin et al., 2004).
After treatment of the NT2 cells with the differentiation medium and regenerating tissue-conditioned medium, they were analyzed for changes in the expression of cell surface markers by flow cytometry. Untreated NT2 cells showed high levels of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 markers along with low expression of SSEA-1 (Figs. 2 and 3). Reduced expression of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 markers, and increased expression of SSEA-1 were observed, in cells under RA treatment. In contrast, NT2-RA-CM-treated cells presented the opposite patterns of antigen presentation, that is, high levels of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 were detected in these cells, whereas SSEA-1 expression was decreased.
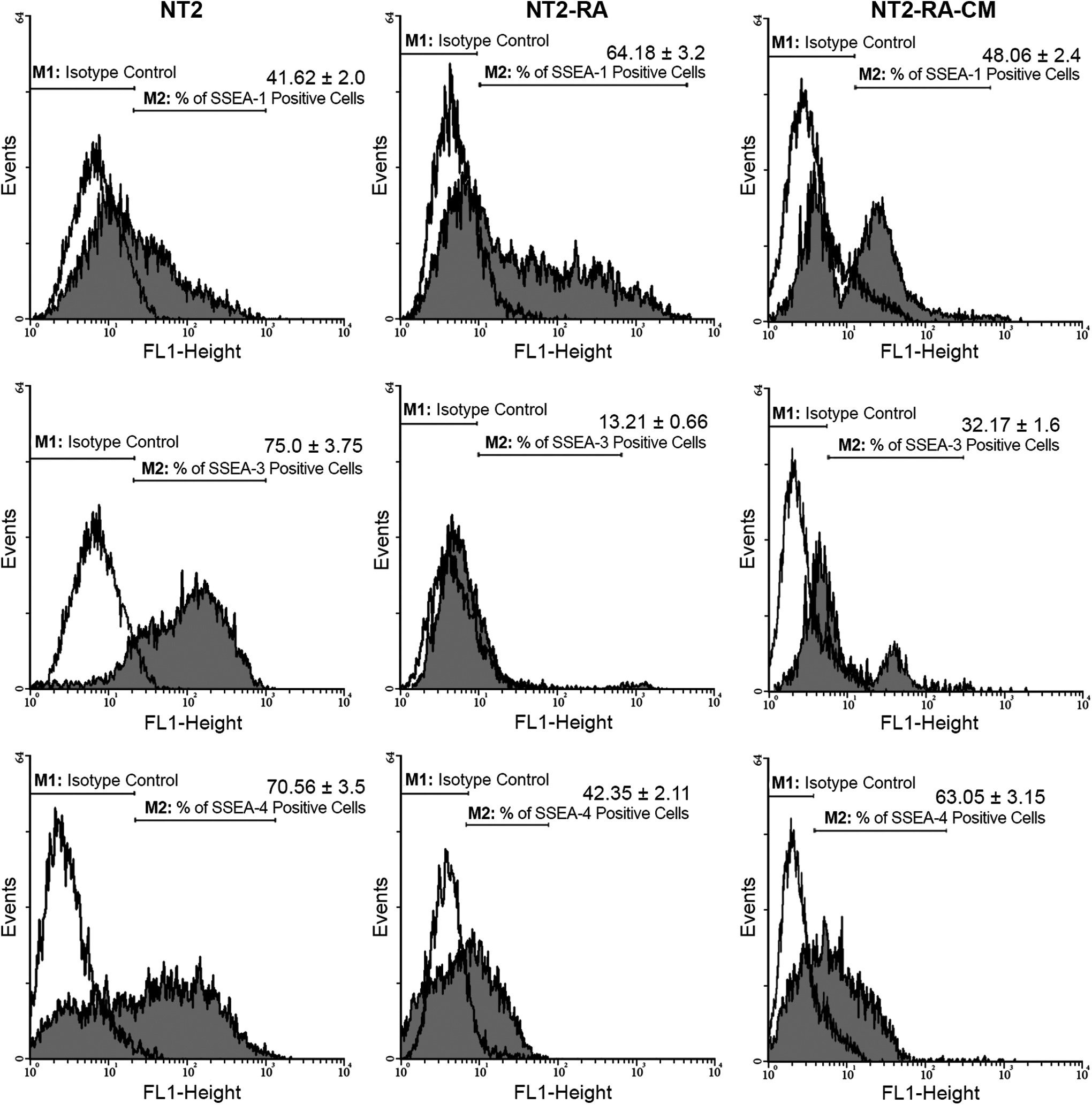
Histogram analyses of cell surface antigen markers of SSEA-1, SSEA-3, and SSEA-4 in control (untreated) and treated (NT2-RA and NT2-RA-CM) cells. The values on top of each bar represent the percentages of the positive cells for the related cell surface antigen marker.
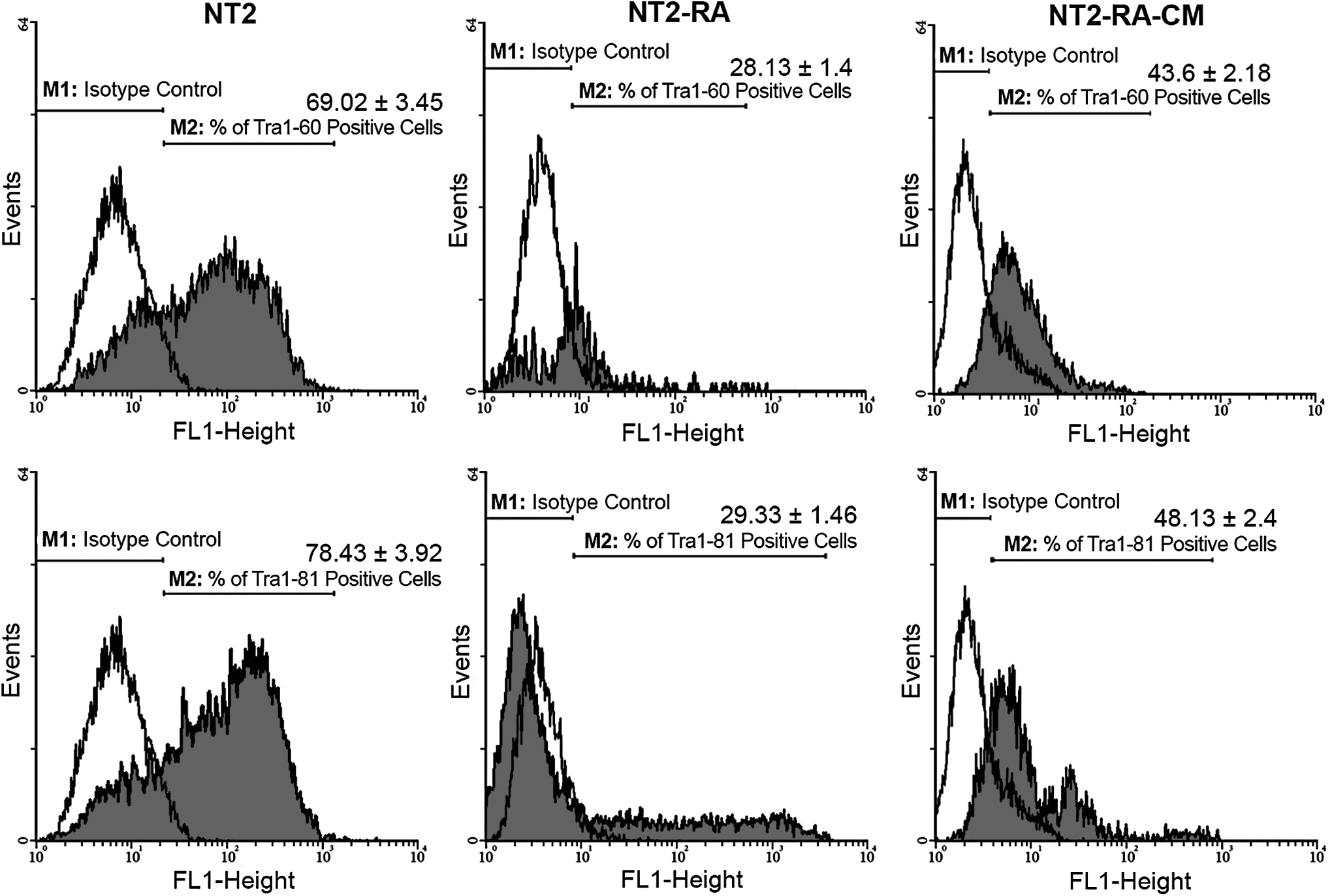
Histogram analyses, which illustrate and compare the expression levels of pluripotency markers: TRA-1-60 and TRA-1-81 in control (untreated) and treated (NT2-RA and NT2-RA-CM) cells. The values on top of each bar represent the percentages of the positive cells for the related cell surface antigen marker.
These changes were also verified by immunocytochemistry analysis in an independent experiment. Overall, as expected, the majority of untreated NT2 cells expressed undifferentiation markers, including SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 4A, Supplementary Figs. S1A–3A; Supplementary Data are available online at www.liebertpub.com/cell), but not SSEA-1 (Fig. 5A). Conversely, in response to RA-induced differentiation, NT2 cells lost the expression of the undifferentiation markers (Fig. 4B, Supplementary Figs. S1B–3B), whereas SSEA-1 expression was detectable in most of the cells (Fig. 5B). Following CM treatment, NT2-RA-CM-treated cells restored the expression of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 4C, Supplementary Figs. S1C–3C), and in contrast, most of the cells mainly became negative for SSEA-1 expression (Fig. 5C). In summary, it was demonstrated that rabbit regenerating tissue-CM induces the expression of pluripotency markers in partially differentiated cells to a level that is similar to untreated NT2 cells.
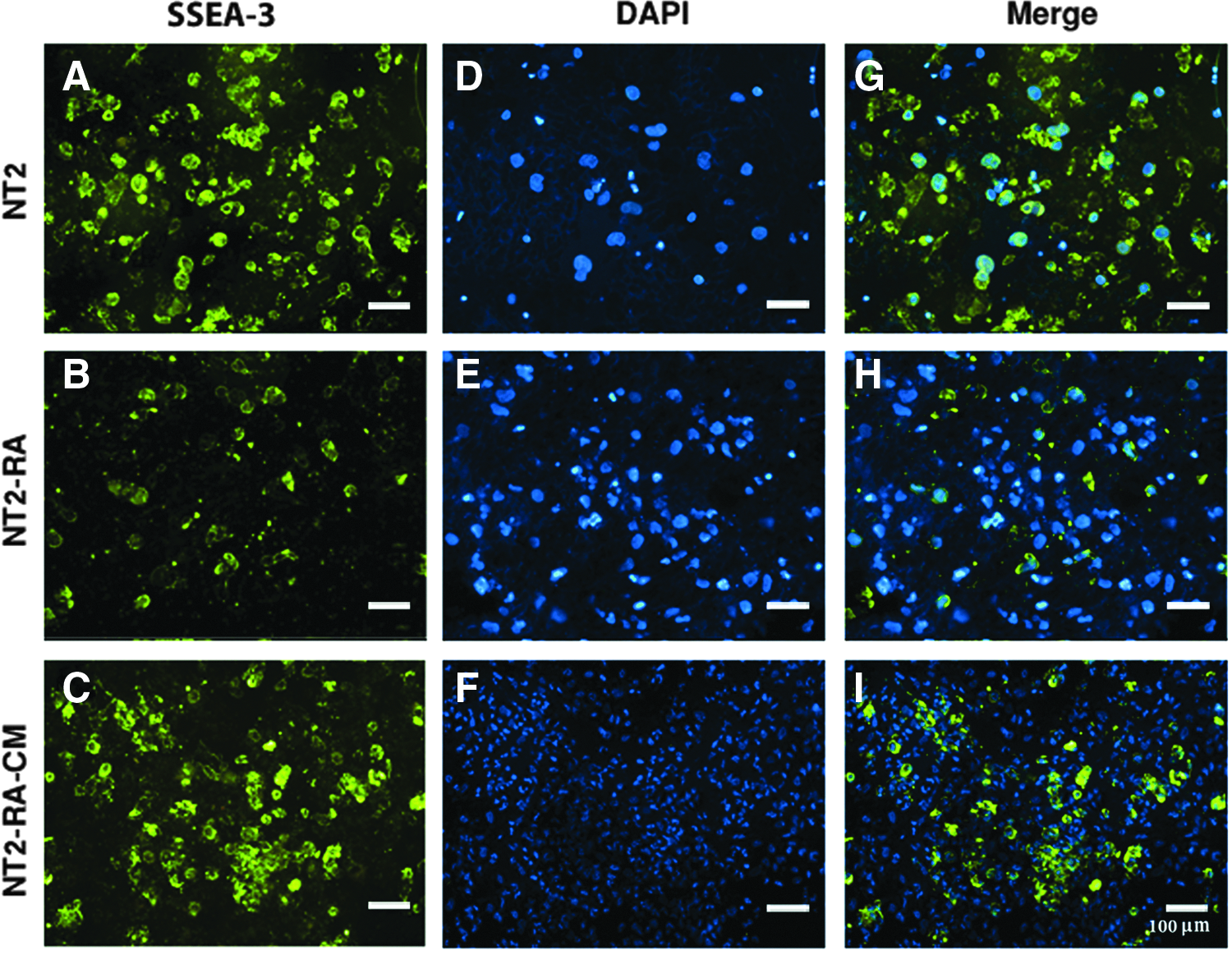
CM from the regenerating rabbit ear restores the undifferentiation marker of SSEA-3 in differentiated NT2 cells. Immunocytochemistry experiments indicate:
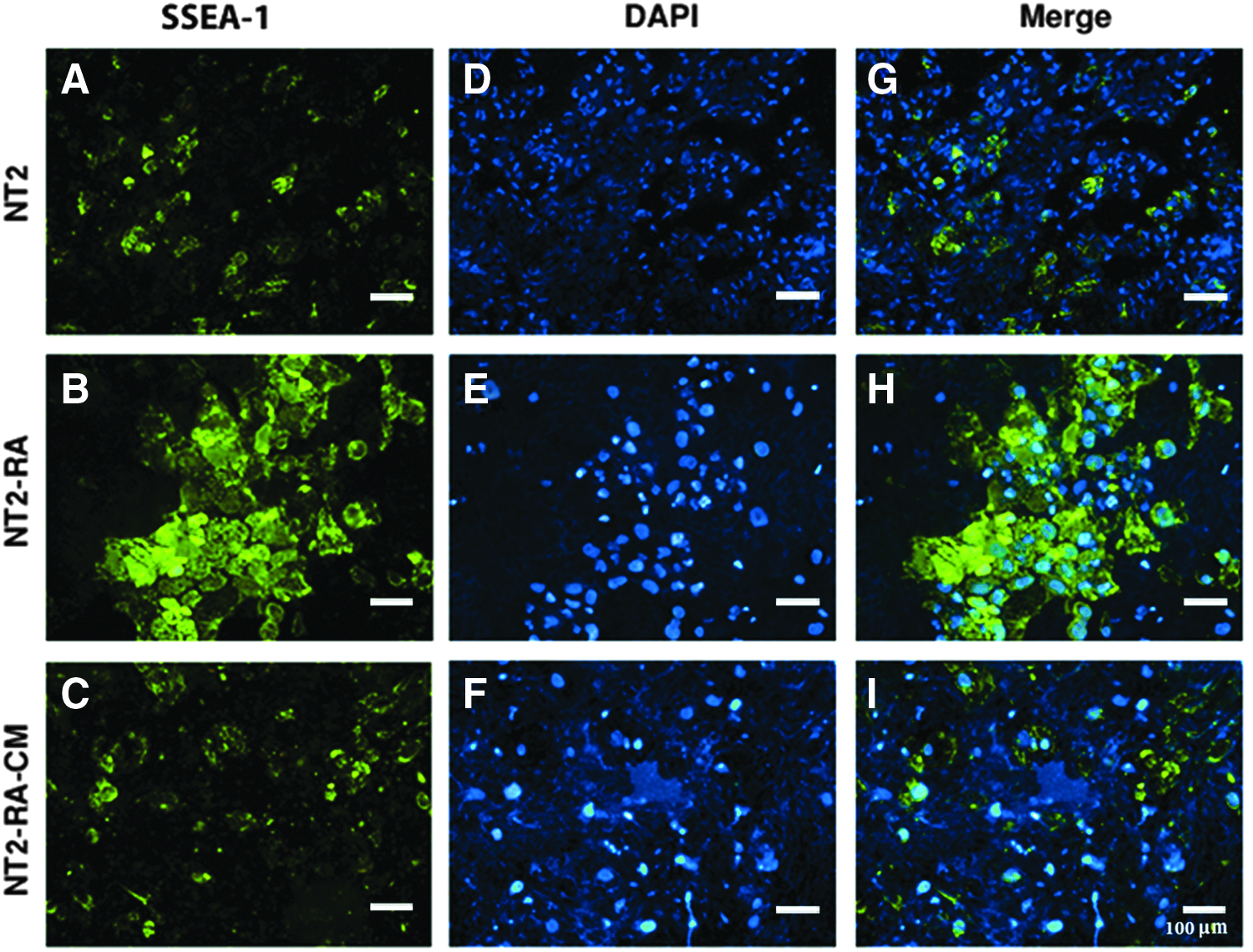
The rabbit regenerating tissue-CM downregulates the expression of differentiation indicator in differentiated NT2 cells. Immunostaining results for SSEA-1 expression show the following:
OCT4 expression changes in response to RA and RA-CM treatment of NT2 cells
To investigate the effects of CM on the regulation of key reprogramming factors, the expression of these factors were monitored at the mRNA level with the conventional RT-PCR method. As expected, before induction of differentiation, the untreated NT2 cells were expressing the pluripotency markers, including OCT4, SOX2, and NANOG. These levels dropped significantly upon treatment with RA in all cases. Treatment of the NT2-RA cells with CM restored the expression of OCT4 gene, without an obvious impact on the level of SOX2 and NANOG transcripts (Fig. 6A).
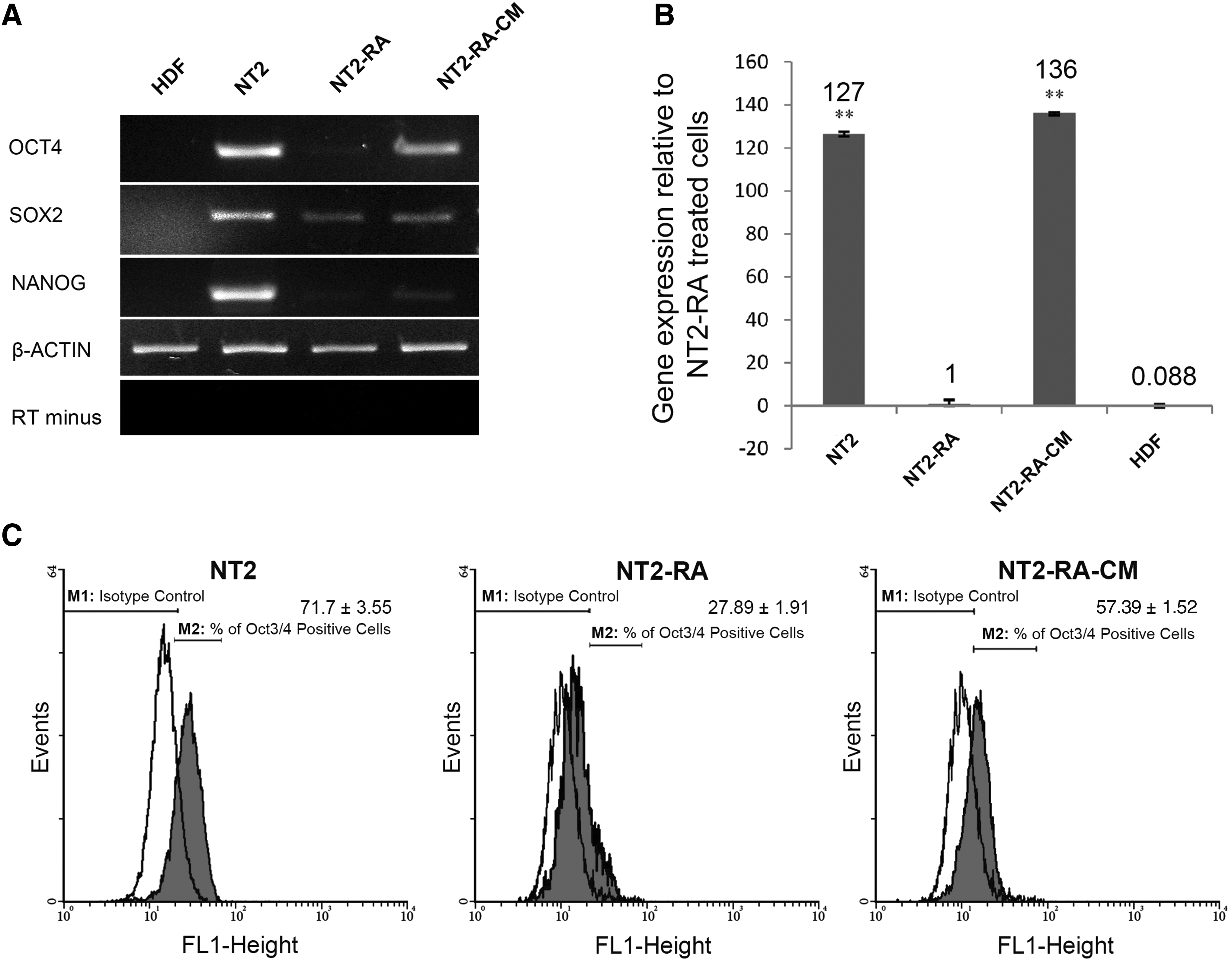
OCT4 expression is restored in the differentiated NT2 cells after the CM treatment. Semiquantitative analysis indicates the following:
To confirm the changes of OCT4 expression both at transcript and protein levels, more precise assessments were performed by relative real-time PCR and indirect flow cytometry analyses, respectively (Fig. 6B, C). The real-time PCR results detected an increase in OCT4 expression following CM treatment of NT2-RA-treated cells. Interestingly, in NT2-RA-CM-treated cells, the OCT4 expression was higher than untreated NT2 cells. In addition, we applied the CM from a nonregenerating tissue (rabbit skin) as a negative control, and it could not induce OCT4 mRNA expression in NT2-RA-treated cells (Supplementary Fig. S4). Furthermore, flow cytometry analysis showed that in addition to mRNA level, CM treatment positively affected the OCT4 protein level. Altogether, these results indicate that rabbit regenerating tissue-CM is able to restore the expression of OCT4 in NT2-RA-treated cells (NT2-RA-CM).
Discussion
Among vertebrates, mammals show limited regeneration capacity in comparison with urodele amphibians (Carlson, 2005). This is considered an unfortunate limitation in humans because of its adverse impacts on recovery of degenerative diseases. Having access to cells with stemness characteristics, for example, differentiation into different cell lines is sought by different strategies to overcome this limitation. The somatic cell reprogramming toward a pluripotent state is believed to provide useful cell sources for applications in cell therapy if the main concerns regarding their side effects are addressed properly.
To translate this capacity to regenerative medicine, intensive efforts have been dedicated in the last two decades. Somatic cell nuclear transfer (Wilmut et al., 1997), fusion of somatic cells with ES cells (Do and Scholer, 2004), administration of stem cell extracts in the somatic cell culture media (Freberg et al., 2007; Hansis et al., 2004; Taranger et al., 2005), and most recently, induction of reprogramming by forced expression of key transcription factors (iPSCs) (Takahashi and Yamanaka, 2006) have served as successful approaches for induction of stemness in somatic cells. Development of safe, feasible, and ethical protocols has been a milestone for most of these approaches.
Wound healing has been shown to be a quick and exceptionally perfect process in rabbit ear (Goss and Grimes, 1972, 1975). The question is whether this happens by recruitment of the circulating stem cells or induction of the local cells to dedifferentiate and play as supplying stem cells. The later scenario, which is referred as blastema formation has been suggested in amphibians (Thornton, 1957, 1968; Wallace, 1981). This study sought the possible mechanism for regeneration of wounded ears in rabbit by introduction of a simple, fast, and safe model for dedifferentiation screening. We took advantage of NT2 pluripotent cells in responding to RA by downregulation of certain stemness-associated molecular markers and consequent commitment to differentiation (Bahrami et al., 2005).
There is a report of using CM from mouse ES cells to induce pluripotency-related genes in human keratinocytes (Grinnell and Bickenbach, 2007). We have previously shown that blastema-like cells are formed in the regenerating wounds of rabbit ears (Mahmoudi et al., 2011). These cells were isolated and shown to express the pluripotency genes of Oct4 and Sox2. In this study, CM from the regenerating rings of the ears were administered for reprogramming of partially differentiated NT2-RA-treated cells and showed that this CM could significantly increase the OCT4 gene expression. This effect was apparently translated to dedifferentiation of the cells by recovering the expression of SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81, and repressing the expression of SSEA-1 cell surface antigens. Pluripotency and differentiation markers used in this study have been compared in a single table (Table 3), showing their expression pattern during different treatments.
+, positive expression or overexpression compared to control; −, no expression or reduced expression compared to control.
Cellular and molecular mechanisms underlying regeneration and formation of blastema-like cells in the rabbit ear remain poorly understood. Our data suggest that rabbit regenerating tissue, like oocyte cytoplasm, contains the appropriate dedifferentiation initiation factors. This combination of soluble factors, which are released during the regeneration of rabbit ear, needs to be addressed. We categorized these possible molecules in three groups: (1) Common signaling molecules with critical roles in self-renewal and survival of stem cells, including BMP-4 (Grinnell and Bickenbach, 2007), bFGF (Xie et al., 2008), IGFI, IGFII (Bos et al., 2001) Shh, and Wnt3a (Singh et al., 2012). (2) Inhibitors of cell differentiation program; previous studies have confirmed the influence of such inhibitors in the improvement of somatic cell reprogramming, these factors include MEK/ERK, GSK3 (Silva et al., 2008), RasGAP (Chen et al., 2006), and ALK5 inhibitors (Maherali and Hochedlinger, 2009). (3) The epigenetic modifiers that act directly at the gene level to reactivate epigenetically silenced stemness genes or repress the expression of lineage-specific genes.
In this regard, it would be useful to apply different agonists and antagonists for the candidate pathways to clarify the key signaling mechanism(s) involved in cellular dedifferentiation. Apart from this broad range of candidate factors, which should be investigated in details, there is a possibility to find a new combination of molecules that coordinate in the regeneration process and cellular dedifferentiation. Therefore, the next aim would be the performance of proteomics analyses between regenerating tissue-CM and nonregenerating-CM to explore the new components in the regeneration medium. Moreover, the expression profiling of rabbit regenerative tissue can determine the factors that are expressed differently during regeneration. Collectively, these approaches can shed light to improve our knowledge about mechanisms governing regeneration and dedifferentiation in rabbit ear.
In summary, we reconstructed a conventional dedifferentiation protocol and introduced a novel reprogramming cocktail. Moreover, a simple biological model is introduced for screening the reprogramming of small molecules. Based on this model, our results provide molecular evidence to add some weight to the notion that regenerative wound healing in mammals is a result of cellular reprogramming and dedifferentiation. Finally, it seems that the wounded ears of rabbits include signals/components that not only induce local blastema formation but also could be functional for induction of reprogramming in other differentiated cell lines. Although this property looks promising for promotion of iPSC technology, the nature of signals responsible for it remains to be elucidated.
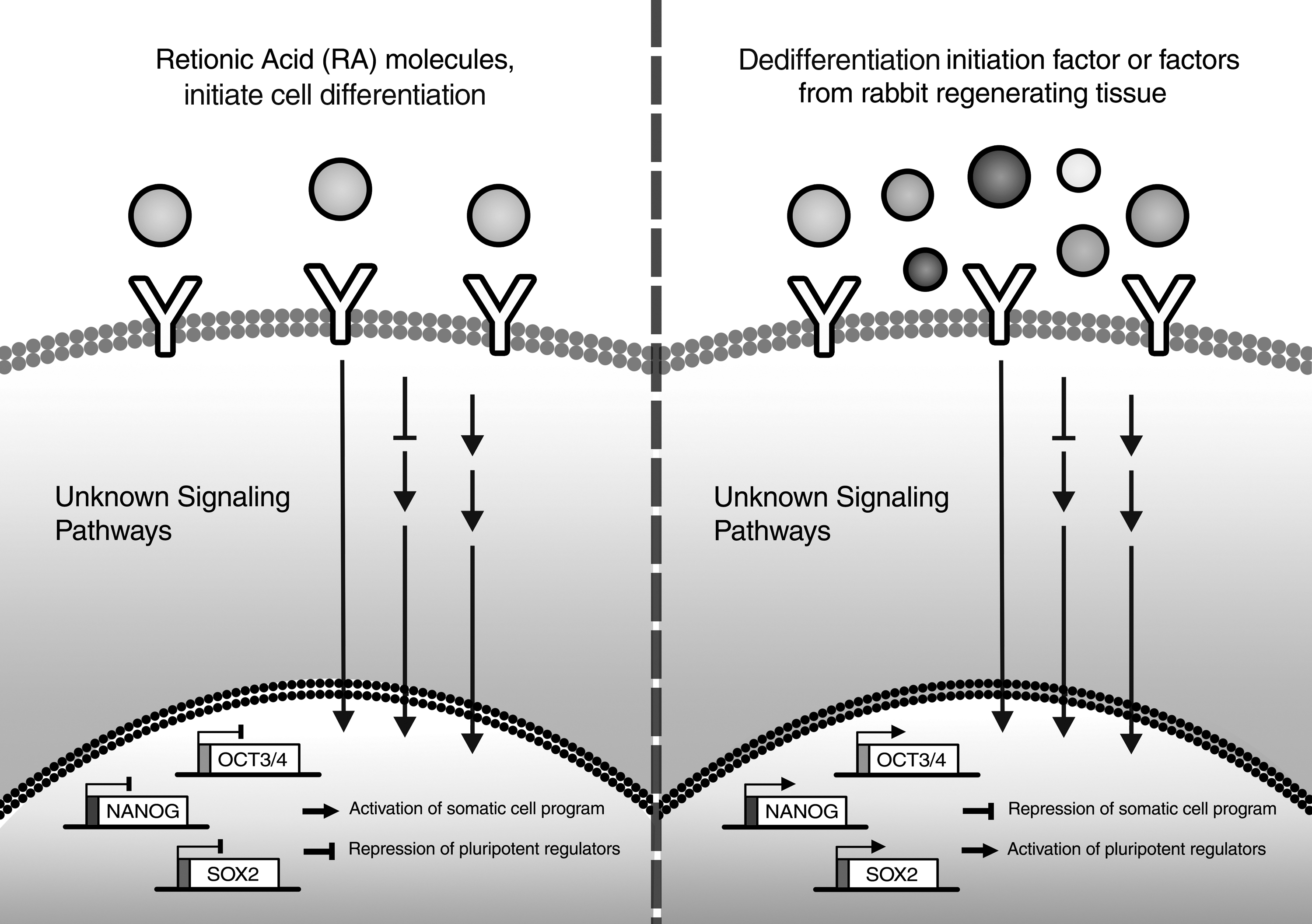
Proposed model of this study.
Footnotes
Acknowledgments
Authors would like to thank Mr. Malaekeh and Ms. Tabasi for providing technical assistance in flow cytometry analyses, and Mr. Farshchian and Dr. Haddad-Mashadrizeh for their assistance in gene expression profiling. This work was supported by a grant (No. 871339052) from Institute of Biotechnology, Ferdowsi University of Mashhad, Mashhad, Iran.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.