
Abstract
Abstract
Cataract, the leading cause of blindness worldwide, is caused by the apoptosis of lens epithelial cells (LECs). αA-crystallin is a major structural protein of the lens. However, the antiapoptotic function of αA-crystallin in lens stem cells remains unclear. In this study, primary LECs were isolated from postnatal 3–5 days of SD rats and transfected by Sendai virus loaded with four factors, OCT3/4, Sox2, c-Myc, and Klf4, to induced pluripotent stem cells (iPSCs). LEC-iPSC-like cells were identified by immunofluorescent staining. CryαA-specific shRNA lentivirus was used to knockdown αA-crystallin in LEC-derived iPSC-like cells, which were treated with tert-Butyl hydroperoxide. The apoptosis of LEC-iPSC-like cells was examined by flow cytometry. We reprogrammed LECs and obtained embryonic stem cell-like colonies. LEC-iPSC-like cells with normal karyotype expressed pluripotent markers such as SSEA-4, TRA-1-60, and TRA-1-81. Knockdown of αA-crystallin increased the apoptosis of LEC-iPSC-like cells and rendered them less resistant to oxidation stress induced by tert-Butyl hydroperoxide. In conclusion, LECs could be reprogrammed into iPSC-like cells and αA-crystallins could protect LEC-iPSC-like cells from oxidation stress-induced apoptosis.
Introduction
Y
Cataract is generally defined as any opacity in the lens, and cataract disease represents an important health problem that accounts for 46% of all cases of blindness (Zhang et al., 2010). LEC apoptosis is an initiating factor in noncongenital cataract formation, and reactive oxygen species can damage the human lens and contribute to cataract formation. Studies from different animal models provide strong evidence that stress-induced apoptosis is the common initiating event during cataractogenesis (Kalariya et al., 2010).
α-crystallin is one of the major structural proteins of the lens, accounting for nearly 40% of the total proteins in the lens, and is required for the maintenance of lens transparency (Bloemendal, 1982). α-crystallin confers protection against many eye diseases, including cataract, retinitis pigmentosa, and macular degeneration (Christopher et al., 2014). In α-crystallin knockout animals, LECs become genomically unstable and undergo enhanced apoptosis in the absence of α-crystallin (Yaung et al., 2007). In addition, cells treated with exogenous α-crystallin protein had reduced apoptosis upon stress from oxidative or thermal insults (Christopher et al., 2014). The two forms of α-crystallin (αA-crystallin and αB-crystallin) share 57% sequence homology and form multiple complexes with each other at the ratio of 3:1 (Gopalakrishnan and Takemoto, 1992). At a given expression level, the antiapoptotic activity of αA-crystallin was significantly higher compared with αB-crystallin (Andley et al., 2000).
Understanding of the molecular events involved in LECs apoptosis may help develop therapeutic strategies to prevent or delay cataractogenesis. In this study, we used shRNA lentivirus to knockdown the expression of CRYAA gene in LEC-derived iPSC-like cells and examined the apoptosis rate of iPSC-like cells under oxidative stress. Our results suggest that αA-crystallin protects iPSC-like cells from oxidative stress-induced apoptosis.
Materials and Methods
Establishment of iPSC-like cells from rat LECs
LECs were isolated from postnatal 3–5 days SD rats (provided from Laboratory Animal Department, XiangYa School of Medicine, Central South University). The capsules of the rat lens were cut into small pieces and digested with 0.25% Trypsin-EDTA (Gibco) for 10 minutes, then plated in a dish with Dulbecco's modified Eagle's media/Nutrient Mixture F-12 (DMEM/F12; Hyclone) supplemented with 20% fetal bovine serum (Hyclone), 100 U/mL penicillin (Sigma), 100 μg/mL streptomycin (Sigma), and 0.5 mL epithelial growth factor (EpiCGS; Sciencell), and cultured in 100% humidity at 37°C in 5% carbon dioxide atmosphere. Sendai virus loaded with four factors, Oct3/4, Sox2, c-Myc, and Klf4 (CytoTune®-iPS 2.0 Sendai Reprogramming Kit; Life Tech), were used to reprogram rat LECs.
Rat LEC-derived iPSC-like cells were cultured on mitomycin C-treated mouse embryonic fibroblast (MEF) feeder layer in embryonic stem (ES) culture medium containing DMEM/F12 (Hyclone) supplemented with 20% knockout serum replacement (Gibco), 0.1 mM 2-mercaptoethanol (Amresco), 0.1 mM nonessential amino acid (Gibco), and 4 ng/mL bFGF (basic fibroblast growth factor) (Sigma).
Immunofluorescent staining
Rat LECs and LEC-derived iPSCs were fixed in 4% paraformaldehyde, washed with phosphate-buffered saline (PBS; Hyclone) three times, permeabilized with PBS containing 0.1% Triton X-100 for 10 minutes, and incubated with PBS containing 4% bovine serum albumin (PBS-B; Sigma) for 30 minutes to block nonspecific reactions. LECs were incubated overnight at 4°C with the following antibody: anti-αA-crystallin (1:200; Santa Cruz); iPSs were incubated overnight at 4°C with the following antibodies: anti-OCT4, anti-Sox2, anti-nanog, anti-SSEA-4, anti-TRA-1-60, and anti-TRA-1-81 (1:200; Chemicon). Next, the cells were washed three times with PBS and incubated with fluorescence-conjugated secondary antibodies (Zhongshanjinqiao Biotech, Beijing) in PBS-B for 1 hour at room temperature and then incubated with DAPI (Beyotime Institute of Biotechnology) for 1 minute. Images were obtained with an inverted fluorescence microscope (LEICA).
Construction of aA-crystallin shRNA lentiviruses
The shRNA target sequence (5′-GACTGTTCGACCAGTTCTTCG-3′) was designed for CryαA gene, and nonspecific sequence (5′-GGTTTGGCATATAATCTATCG-3′) was inserted into the lentiviral vector LV-008 (Forevergen Biosciences, China), which was used to express small hairpin RNAs with GFP (greenfluoroscent protein) as a reporter. Lentiviral production was performed as a routine. Briefly, LV-008-shCryαA plasmids and packaging vectors were cotransfected into HEK 293T cells, and the supernatant containing lentiviruses was collected 72 hours post-transfection. The lentiviruses were concentrated by ultracentrifugation for 1.5 hours at 25,000 rpm and resuspended in PBS. For lentivirus infection, LEC-iPSCs were cultured in a six-well plate and infected with lentivirus in the presence of 5–10 μg/mL of polybrene. Cells were screened with 2 μg/mL puromycin for 10–15 days to get αA-crystallin knockdown cells.
Western blot analysis
Cells were harvested and washed with PBS and then lysed in lysis buffer (50 mM Tris, 150 mM NaCl, 1% Triton X-100, 0.5% deoxycholate, 0.5 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride, 1 mg/mL aprotinin, and 2 μg/mL leupeptin). Proteins were quantified and electrophoresed through 10% SDS-PAGE gel and transferred to polyvinylidene difluoride membranes (Millipore). Membranes were blocked for 1 hour in Tris-buffered saline with 0.05% Tween-20 (TBST) containing 5% (w/v) nonfat milk, incubated with αA-crystallin antibody (1:200; Santa Cruz) and β-actin antibody (1:200; Santa Cruz) for 2 hours, washed twice with TBST for 15 minutes, and then incubated with peroxidase-conjugated goat antibody to mouse IgG (1:200; Amersham Pharmacia Biotech) for 30 minutes. After washing three times, the membranes were visualized by enhanced chemiluminescence system (Amersham Pharmacia Biotech).
FACS analysis of apoptosis
Quantification of apoptotic LEC-iPSCs was determined by FITC-conjugated annexin V and PI (Apoptosis Detection Kit; Oncogene) staining. After treatment, cells were washed in PBS and centrifuged at 200 g for 5 minutes. Cell pellets were resuspended and stained according to the manufacturer's instructions. Flow cytometry was performed using a guava easycyte 8HT FACScan.
Results
Cultrue and maintenance of primary LECs
We cut the lens capsules into small pieces and digested them with 0.25% Trypsin-EDTA to separate the cells. After culture for 2 hours in epithelial DMEM/F12 medium, some lens capsular tissues started to attach to the bottom of the flasks. After culture for 24 hours, most cells attached to the bottom of the dishes and showed flattened and irregular morphology (Fig. 1A). Starting from the forth passage, the cells proliferated rapidly, dedifferentiated, and displayed a spindle morphology as the fibroblast (Fig. 1B).
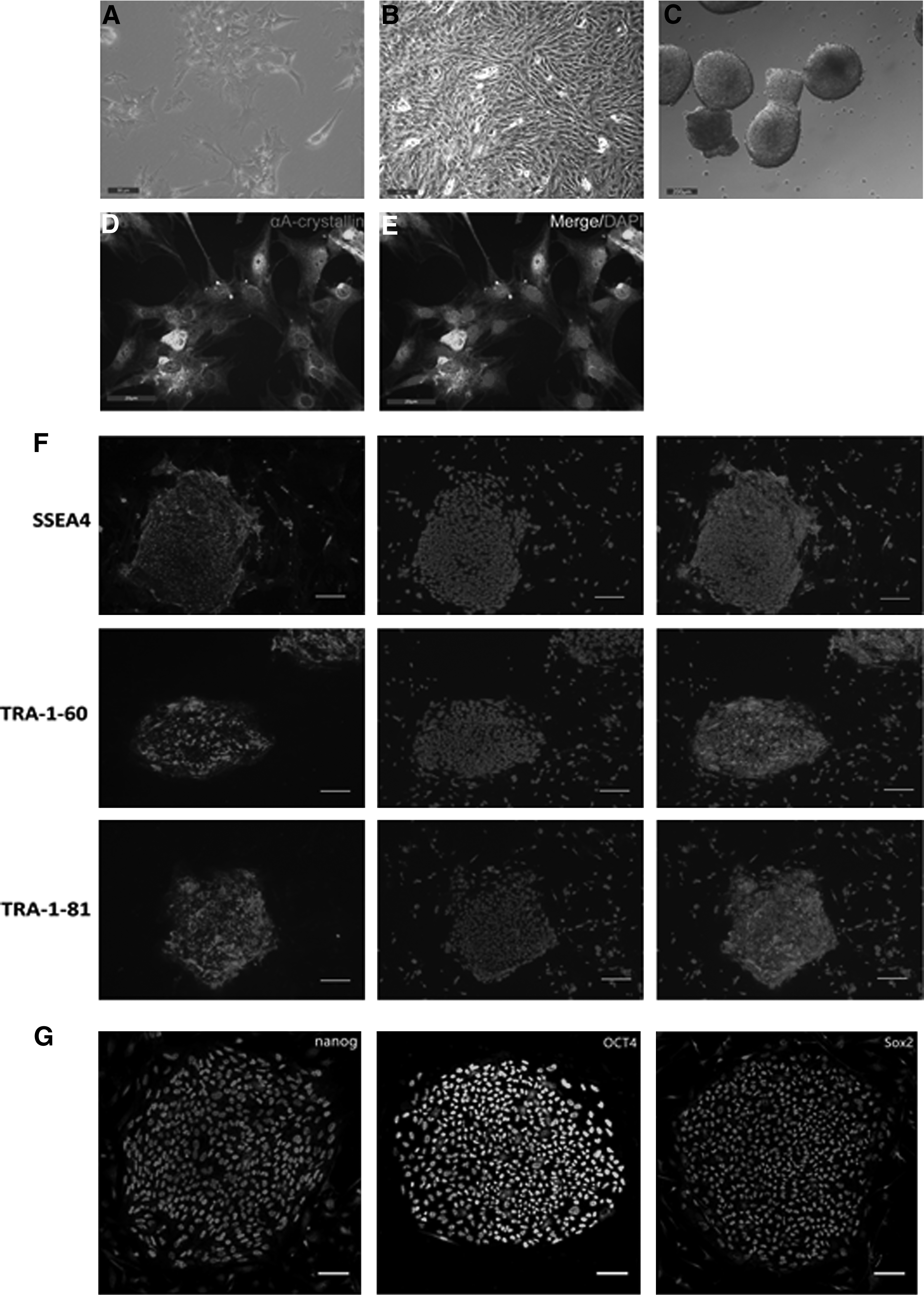
Preparation and characterization of LECs.
We selected the third passage cells for reprogramming and performed immunofluorescent staining to detect LEC-specific marker αA-crystallin. We observed that almost 100% cells expressed αA-crystallin (Fig. 1D, E).
Generation of iPSC-like cells from primary LECs
LECs at passage 3 were transduced with Sendai virus containing Oct4, Sox2, Klf4, and c-Myc at 1:1:2:1 ratio on day 0. On day 6 after the transduction, cells were transferred onto mitomycin C-treated MEF feeder layers and cultured in ES culture medium. From day 9, we observed tightly packed colonies with morphologies similar to ESC, and the size of ESC-like colonies increased over time (Fig. 1C).
LEC-iPSC-like cells share key expression markers with ESCs
We next characterized LEC-iPSC-like cells. Immunofluorescence staining with antibodies against SSEA-4, TRA-1-60, and TRA-1-81 showed that these cells expressed these three typical ESC surface markers (Fig. 1F). In addition, these cells expressed pluripotency markers OCT4, Sox2, and nanog (Fig. 1G). Taken together, these data suggest that LEC-derived iPSC-like colonies share the majority of key markers of ESCs.
shRNA letiviruses-mediated knockdown of αA-crystallin in LEC-derived iPSC-like cells
We tranduced LEC-iPSC-like cells with lentiviruses harboring CryαA-specific shRNA or control shRNA. Both viral constructs expressed GFP from an independent promoter (Fig. 2A). Under fluorescence microscope we observed that most cells were efficiently infected by shRNA. Western blot analysis showed that αA-crystallin expression was significantly reduced in CryαA-specific shRNA group compared to control shRNA group (Fig. 2B). Quantification of band intensity showed that αA-crystallin expression was significantly reduced in CryαA-specific shRNA group compared to the control group (Fig. 2C).
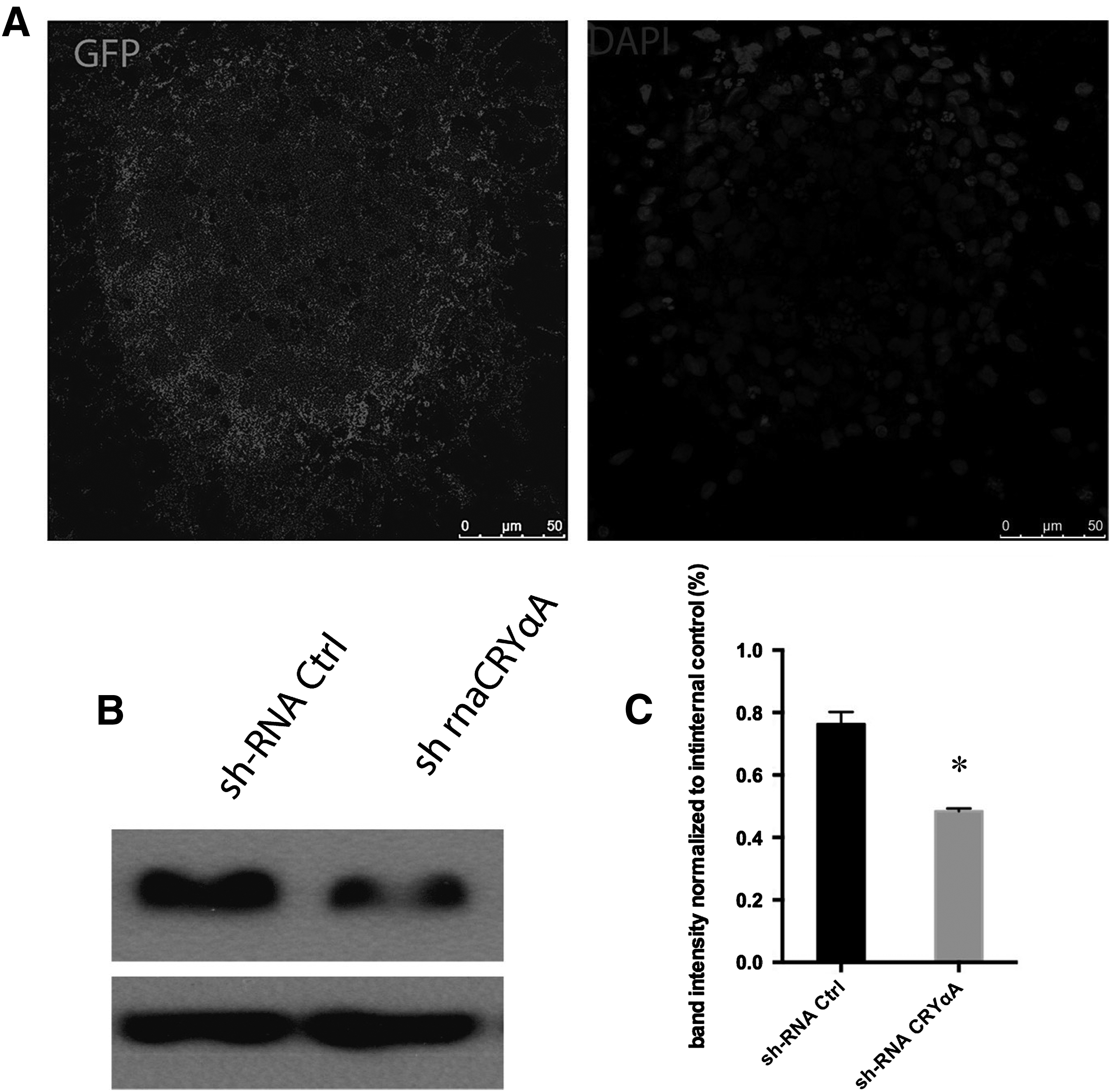
Knockdown of CryαA in LEC-iPSC-like cells.
αA-crystallin knockdown increases oxidation stress-induced apoptosis of LEC-iPSC-like cells
Next, we performed shRNA-mediated knockdown of αA-crystallin in LEC-iPSC-like cells treated with or without 200 μM tert-Butyl hydroperoxide (TBHP) for 12 hours. We found that knockdown of αA-crystallin significantly increased annexin V-FITC-stained apoptotic cells both in the treated and untreated LEC-iPSC-like cell groups. In the untreated control group, the percentage of apoptotic cells was 3.8% and 7.3%, respectively, after the introduction of shRNA control and shRNA-CryαA. In contrast, in TBHP-treated group the percentage of apoptotic cells was increased to 13.1% and 22.5%, respectively, after the introduction of shRNA control and shRNA-CryαA (Fig. 3). These data suggest that knockdown of αA-crystallin promotes oxidation stress-induced apoptosis of LEC-iPSC-like cells.
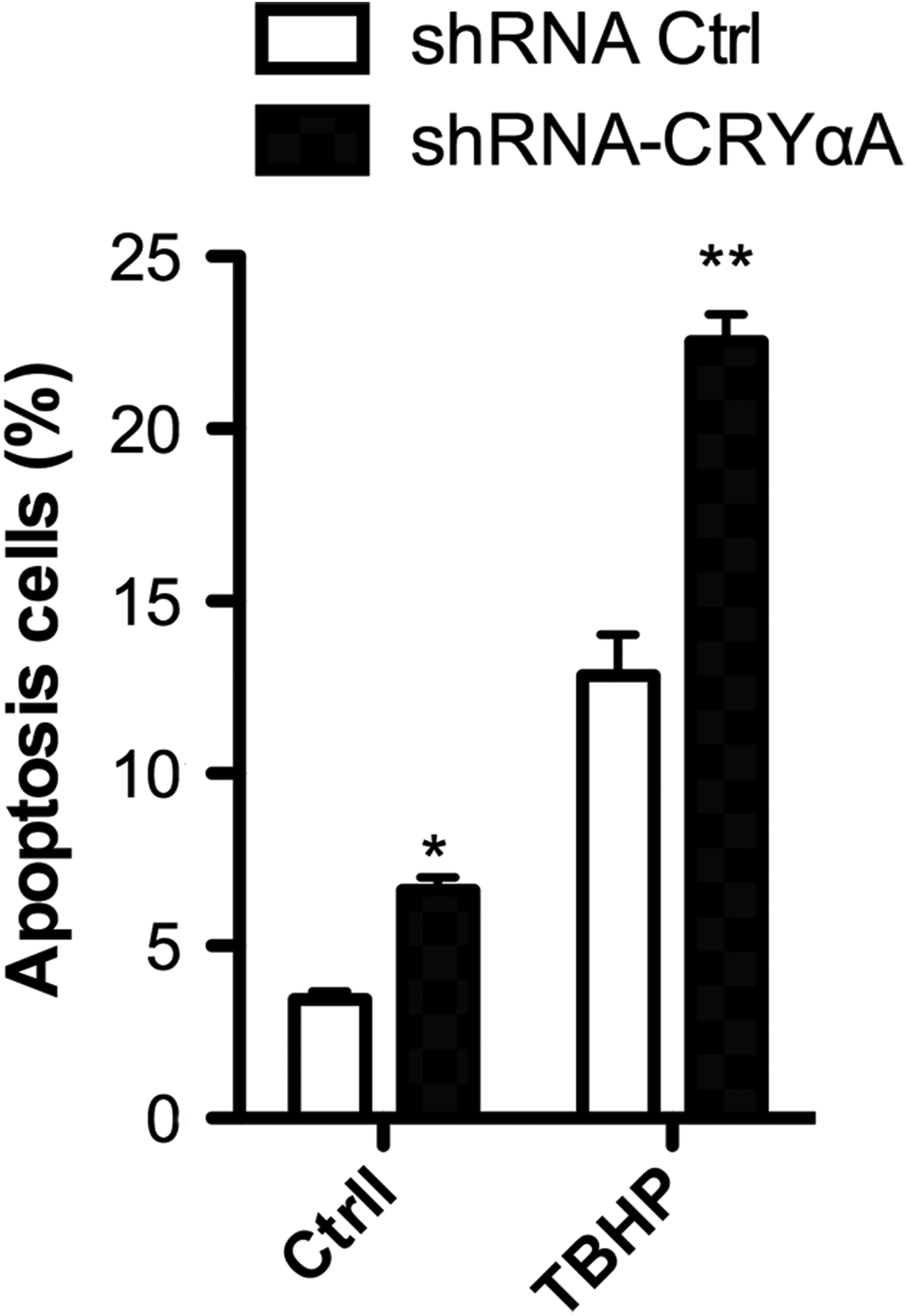
TBHP increased apoptosis of LEC-iPSC-like cells. LEC-iPSC-like cells were tansduced with CryαA-specific shRNA or control shRNA and treated with or without TBHP. Apoptosis was detected by flow cytometry. Data are expressed as mean ± SD (n = 5). *P < 0.05 compared to cells treated with control shRNA. **P < 0.01 compared to cells treated with control shRNA and TBHP. TBHP, tert-Butyl hydroperoxide.
Discussion
In this study, we developed a procedure for generating iPSC-like cells from LECs and showed that αA-crystallins were expressed in LEC-derived iPSC-like cells and protected them from oxidative stress-induced apoptosis.
iPSCs generally exhibit a normal karyotype, are transcriptionally and epigenetically similar to ESCs, and maintain the capacity to differentiate into derivatives of all three germ layers (Takahashi and Yamanaka, 2006). In this study, we showed that LEC-derived colonies were indistinguishable from ESCs with respect to the morphology and the expression of pluripotent markers, thus, we defined them as iPSC-like cells, although we could not completely characterize the induced cells to cofirm that they are indeed iPSCs. LEC-derived iPSC-like cells can be highly and efficiently differentiated into large numbers of lens progenitor cells through the use of defined factors, and they are more suitable as the candidate for the therapy of lens disease than other cell line-derived iPSCs.
It is clear that apoptosis is a general feature of lens development and pathogenesis (Yan et al., 2006). During lens development, the detachment of the lens vesicle from corneal ectoderm and the crafting of the lens vesicle rely on timely apoptosis (Blixt et al., 2000; Mohamed and Amemiya, 2003). Apoptosis helps to remove aberrantly differentiated lens cells or damaged epithelial cells (Bozanić et al., 2003). When the normal apoptotic process is blocked either by pharmacological drugs or through inactivation of apoptosis regulators, cataratogenesis may develop (Zhang et al., 2010). In addition, stress-induced apoptosis is the common initiating event during cataractogenesis (Yan et al., 2006). LEC apoptosis was associated a loss of rat lens transparency and could be stimulated by oxidative stress (Spector, 1995). As an oxidative stress, TBHP can result in lipid oxidation, which is known to deplete intracellular pools of calcium and cause apoptosis (Spector et al., 2002). In this study, we demonstrated that TBHP promoted the apoptosis of LEC-iPSC-like cells.
Transcripts for αA-crystallin can be detected in LECs at early stages of mouse lens development (Robinson et al., 1995). In this study, we confirmed that LEC-iPSC-like cells expressed αA-crystallin. Moreover, we found that Lentivirus-mediated knockdown of αA-crystallin increased LEC-iPSC-like cell apoptosis rate and decreased their ability to resist TBHP-induced apoptosis. Thus, αA-crystallin expression can protect LEC-iPSC-like cells from apoptosis and oxidative stress. α-crystallin is the major soluble protein in the lens and plays a critical role in the maintenance of lens transparency (Gopalakrishnan and Takemoto, 1992). αA-crystallin is also a member of the small heat shock protein family. The inactivation of heat shock transcription factor 4 in mice caused cataract at early postnatal days (Fujimoto et al., 2004; Min et al., 2004; Shi et al., 2009). αA-crystallin provides protection against a wide range of cellular stresses, because cells overexpressing αA-crystallin were more resistant to thermal, osmotic, and oxidative stress (Andley et al., 2002; Aoyama et al., 1993; Dasgupta et al., 1992).
Recent studies have shown that αA-crystallin prevented apoptosis induced by a variety of agents, including ultraviolet light, staurosporine, etoposide, tumor necrosis factor-α, and hydrogen peroxide (Andley et al., 2000; Mao et al., 2001, 2004; Mehlen et al., 1996). αA-crystallin could bind proapoptotic factors p53, Bax, and Bcl-X(S), preventing their translocation to the mitochondria to promote apoptosis (Kamradt and Chen, 2002; Liu et al., 2007). In addition, the removal of αA-crystallin caused a decrease in Bad S136 phosphorylation, a potent signal for apoptosis (Xi et al., 2008). It has been hypothesized that in response to environmental and metabolic stresses in the lens, αA-crystallin binds to both unfolded proteins and those involved in apoptosis, including cytochrome c and caspase-3 (Kamradt et al., 2005; McGreal et al., 2012). The antiapoptotic function of α-crystallin may also be important in other tissues. It has been shown that αA-crystallin was expressed in the brain in a highly region-specific manner and acted as an essential survival factor in dopaminergic olfactory bulb interneurons (Ninkovic et al., 2010).
In the ischemic heart, phosphorylated αB-crystallin was translocated to the mitochondria where it stabilized mitochondrial membrane potential and prevented apoptosis (Whittaker et al., 2009). α-crystallin has also been shown to protect glioblastoma and retinoblastoma cancer cells from apoptosis (Kase et al., 2009; Stegh et al., 2008). Overexpression of αB-crystallin could inhibit hyperglycemia-induced apoptosis of human umbilical vein endothelial cells (Nagaraj et al., 2005). While the delivery of exogenous recombinant α-crystallin to the tissues offers an intriguing approach to prevent protein aggregation disease, the uptake of α-crystallin into the cells at levels sufficient for efficacy is likely to be a challenge.
In summary, we demonsntrate that αA-crystallins could protect LEC-iPSC-like cells from apoptosis and oxidation stress. Considering that the dysregulation of developmental apoptosis due to genetic mutations or environmental factors causes microphthalmia or cataractogenesis, our findings help better understanding of the role of αA-crystallin in the early cataractogenesis.
Footnotes
Author Disclosure Statement
The authors declare no conflicting financial interests exist.