
Abstract
Abstract
Oocytes rapidly lose their developmental potential after ovulation, termed postovulatory oocyte aging, and often exhibit characteristic phenotypes, such as cytofragmentation, abnormal spindle shapes, and chromosome misalignments. Here, we reconstructed mouse oocytes using somatic cell nuclear transfer (SCNT) to reveal the effect of somatic cell-derived nuclei on oocyte physiology during aging. Normal oocytes started undergoing cytofragmentation 24 hours after oocyte collection; however, this occurred earlier in SCNT oocytes and was more severe at 48 hours, suggesting that the transferred somatic cell nuclei affected oocyte physiology. We found no difference in the status of acetylated α-tubulin (Ac-Tub) and α-tubulin (Tub) between normal and SCNT aging oocytes, but unlike normal oocytes, aging SCNT oocytes did not have astral microtubules. Interestingly, aging SCNT oocytes displayed more severely scattered chromosomes or irregularly shaped spindles. Observations of the microfilaments showed that, in normal oocytes, there was a clear actin ring beneath the plasma membrane and condensed microfilaments around the spindle (the actin cap) at 0 hours, and the actin filaments started degenerating at 1 hour, becoming completely disrupted and distributed to the cytoplasm at 24 hours. By contrast, in SCNT oocytes, an actin cap formed around the transplanted nuclei within 1 hour of SCNT, which was still present at 24 hours. Thus, SCNT oocytes age in a similar but distinct way, suggesting that they not only contain nuclei with abnormal epigenetics but are also physiologically different.
Introduction
I
It has been consistently shown that the transfer of a nucleus from an aged oocyte into the enucleated cytoplasm of a fresh oocyte results in a reconstructed oocyte that has greater developmental potential than that occurs with the opposite combination (i.e., a fresh nucleus being transferred into an aged cytoplasm) (Bai et al., 2006). However, both combinations exhibit significantly lower rates of development than fresh oocytes, suggesting critical changes of both the nucleus and the cytoplasm during aging.
Somatic cell nuclear transfer (SCNT) is a valuable tool for examining the role of genetic changes and the reversibility of epigenetic marks during cell differentiation (Beetschen and Fischer, 2004). However, one serious issue with this technology is the low embryonic developmental rate, which has been largely attributed to the abnormal epigenetic status of the transplanted nuclei that have been derived from somatic cells (Ogura et al., 2013). In support of this theory, it has been shown that the treatment of SCNT embryos with histone deacetylase inhibitors such as trichostatin A significantly improves their development (Bui et al., 2010; Kishigami et al., 2006; Rybouchkin et al., 2006; Wang et al., 2007). However, it is also possible that SCNT impacts oocyte physiology, contributing to their lower developmental rate and these differences between normal and reconstructed oocytes are also expected to lead to different postovulatory aging phenotypes.
In the present study, we investigated how somatic cell-derived nuclei affect the aging and thus the quality of mouse oocytes, particularly in terms of the stability of the spindle and cytoskeleton. We also assessed the effects of somatic cell-derived nuclei on the acetylation status of α-tubulin, a marker that is used for the in vitro aging of mouse oocytes.
Materials and Methods
Animals
B6D2F1 (C57BL/6 X DBA/2) female mice aged 8–10 weeks (Japan SLC, Inc.) were used for the collection of oocytes and donor cumulus cells. Animal care and experiments were carried out in accordance with the Guiding Principles for the Care and Use of Laboratory Animals in a Kinki University Animal Facility.
Oocyte collection
The mice were superovulated by injecting them with 10 IU of pregnant mare's serum gonadotropin (Serotropin; Teikoku Zoki), followed by 10 IU of human chorionic gonadotropin (hCG, Puberogen; Sankyo) 48 hours later. Oocytes were collected from the oviducts into M2 medium 14 hours after hCG injection, and cumulus cells were removed using M2 containing 0.1% hyaluronidase. The cumulus-free oocytes were then cultured in potassium simplex optimized medium (KSOM, EmbryoMax® KSOM, MR-020P-5D; Millipore) for the experiment.
Experimental groups
This study examined oocyte aging in three experimental groups: “MII” oocytes, normal metaphase II oocytes without any manipulation; “ENU” oocytes, enucleated oocytes without any nuclear transfer; and “SCNT” oocytes, standard, enucleated oocytes that were reconstructed by cumulus cell nuclear transfer (Fig. 1A).
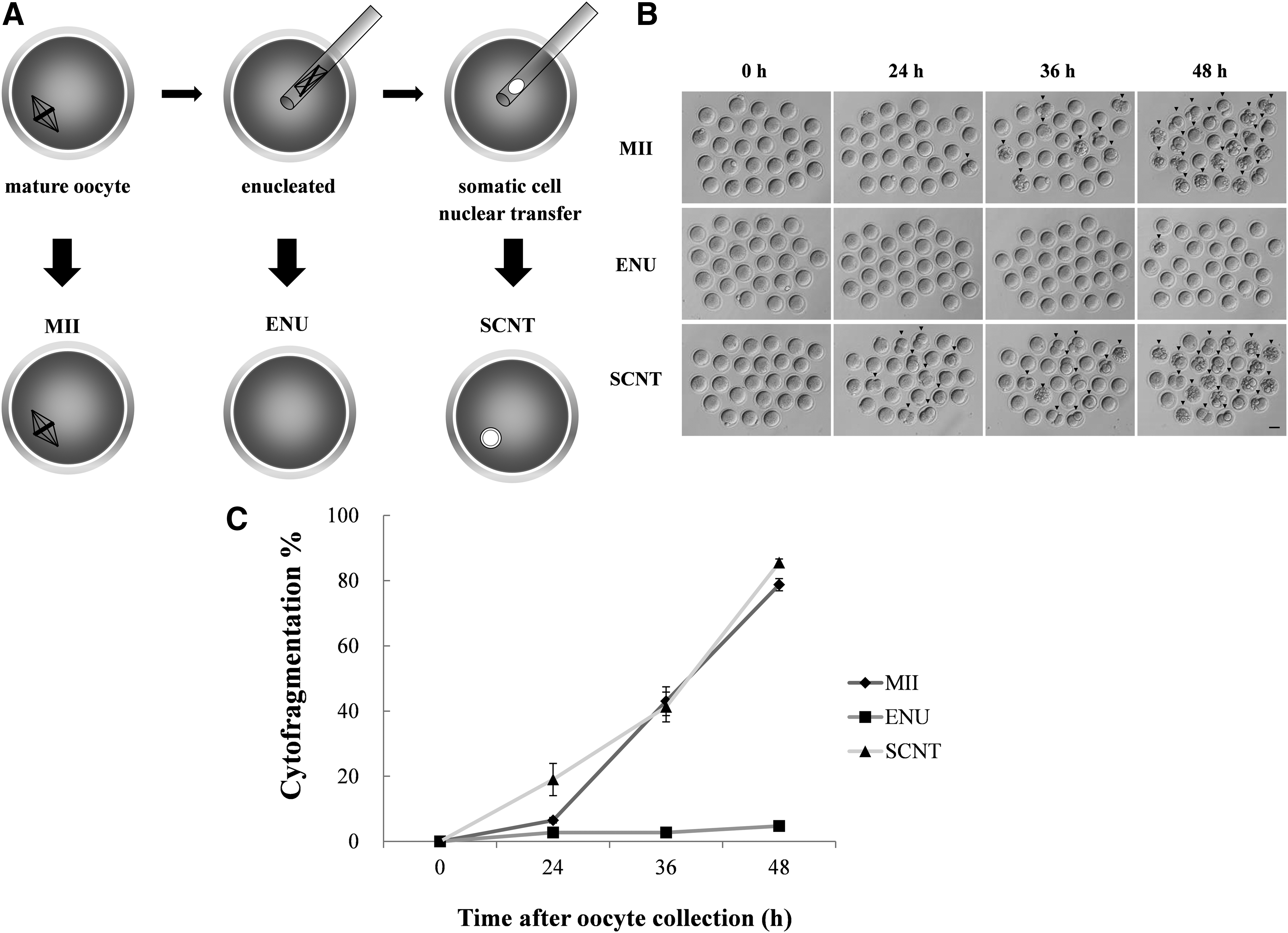
Morphological changes in normal MII, ENU, and SCNT oocytes during postovulatory aging.
Nuclear transfer procedure
Cumulus cell nuclear transfer was performed using a previously described method (Wakayama et al., 1998). Briefly, the B6D2F1 oocytes were enucleated and then injected individually with cumulus cells from B6D2F1 mice using a Piezo-driven micromanipulator (PrimeTech Corporation). After nuclear transfer, the reconstructed oocytes were cultured in KSOM and observed at 0, 24, 36, and 48 hours.
Immunofluorescence
Oocytes were washed in phosphate-buffered saline containing 0.1% polyvinyl alcohol (PBS-PVA) and then fixed in 4% w/v paraformaldehyde at room temperature for 30 minutes. The oocytes were then washed in PBS-PVA and incubated overnight in PBS containing 1% bovine serum albumin (BSA) and 0.1% Triton X-100 at 4°C. After this, the oocytes were washed three times in PBS-0.1% BSA and incubated with a 1:200 dilution of a rabbit monoclonal anti-α-tubulin primary antibody (T3526; Sigma) and a 1:1000 dilution of a mouse monoclonal anti-acetylated α-tubulin (Ac-Tub) primary antibody (T7451; Sigma) for 2 hours at room temperature. The oocytes were then washed three times in PBS-0.1% BSA and incubated with a 1:200 dilution of goat anti-mouse or a 1:200 dilution of chicken anti-rabbit secondary antibodies for 1 hour at room temperature. After this, they were washed in PBS-0.1% BSA a further three times. To stain the actin filaments (F-actin), the oocytes were exposed to 14 μM/500 μL Actin-stain™ 555 fluorescent phalloidin with PBS containing 0.1% BSA for 1 hour at room temperature.
The DNA was visualized by staining the oocytes with 4′,6-diamidino-2-phenylindole (DAPI). The oocytes were mounted on glass slides with a drop of fluorescent mounting medium and then covered with a glass cover slip. We observed the cytoplasm and spindle in each of the aging oocytes using fluorescence microscopy.
Statistical analysis
Data for the normal and reconstructed oocytes were analyzed using a Z-test. A value of p < 0.05 was considered statistically significant.
Results
Fragmentation in SCNT oocytes during aging
Very few MII oocytes exhibited abnormal morphologies at 24 hours, but more than 80% had undergone cytofragmentation by 48 hours (Fig. 1B, C). By contrast, most of the ENU oocytes showed no fragmentation even at 48 hours (Fig. 1B, C). The SCNT oocytes underwent cytofragmentation at an earlier stage than the MII oocytes, with ∼20% exhibiting this at 24 hours; however, there was no significant difference between MII and SCNT oocytes after 36 hours (Fig. 1B, C).
We classified the fragmented oocytes into three grades according to the extent of fragmentation: Grade 1 = one large cytoplasm with small cytoplasmic droplets (Fig. 2A-b); Grade 2 = two-cell-like oocytes with some small cytoplasmic droplets, often clustered in the crevice between the blastomeres (Fig. 2A-c); and Grade 3 = totally fragmented oocytes, where all of the cytoplasmic droplets were less than a quarter of the size of the oocyte (Fig. 2A-d). This categorization revealed that a larger number of the aged SCNT oocytes were severely fragmented and in Grade 3 at 48 hours than the other types of oocytes (Fig. 2B).
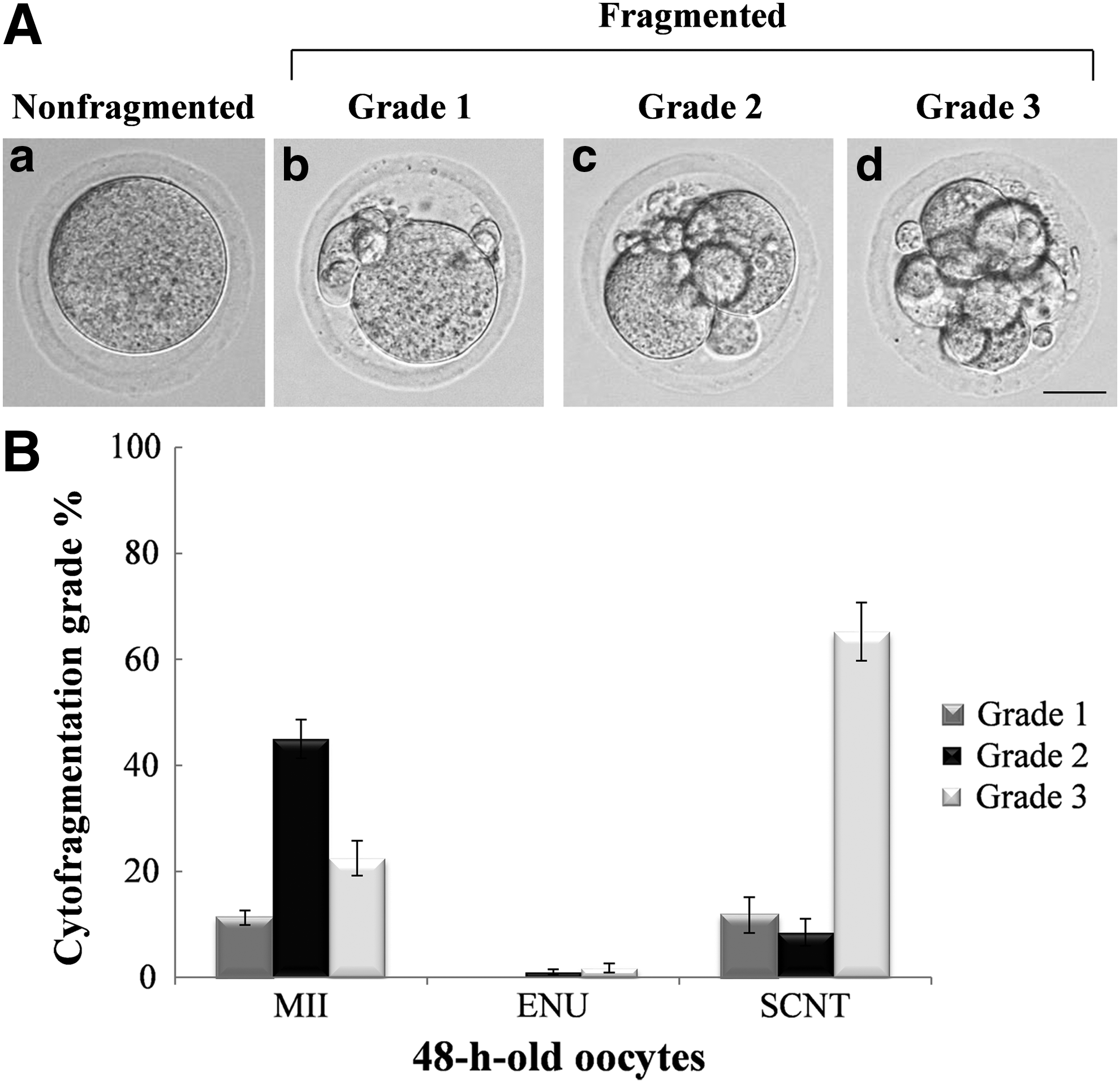
Grades of fragmentation in normal MII, ENU, and SCNT oocytes after aging.
Acetylation status of α-tubulin in SCNT oocytes during aging
In all three types of oocytes (MII, EN, and SCNT), the amount of Ac-Tub and Tub in the cytoplasm gradually increased until 24 hours and then began decreasing at 36 hours. In aged MII oocytes, Ac-Tub was assembled into astral microtubules at 36 hours. Unexpectedly, ENU oocytes also dynamically assembled astral microtubules; however, this occurred 24 hours earlier than in the MII oocytes and so these were more strongly expressed in the cytoplasm after 36 hours. By contrast, the SCNT oocytes did not exhibit any astral microtubules in the cytoplasm even at 36 hours (Fig. 3).
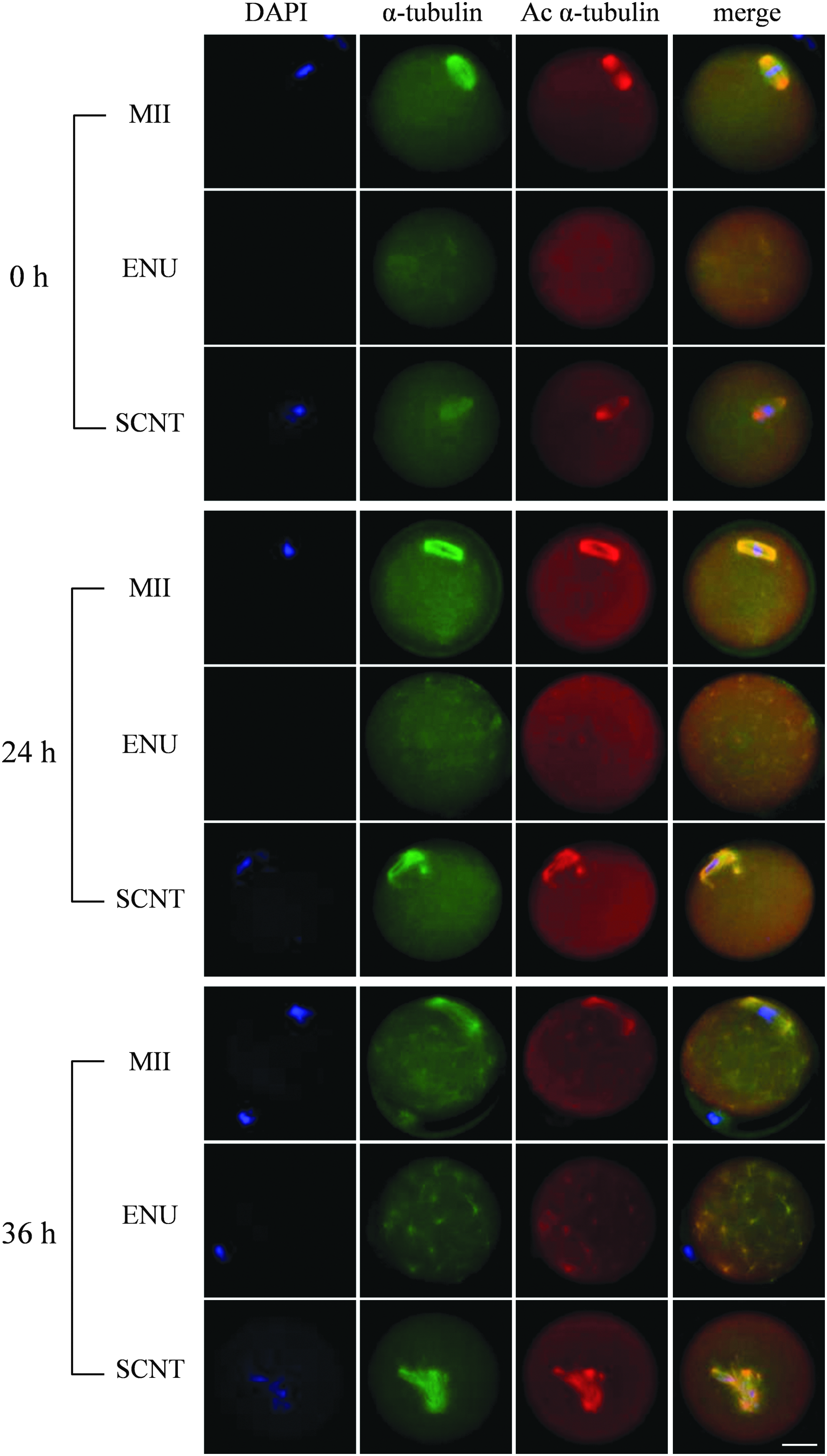
The behavior of α-tubulin in aging normal MII, ENU, and SCNT oocytes 0, 24, and 36 hours after immunostaining with anti-Ac α-tubulin and α-tubulin antibodies. All three types of oocytes showed hyperacetylation after 24 hours, but the astral microtubule structures appeared at different times in each. Ac, acetylated.
Spindle morphology in SCNT oocytes during aging
At 6 hours, SCNT oocytes exhibited a disorganized spindle shape and chromosome misalignment, with a lower content of both acetylated α-tubulin and α-tubulin (Fig. 4). At 24 hours, all SCNT oocytes exhibited a multipolar spindle shape and chromosome scattering (Table 1). By contrast, most MII oocytes (72%) exhibited spindle elongation at 24 hours (Table 1). At 36 hours, the SCNT oocytes exhibited multipolar spindles with extensive chromosome scattering (Fig. 4). Thus, spindle morphology and chromosome alignment in aging SCNT oocytes were characteristically distinct from those seen in normal oocytes.
Different superscript letters denote significant differences (p < 0.05) between the different types of oocytes at the same age.
ENU, enucleated; MII, metaphase II; SCNT, somatic cell nuclear transfer.
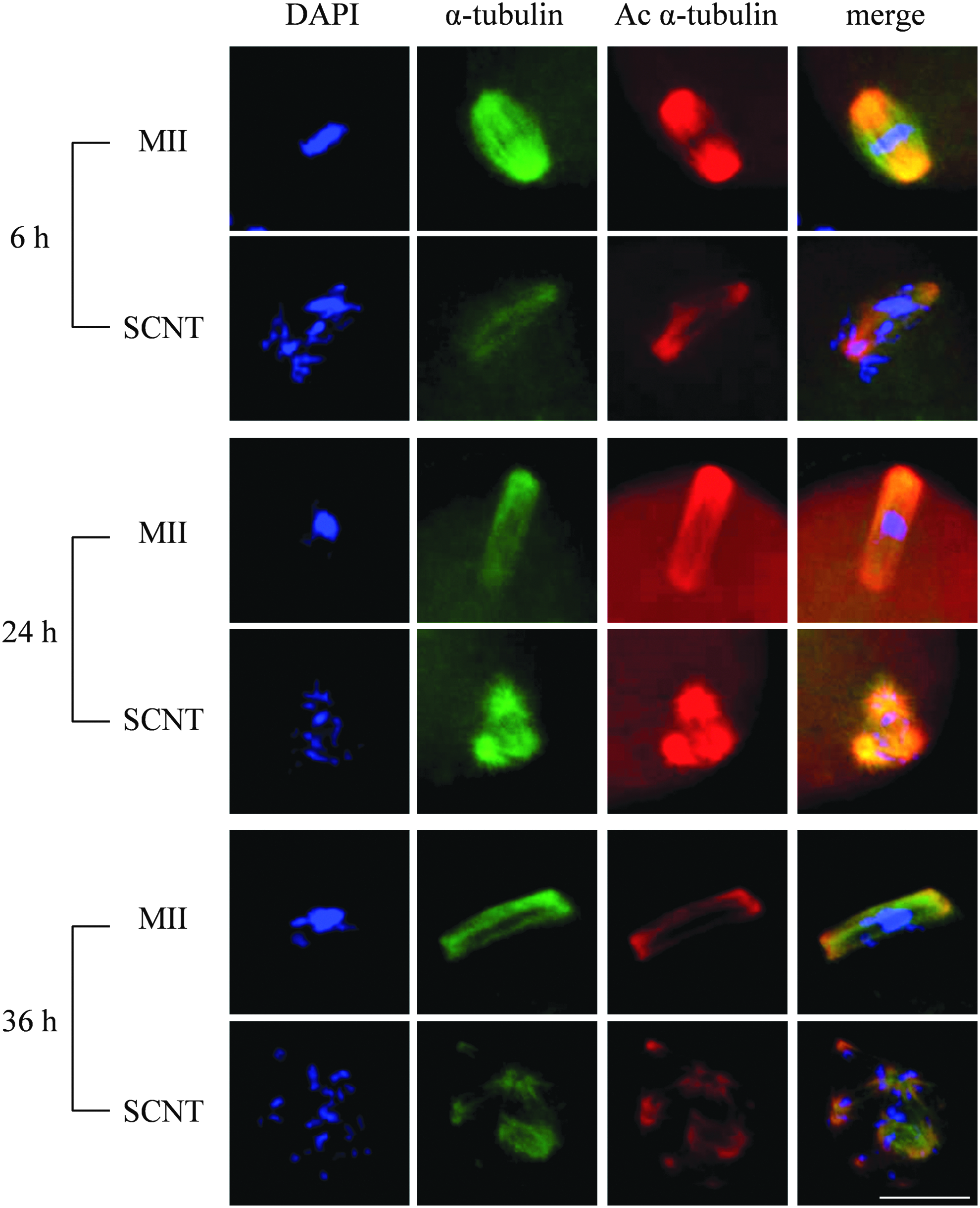
Spindle morphology and chromosome scattering in aging normal MII and SCNT oocytes 0, 24, and 36 hours after immunostaining with anti-Ac α-tubulin and α-tubulin antibodies. Distinct patterns in the spindles of these two types of oocytes can be seen.
Behavior of microfilaments in SCNT oocytes during aging
No obvious changes were observed in the cortical filaments in MII and ENU oocytes by 1 hour (Fig. 5). Within 1 hour, the SCNT oocytes formed an actin cap near the spindle that had a similar morphology to that in MII oocytes, whereas this was not observed in the ENU oocytes. In MII oocytes, the signal of the cortical filament, including the actin cap, had weakened by 3 hours, and only a faint actin cap could be seen at 24 hours. However, unexpectedly, the SCNT oocytes maintained the actin cap even after 24 hours (Fig. 5).
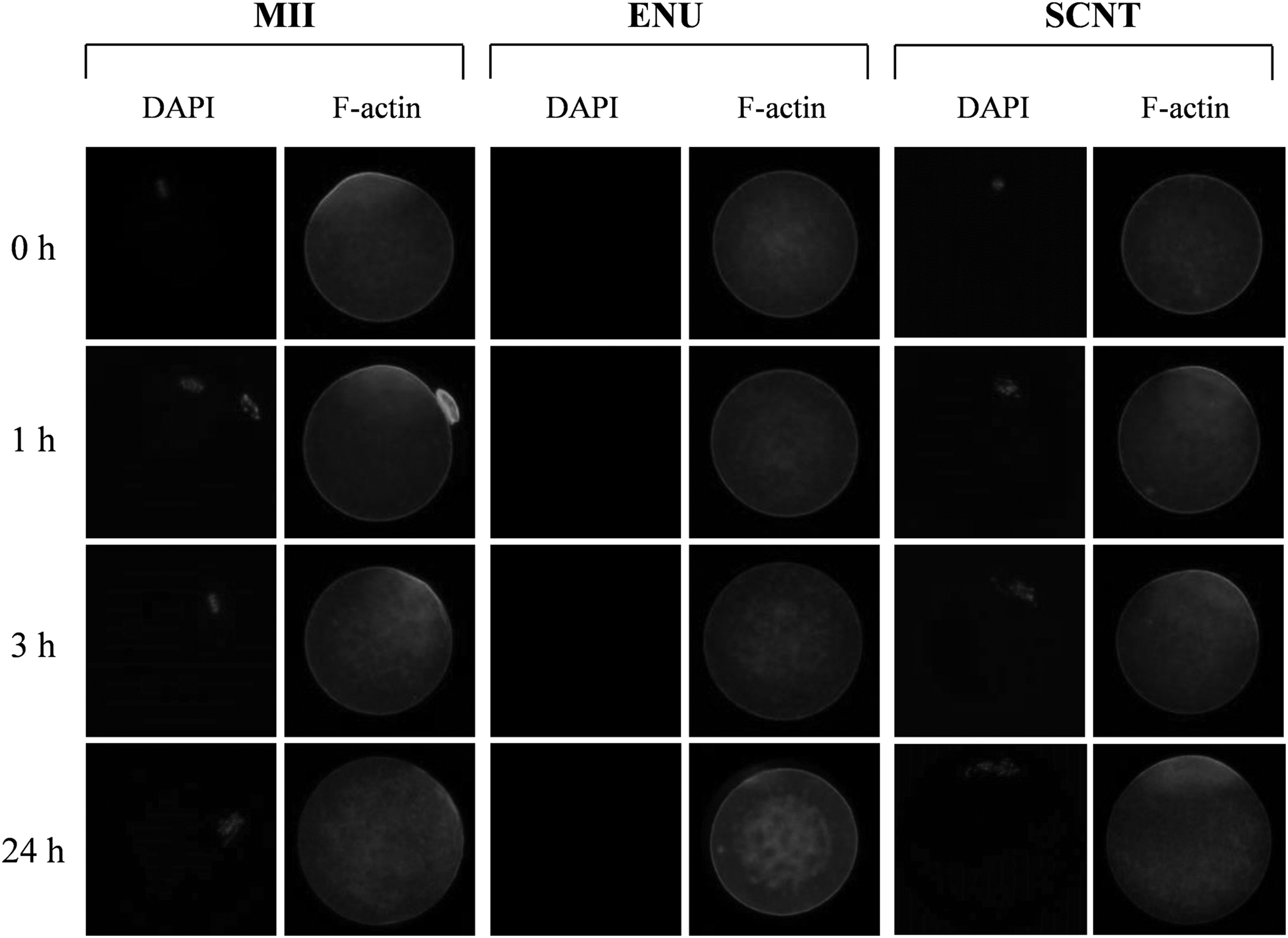
The behavior of F-actin in aging normal MII, ENU, and SCNT oocytes 1, 2, 3, and 24 hours after staining the microfilaments with anti-F-actin antibody. The actin caps were recognized as a strong signal around the spindle. During aging, the microfilaments of the MII oocytes were disrupted and distributed to the cytoplasm; whereas the SCNT oocytes maintained the actin cap even at 24 hours.
Discussion
We observed cytofragmentation of MII oocytes during prolonged in vitro culture (spontaneous fragmentation), supporting the findings of previous studies (Lanman, 1968; Lee et al., 2013; Sakai and Endo, 1988). It has previously been shown that cytofragmentation is dependent on the functional expression of several genes comprising the evolutionarily conserved apoptotic cell death program (Morita et al., 1999; Perez et al., 1999). Therefore, we aimed at demonstrating the role that the nucleus plays in controlling cytofragmentation during oocyte aging. We found that the ENU oocytes retained a normal morphology even at 48 hours, as previously found (Alikani et al., 2005). By contrast, although some of the SCNT oocytes started undergoing cytofragmentation at an earlier stage than the MII oocytes, most underwent fragmentation over a similar time course, demonstrating that even exogenous nuclei can cause cytofragmentation. Thus, these results suggest that a nucleus may play an important role in the initiating stimulus of apoptosis by way of such as oxidative-stress-induced DNA damage along with postovulatory aging (Takahashi et al., 2003; Tang et al., 2013). Further studies are needed to elucidate the precise mechanism. Interestingly, when somatic cell-derived nuclei were injected into the ENU oocytes, the extent of cytofragmentation was greater than that observed in aged MII oocytes, possibly due to an abnormal configuration and stability of the cytoskeleton, including actin and tubulin. Thus, our study shows that nuclear factors are essential for cytofragmentation during oocyte aging and that SCNT oocytes have an aging phenotype that is similar but distinct from normal MII oocytes.
Oocyte aging results in the hyperacetylation of histone (Huang et al., 2006), and we previously demonstrated that the hyperacetylation of α-tubulin also occurs during prolonged culture in vitro (Lee et al., 2013). Normal mouse oocytes exhibit microtubule acetylation only after fertilization and early development (Matsubara et al., 2013; Schatten et al., 1988). In this study, the localization and nuclear progression of microtubules in SCNT oocytes were visualized by immunostaining Tub and Ac-Tub. We found that the amount of Ac-Tub and Tub in SCNT and ENU oocytes gradually increased until 24 hours and then started decreasing again at 36 hours, which is similar to the pattern seen in aging MII oocytes (Lee et al., 2013). These results suggest that tubulin acetylation is increased by cytoplasmic aging, independent of the presence of a nucleus.
We previously demonstrated an interphase-like organization of the cytoskeleton in aged MII oocytes, with astral microtubules frequently being observed at 36 hours (Lee et al., 2013). Therefore, we investigated the stability of astral microtubules in SCNT oocytes during aging. In this study, Ac-Tub was also dynamically assembled into astral microtubules in the cytoplasm at 36 hours in the MII oocytes. However, the ENU oocytes started assembling Ac-Tub into astral microtubules 24 hours earlier than the MII oocytes, and had more of these in the cytoplasm after 36 hours, which is consistent with the previous report (Alikani et al., 2005). By contrast, SCNT oocytes had significantly fewer astral microtubules in the cytoplasm even at 36 hours. Instead, more Ac-Tub was more accumulated around the spindle, suggesting that they participated in the formation of the pseudo-spindle, including the somatic cell-derived chromosomes in return for the formation of astral microtubules (Miki et al., 2004). These results suggest that Ac-Tub itself is not directly correlated with the occurrence of astral microtubules and that the assembly of astral microtubules may be associated with the amount of Ac-Tub free from the spindle.
It is known that both cytoplasmic and nuclear factors are important for metaphase chromosome alignment and the spindle checkpoint during meiosis (Liu and Keefe, 2004). Abnormalities in the metaphase II spindle, including misaligned chromosomes, or dispersed, elongated, or completely disrupted spindles, have commonly been observed in aging oocytes (Wakayama et al., 2004). We found that within 6 hours of being injected with somatic cells, SCNT oocytes reconstructed the spindle, which is consistent with our previous observation (Thuan et al., 2006), and that after 24 hours, abnormal, multipolar spindles with chromosome scattering occurred in these aging oocytes, which is not usually observed in aged MII oocytes. Thus, the spindle that contained somatic cell-derived chromosomes had a much higher incidence of chromosome scattering, potentially due to its lower stability.
Microfilament dynamics plays a critical role in asymmetric spindle positioning and cortical polarization during meiotic division (Almonacid et al., 2014). The distribution of this cortical actin is changed during meiotic maturation (Longo, 1987), and the actin cap is critical for the asymmetrical division of the oocyte, which maintains the maternal component for early embryo development (Maro and Verlhac, 2002). It has previously been shown that a thick microfilament domain in the oocyte cortex, including the cortical apical cap, is disrupted in aged oocytes (Kim et al., 1996; Miao et al., 2009). Therefore, we also investigated the dynamics of the microfilaments in MII, ENU, and SCNT oocytes. We found that after 3 hours, the actin cap started becoming disrupted and was distributed to the cytoplasm in the MII oocytes. By contrast, the ENU oocytes did not make an actin cap and aggregated to the center of the cytoplasm at 24 hours, whereas the SCNT oocytes constructed an actin cap at 1 hour after nuclear transfer and retained this even at 24 hours. Actin is a major constituent of the cytoskeleton (Miyamoto et al., 2011; Visa and Percipalle, 2010), dynamically changing the polymerized state of the cytoplasm with the help of actin nucleators and actin signaling protein (Campellone and Welch, 2010). Therefore, our results indicate that the nuclei derived from somatic cells formed microfilaments that were localized in the actin cap and that have greater stability than those found in MII oocytes. Thus, our results suggest distinct differences in the dynamics of α-tubulin and actin between MII, ENU, and SCNT oocytes during aging.
In this study, we revealed similar but distinct phenotypes of aged SCNT and MII oocytes, demonstrating the effect of somatic cell-derived nuclei on oocyte physiology. Our observations provide insight into the mechanisms underlying oocyte aging and the lower developmental potential of SCNT embryos, as well as of their abnormal epigenetic states.
Footnotes
Acknowledgments
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.