
Abstract
Abstract
Induced pluripotent stem cells (iPSCs) share many characteristics with embryonic stem cells, but lack ethical controversy. They provide vast opportunities for disease modeling, pathogenesis understanding, therapeutic drug development, toxicology, organ synthesis, and treatment of degenerative disease. However, this procedure also has many potential challenges, including a slow generation time, low efficiency, partially reprogrammed colonies, as well as somatic coding mutations in the genome. Pioneered by Shinya Yamanaka's team in 2006, iPSCs were first generated by introducing four transcription factors: Oct 4, Sox 2, Klf 4, and c-Myc (OSKM). Of those factors, Klf 4 and c-Myc are oncogenes, which are potentially a tumor risk. Therefore, to avoid problems such as tumorigenesis and low throughput, one of the key strategies has been to use other methods, including members of the same subgroup of transcription factors, activators or inhibitors of signaling pathways, microRNAs, epigenetic modifiers, or even differentiation-associated factors, to functionally replace the reprogramming transcription factors. In this study, we will mainly focus on the advances in the generation of iPSCs with substitutes for OSKM. The identification and combination of novel proteins or chemicals, particularly small molecules, to induce pluripotency will provide useful tools to discover the molecular mechanisms governing reprogramming and ultimately lead to the development of new iPSC-based therapeutics for future clinical applications.
Introduction
I
However it remains controversial whether iPSCs are molecularly and functionally equivalent to blastocyst-derived ESCs (Stadtfeld et al., 2010). The differentiation potential of iPSCs generated by forced expression of defined sets of transcription factors in somatic cells is limited as most have failed to support the development of entirely iPSC-derived animals (“all-iPSC animals”) using the tetraploid complementation assay. All iPSC animals and its offspring have potentially increased incidences of tumors. Therefore, scientists attempt to explore new substitutes, especially small molecules to replace all Yamanaka's four transcription factors OSKM, which will hopefully establish new reprogramming methods with higher safety and availability for clinical translation (Xu et al., 2015).
Substitutes for Oct 4
Oct 4, octamer-binding transcription factor 4, also known as Pou5fl, belongs to the octamer-binding subgroup of the POU family of transcription factors. It was found to be expressed in the oocytes, early embryos, inner cell masses (ICM), germ cells, and PSCs, including ESCs, embryonic germ cells (EGCs), and embryonic carcinoma cells (ECCs). Oct 4 was considered to be the genetic “master switch” in the establishment of totipotency–pluripotency during the life cycle of mammals (Pesce et al., 1999), and presumed to be the most upstream gene in the molecular circuitry of pluripotency (Jaenisch and Young, 2008). The combination of this single transcription factor and chemical compounds was found to be sufficient to reprogram somatic cells into iPSCs (Li et al., 2011; Zhu et al., 2010). It was once thought to be the only factor that could not be substituted in Yamanaka's 4-factor reprogramming process by other substances, including its similar homologs Oct 1 and Oct 6 (Jerabek et al., 2014; Nakagawa et al., 2008).
However, a recently study indicated that maternal Oct 4A was crucial for neither the establishment of totipotency in embryos nor the induction of pluripotency in somatic cells using oocytes, Oct 4A-deficient oocytes were able to reprogram fibroblasts into pluripotent cells (Wu et al., 2013). The initial stages of reprogramming may also proceed by an alternative, Oct 4-independent mechanism, allowing Oct 4 to be omitted from the initial reprogramming cocktail. It is of great significance to find substitutes for Oct 4 to elucidate its physiological roles and gain a better understanding of the reprogramming mechanisms, which remain largely unknown.
Inhibitor of transforming growth factor beta signaling pathway: SB431542
The signaling pathways orchestrated by leukemia inhibitor factor (LIF)/signal transducer and activator of transcription 3 (STAT3), Wnt/β-catenin, and fibroblast growth factor (FGF)/mitogen-activated protein kinase kinase (MEK)/extracellular signal-regulated kinase (ERK) play key roles in the generation of pluripotency. The transforming growth factor beta (TGF-β) signaling pathway also contributes to the regulation of many cellular processes, including cell growth, cell differentiation, development, apoptosis, and cellular homeostasis in a wide range of biological systems. Inhibition of TGF-β signaling cooperates in the reprogramming of murine fibroblasts by enabling faster, more efficient induction of iPSCs, whereas activation of TGF-β signaling blocks reprogramming. Inhibition of TGF-β signaling using SB431542 can substitute for Oct 4 in the generation of iPSCs and sustain the pluripotency of iPSCs through modulating FGF/MEK/ERK signaling (Tan et al., 2015).
Differentiation-associated factors: GATA transcription factors and PAX1
According to the prevailing opinion, pluripotency-associated factors and their rivals, lineage specifiers, determine the identities of pluripotent and differentiated cells, respectively. However, an alternative view of pluripotency is that rather than being an intrinsically stable “ground state,” pluripotency is an inherently precarious condition in which rival lineage specifiers continually compete to specify differentiation along mutually exclusive lineages (Loh and Lim, 2011).
Shu et al. provided experimental and computational supports for this idea, which revealed that mouse fibroblasts could be reprogrammed into iPSCs using lineage specifiers, including GATA3, GATA6, and PAX1. This combination of lineage specifiers, which are involved in mesendodermal lineage specification, did not include any of the core pluripotency factors, while effectively replacing Oct 4. Based on these results, Shu et al. proposed a “seesaw model” in which a balance was established using pluripotency factors and/or counteracting lineage specifiers that facilitated reprogramming in somatic cells. The pluripotent state could be reached only if all specification forces were well balanced (Shu et al., 2013). Recent data indicated that all six members of the GATA transcription factor family (including GATA1, GATA2, GATA3, GATA4, GATA5, and GATA6) could inhibit the overrepresented ectodermal lineage genes (such as Dlx3 and Lhx5), substitute for Oct 4, and reprogram mouse somatic cells to a pluripotent state. The GATA family may serve as an important mediator of cell fate conversion (Shu et al., 2015). These results change the traditional perspective that the conversion of cell fate requires induction by the high expression of the Yamanaka's factors.
Therefore, screening for novel Oct 4 substitutes should not be limited to pluripotency-associated factors. This new method will cast light on the molecular mechanisms that underlie reprogramming and cell fate determination, and hopefully facilitate the development of safer and more efficient reprogramming strategies (Ben-David et al., 2013).
Nuclear receptors: Nr5a1 and Nr5a2
Several nuclear receptors (NRs), including estrogen-related receptor beta (Esrrb) (Feng et al., 2009a; Katano et al., 2015), Dax1 (Uranishi et al., 2013; Zhang et al., 2014), nuclear receptor 5A (Nr5a) family (Lu et al., 2013), and retinoic acid receptors (Yang et al., 2015) have been shown to regulate the expression of pluripotency factors and play imperative roles in the maintenance of pluripotency. Nr5a family proteins, particularly its two members, steroidogenic factor-1 (SF-1 or Nr5a1) and liver receptor homolog-1 (LRH-1 or Nr5a2), have emerged as major players in the molecular network governing reprogramming in somatic cells (Gonzales and Ng, 2013). When overexpressed with Oct 4, Sox 2, and Klf 4 (OSK), either Nr5a2 or Nr5a1 was able to enhance the efficiency of iPSC derivation (Heng et al., 2010). Coexpressing retinoic acid receptor gamma and Nr5a2 with OSKM greatly accelerated the reprogramming process and readily reprogrammed primary human neonatal and adult fibroblast cells into iPSCs (Wang et al., 2011).
Nr5a2 and Nr5a1 also can substitute for Oct 4, and generate iPSCs with Sox 2, Klf 4, and c-Myc (SKM) from mouse embryonic fibroblasts (MEFs) (Heng et al., 2010; Rony et al., 2015), but this may be due to their ability to activate Oct 4 expression (Wagner et al., 2010). However, many other factors that strongly regulate Oct 4 expression were unable to successfully obtain iPSCs, suggesting that only upregulation of endogenous Oct 4 is not sufficient to drive reprogramming (Gonzales and Ng, 2013). Similar to other NRs, Nr5a2 possesses a DNA-binding domain (DBD) and a ligand-binding domain (LBD); the DBD is crucial for the reprogramming function of Nr5a2, whereas LBD is dispensable, and the orphan NR does not depend on any ligand to carry out its role in the reprogramming process (Heng et al., 2010).
Direct transfection with Nr5a1 or Nr5a2 was able to convert egg cylinder-derived epiblast stem cells (EpiSCs) to naive pluripotency, implying that their function specifically lies in naive pluripotency; Nr5a receptors are potent reprogramming factors in EpiSCs. Although sustaining the expression of Oct 4 may be part of the contribution of Nr5a receptors to reprogramming, it is not sufficient. Furthermore, unlike previously identified reprogramming factors, no evidence indicated that Nr5a receptors could sustain self-renewal of ESCs (Guo and Smith, 2010). Therefore, those results imply a different foundation for their capacity to reset pluripotency.
Pluripotency can be defined as two distinct phases: naive and primed. Naive (ground) pluripotency is established in the preimplantation epiblasts in vivo and can be captured in vitro in the form of mouse ESCs, naive PSCs are characterized by compact dome-like colonies, dependency on LIF, and having the capacity for producing chimeras with germline transmission; EpiSCs are derived from postimplantation epiblasts; they are also pluripotent, but they represent a primed pluripotency, a postimplantation pluripotent state prepared for lineage specification and commitment. Primed PSCs are characterized by flat colonies, dependency on basic FGF and activin instead of LIF, and having very limited capacity to contribute to chimeric offspring (Dodsworth et al., 2015; Fujishiro et al., 2013; Nichols and Smith, 2009; Van Bortle and Corces, 2014). ESCs can be induced to differentiate into EpiSCs by exposure to activin and FGF, but the reverse transition requires transfection with Klf 4 or inducement of small molecules (Guo et al., 2009a; Zhou et al., 2010).
Ten-eleven translocation proteins
DNA demethylation has been playing a prominent role in reactivating pluripotency loci, which are hypermethylated and silenced in somatic cells. It is likely that both active demethylation and replication-coupled passive demethylation mechanisms contribute to pluripotency gene demethylation for cell reprogramming (Hu et al., 2014). Ten-eleven translocation (Tet) proteins are α-ketoglutarate (α-KG)- and Fe (II)-dependent dioxygenases that catalyze the iterative oxidation of 5-methylcytosine (5mC) to 5-hydroxymethyl cytosine (5hmC), and its higher oxidative products 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Tet enzyme-driven 5mC oxidation provides a direct mechanistic route for both active and passive DNA demethylation (Hill et al., 2014; Nakagawa et al., 2015). Tet proteins can also physically interact with O-linked β-N-acetylglucosamine (O-GlcNAc) transferase (OGT), and mediate the recruitment of OGT to genomic loci to modify histones and regulate gene expression (Deplus et al., 2013; Vella et al., 2013). As they have the potential to switch on or off specific gene expression, it is not a surprise that they play a crucial role in epigenetic modifications and the induction of transcription at endogenous pluripotency loci such as Oct 4, Nanog, and Esrrb.
Depletion of Tet1 or Tet2 significantly reduced the reprogramming efficiency (Costa et al., 2013; Doege et al., 2012; Piccolo et al., 2013), Tet1, Tet2, and Tet3 triple-knockout MEFs could not be reprogrammed into iPSCs after transduction of the reprogramming factors OSK, because they failed to initiate the mesenchymal-to-epithelial transition (MET) process due to their inability to downregulate the key mesenchymal regulators (Hu et al., 2014). Gao et al. showed that DNA hydroxylase Tet1 acted at the Oct 4 loci to promote 5mC-to-5hmC conversion and facilitated iPSC induction by promoting Oct 4 demethylation and reactivation. Especially, interesting is the finding that Tet1 could substitute for Oct 4 in the initiation of somatic cell reprogramming in conjunction with SKM, partly by promoting the demethylation and reactivation of endogenous Oct 4. The TSKM 2° (a secondary reprogramming system) system, established using fibroblasts derived from full-term TSKM (Tet1 and SKM) mice, could initiate reprogramming rapidly and efficiently. The lower level of 5mC and higher level of 5hmC in TSKM iPSCs compared to OSKM iPSCs were more inclined to an ESC-like state. Furthermore, unlike the OSKM mice, all TSKM mice were free of tumors, which hints that TSKM-mediated reprogramming may be more beneficial to iPSC production due to this relatively lower risk of tumorigenesis (Gao et al., 2013).
Epithelial cadherin
Epithelial cadherin (E-cadherin), the prototype member of the classical cadherin family, is a Ca2+-dependent transmembrane glycoprotein that mediates specific cell-cell adhesion (Cavallaro and Christofori, 2004). It is also a master stem cell regulator that is required for both ESC maintenance and differentiation (Pieters and van Roy, 2014). Differentiation of ESCs has been associated with downregulation of E-cadherin (Soncin et al., 2009), while expression of E-cadherin was markedly higher in undifferentiated ESCs (Li et al., 2010b). E-cadherin promotes Klf 4 and Nanog transcription and protein expression in mouse ESCs through STAT3 phosphorylation (Hawkins et al., 2012).
E-cadherin knockout ESCs irreversibly lost most of the key pluripotency features and differentiation capacity, and no iPSCs were obtained from E-cadherin-depleted fibroblasts, showing that E-cadherin-mediated cell adhesion is essential for the generation of iPSCs (Bedzhov et al., 2013). Both knockdown and knockout studies provided functional evidence that E-cadherin is a crucial mediator of MET during early reprogramming (Chen et al., 2010). Overexpression of E-cadherin in combination with the cytokine LIF dramatically improved the efficiency of conversion of EpiSCs to naive-like PSCs (Murayama et al., 2015). Those results indicate that E-cadherin has a crucial role in the maintenance of ESC pluripotency and formation of iPSCs. Ectopic expression of E-cadherin can also replace the requirement for exogenous Oct 4 during reprogramming and generate iPSCs in combination with SKM. ESKM-reprogrammed iPSC clones produce endogenous Oct 4 and E-cadherin protein. Signaling events downstream from E-cadherin might provide a stimulus to upregulate endogenous Oct 4 during successful reprogramming (Redmer et al., 2011).
Nanog and BMI1/sonic hedgehog agonists
Nanog is composed of 305 amino acids, which contains an N-terminal domain (ND), a DNA-binding homeodomain (HD), and a C-terminal domain (CD). ND (Guo et al., 2009c) and CD (Wang et al., 2008) are transcriptional activation domains required for Nanog to mediate self-renewal of mouse ESCs. HD shares high similarity among species and is sufficient to induce naive pluripotency (Theunissen et al., 2011). Nanog is also expressed exclusively in the ICM, EGCs, ECCs, and germ cell tumors (Clark et al., 2004; Hart et al., 2004), but it is downregulated upon differentiation (Hyslop et al., 2005). Scientists found that Nanog was dispensable for the establishment and maintenance of primed pluripotency (pre-iPSCs) from somatic cells (Carter et al., 2014; Osorno et al., 2012), but it was very important for the acquisition of naive pluripotency (Silva et al., 2009).
Bmi1, B cell-specific Moloney murine leukemia virus integration site 1, is a polycomb group family member, which has been reported to regulate the proliferative activity of normal cells, stem cells, and progenitor cells, and plays an important role in cell cycle, cell immortalization, senescence, and cancer recurrence/chemoresistance (López-Arribillaga et al., 2015; Siddique and Saleem, 2012).
Oct 4 and Nanog are key regulators in the formation and maintenance of pluripotency (Mitsui et al., 2003). More than 90% of the gene promoter regions bound by Oct 4 and Sox 2 are also bound by Nanog (Boyer et al., 2005). MEFs could be reprogrammed into iPSCs by the transduction of Bmi1 and Nanog together, which replaced SKM, in the absence of Oct 4. The function of Bmi1 could be replaced by sonic hedgehog (Shh) agonists (oxysterol and purmophamine), iPSCs could also be generated from MEFs by Nanog and Shh agonists. Therefore, Nanog in combination with Bmi1 or Shh agonists is sufficient to replace Oct 4 for the reprogramming of MEFs into iPSCs (Moon et al., 2013). hNanog L122A, one functionally enhanced mutant of Nanog, enhances binding affinity for Oct 4 promoter DNA, self-renewal of mouse ESCs, and reprogramming of primed state pluripotency of human iPSCs into ground state pluripotency (Hayashi et al., 2015).
Substitutes for Sox 2
Sox 2 is a member of the sex-determining region of the Y chromosome-related high-mobility group box (Sox) family of transcription factors. It expresses during the preimplantation mammalian development with a spatiotemporal pattern and takes part in the regulation of development (Singha et al., 2015; Yu et al., 2015). Sox 2, a faithful pluripotent marker, plays key roles in maintaining the self-renewal and pluripotency of undifferentiated ESCs (Liu et al., 2015b). Reduction of Sox 2 expression results in loss of the undifferentiated stem cell state in human ESCs (Fong et al., 2008) and human PSCs (Wu et al., 2015a), and decreases the self-renewal of mouse ESCs (Singh et al., 2015). Overexpression of Sox 2 in human ESCs results in their differentiation to the trophectoderm lineage (Adachi et al., 2010) and promots neural and glandular epithelium development (Adachi et al., 2011). Sox 2 is also overexpressed in many different kinds of cancer cells, including glioma (Annovazzi et al., 2011), breast cancer (Leis et al., 2012), colorectal cancer (Fang et al., 2010), melanoma (Kasia, 2015), lung cancer (Karachaliou et al., 2013), and endometrial adenocarcinoma (Pityński et al., 2015).
Members of the Sox B1 subgroup: Sox 1 and Sox 3
Sox 1, Sox 2, and Sox 3 are the three members of the Sox B1 subgroup of transcription factors, each having similar sequences, expression patterns, and overexpression phenotypes (Archer et al., 2011); they play critical roles in embryogenesis and the control of cell fate decisions within the early developing nervous system. Sox 1 and Sox 3 are capable of replacing Sox 2 for the generation of iPSCs from mouse fibroblasts (Nakagawa et al., 2008). Similar to this report, individual overexpression of Sox 1 or Sox 3 alongside Oct 4, Klf 4, and c-Myc (OKM) is sufficient for the generation of human iPSCs with an efficiency of 0.01% and 0.004%, respectively. Sox 1 and Sox 3 overexpression in human fibroblasts resulted in the upregulation of endogenous Sox 2 expression. All of the iPSCs generated in the absence of Sox 2 demonstrated a gene expression profile closely resembling that of other PSC lines, including ESCs and OSKM-derived iPSCs (Montserrat et al., 2013). Therefore, Sox 1 and Sox 3 can functionally substitute for Sox 2 during reprogramming.
Inhibitor of TGF-β signaling pathway: RepSox
RepSox, also named E-616452 or activin-like receptor kinase (ALK5) inhibitor II, is a TGF-β receptor 1 kinase inhibitor. RepSox can enhance the efficiency and kinetics of reprogramming, efficiently replace either Sox 2 or c-Myc, but not both simultaneously, and reprogram mouse fibroblasts into iPSCs through the inhibition of TGF-β signaling in the presence of Oct 4 and Klf 4. Replacement of Sox 2 by RepSox does not result from the direct activation of endogenous Sox 2 or other closely related genes, but it increases the expression of Nanog (Ichida et al., 2009). These numerous differing actions suggest that the mechanism of action of RepSox is distinct from those of the OSKM reprogramming factors (Maherali and Hochedlinger, 2009).
Esrrb and nanog
Esrrb is an orphan NR that is part of the pluripotency gene regulatory network (Festuccia et al., 2013). It is a pivotal target of the glycogen synthase kinase-3 (Gsk3)/transcription factor 3 (Tcf3) axis regulating ESC self-renewal (Martello et al., 2012). Overexpression of Esrrb preserved the self-renewal and the naive pluripotent state of ESCs (Katano et al., 2015).
Esrrb was shown to mediate reprogramming of MEFs into iPSCs in conjunction with Oct 4 and Sox 2 through the upregulation of ESC-specific genes (Feng et al., 2009a). Ectopic expression of Esrrb, Sall 4, Nanog, and Lin28 in MEFs generated high-quality iPSCs more efficiently than other combinations of factors, including OSKM (Buganim et al., 2014). Esrrb is expressed in fully reprogrammed iPSCs, but not in partially reprogrammed colonies, making it a better predictive marker for reprogramming than traditional markers like Oct 4 and Fbxo15. Esrrb is paramount for reprogramming in the absence of exogenous Sox 2, probably through activation of endogenous Sox 2 expression. This occurs at a late phase in reprogramming that initiates a series of consecutive steps toward pluripotency. Esrrb can replace Sox 2 in the reprogramming cocktail when supplied with Nanog (Buganim et al., 2012). However, Feng et al. (2009a) found that in the absence of Sox 2, the number of GFP-positive colonies induced by Esrrb and OKM was markedly reduced, indicating that Esrrb is unable to replace Sox 2 in reprogramming MEFs.
Early ectodermal lineage specifiers: zinc finger nuclear protein (ZNF521/Zfp 521)
ZNF521 is a multifunctional cotranscription factor with 30 zinc finger domains and an aminoterminal transcriptional repressor motif that binds to the nucleosome remodeling and histone deacetylase (HDAC) complex (Mega et al., 2011). It is an early neural nuclear protein with a potent neural differentiation-promoting activity. Forced expression of ZNF521 induced the neural conversion of ESCs even in the presence of the neural differentiation inhibitor bone morphogenetic protein 4 (BMP4) (Kamiya et al., 2011). The decline of pluripotency gene expression coincided with the gradual expression of early neural markers, including ZNF521, which were induced at day 2–5 in the SMAD7-mediated RUES1 neural induction of human ESCs (Ozair et al., 2013).
However, overexpression of ZNF521 in human fibroblasts alongside OKM generated iPSCs expressing the hallmarks of pluripotency. ZNF521 could serve as a replacement for exogenous Sox 2; RNA analysis of ZNF521-infected fibroblasts confirmed the rapid upregulation of endogenous Sox 2 expression. ZNF521 and two other different genes related to ectodermal specification (OTX2 and PAX6) alongside Klf 4 and c-Myc expression possessed a sufficient counteracting force to balance the effect of GATA3-VP16 on mesendodermal specification, and resulted in the appearance of iPSCs in the absence of Oct 4 and Sox 2. Therefore, there is a significant overlap between the concepts “pluripotency factors” and “lineage specifiers” and individual factors can play multiple roles depending on specific circumstances (Montserrat et al., 2013).
RCOR 2
In mammalian genomes, RCOR family consists of RCOR 1, RCOR 2, and RCOR 3. RCOR 1 is ubiquitously expressed in somatic cells, whereas RCOR 2 and RCOR 3 are found to be specifically and highly expressed in ESCs and the testis, respectively. ESC-specific corepressor RCOR 2 forms a complex with the histone H3K4me1/2 demethylase lysine-specific demethylase 1 (LSD1) and plays important roles in regulating self-renewal and pluripotency of ESCs. Knockdown of RCOR 2 expression during iPSC induction severely reduced the iPSC generation. Upregulation of BMP4 expression together with impaired expression of TGF-β inhibitor Noggin and Chordin might be correlated with the downregulation of Sox 2 expression in RCOR 2 knockdown ESCs. Mouse and human iPSCs generated by the overexpression of RCOR 2, together with OKM, are pluripotent. Expression of RCOR 2 in somatic cells could activate endogenous Sox 2 expression. RCOR 2 is the first non-Sox family protein factor that can substitute for Sox 2 in the reprogramming of both mouse and human somatic cells into iPSCs (Yang et al., 2011a).
Geminin/GMNN
Geminin is a nuclear protein (McGarry and Kirschner, 1998) and highly expressed in oocytes, early embryonic cells (Yang et al., 2011b), ESCs (Tabrizi et al., 2013), late meiotic spermatocytes (Barry et al., 2012), and neural progenitors, but is downregulated upon neuronal differentiation (Spella et al., 2007). It plays an important role in cell cycle control, cellular proliferation, cell lineage commitment, and neural differentiation (Kroll, 2007). Geminin deficiency in mouse embryos results in developmental arrest at preimplantation stages upon failure to form the ICM (Gonzalez et al., 2006). ESCs could not self-renew after Geminin deletion and pluripotency cannot be induced in fibroblasts in the absence of Geminin, which was required in the late phase of pluripotency induction, similar to the endogenous Sox 2 gene. In Geminin-deficient cells, the downstream, stem cell-specific enhancer of Sox 2 becomes repressed by a prominent repressive histone mark (Tabrizi et al., 2013).
Geminin is involved in ectodermal lineage specification (Seo et al., 2005) and it might attenuate the upregulation of mesendodermal-related genes induced by OKM during the reprogramming process. iPSC colonies could be generated with OKM and Geminin. Geminin, which is generally considered an ectodermal specifying factor and a pluripotency rival, could substitute for Sox 2 to promote reprogramming because it hampers the elevation of mesoendodermal genes induced by Oct 4. Concurrent coexpression of GATA6 and Geminin could substitute for the two most critical pluripotency factors Oct 4 and Sox 2, and successfully reprogrammed mouse fibroblasts into iPSCs in the presence of both Klf 4 and c-Myc (Shu et al., 2013). Therefore, the balance is established by pluripotency factors and/or lineage specifiers and results in induction of pluripotency.
Pan-Src family kinase inhibitors
src family kinases (SFKs) are a class of proto-oncogene tyrosine kinases that include c-Src, Yes, Fyn, Fgr, Lck, Hck, Blk, Lyn, and Frk in mammals (Yeatman, 2004). Several members of the SFK family are required to regulate the self-renewal and differentiation of ESCs (Meyn et al., 2005). SFK signaling is also an important mediator of reprogramming. Activation of c-Src signaling promotes the differentiation of ESCs (Meyn and Smithgall, 2009) and potently inhibits reprogramming mediated by OSKM (Staerk et al., 2011). Staerk et al. discovered that the Pan-SFK inhibitors iPYrazine, dasatinib, and PP1 could replace Sox 2 during the reprogramming of MEFs into iPSCs. These SFK inhibitors gave rise to iPSC colonies with a similar efficiency to that of TGF-β inhibitors (SB431542 and LY-364947). The inhibition of SFK signaling and TGF-β signaling may converge on a similar mechanism: the function of Sox 2 can be replaced during direct reprogramming by activating Nanog expression (Staerk et al., 2011).
Acute loss of retinoblastoma tumor suppressor gene
In mammals, the retinoblastoma (Rb) family is composed of Rb, Rb-like proteins p107 (RBL1), and p130 (RBL2). Rb proteins play very important roles in metabolism, apoptotic response, cell cycle, chromatin remodeling, cellular proliferation, cellular differentiation, cellular dedifferentiation and transformation, and pluripotency (Faial, 2015; Sengupta and Henry, 2015). The cyclin E-cyclin-dependent kinase 2 (Cdk2) complex continuously phosphorylates Rb to release E2F, which constitutively expresses E2F target genes throughout the cell cycle, leading to unusually rapid proliferation without gap phases, resulting in tumor formation (Kanai et al., 2015).
iPSCs and ESCs share some similarities to cancer cells, including the capacity to bypass senescence and form tumors upon transplantation (Goding et al., 2014). Cellular reprogramming to iPSCs has uncovered unsuspected links between tumor suppressors and pluripotency factors. Vilas et al. found that partial depletion of Sox 2-positive cells reduced the pituitary tumor development initiated by Rb loss in vivo. An impaired repression of Sox 2 transcription after the deletion of Rb or p130 results in low, but reproducible levels of this pluripotency gene in primary cells, which is sufficient to allow cell reprogramming to iPSCs without the need for exogenous Sox 2 (Vilas et al., 2015). Kareta et al. discovered that Rb inactivation accelerates and enhances the reprogramming of differentiated fibroblasts to a pluripotent state, Rb normally restricts this process. Surprisingly, this effect was not due to changes in the cell cycle, but rather from interferences with direct binding of Rb to pluripotency-associated loci such as Oct 4 and Sox 2. This contributes to their silencing in differentiated cells and restricts their reexpression during the reprogramming process. Thus, acute loss of Rb can functionally replace Sox 2 during iPSC formation (Kareta et al., 2015).
Substitutes for Klf 4
Klf 4 is a member of the Klf family of evolutionarily conserved zinc finger transcription factors (Shields et al., 1996). It plays major roles in the regulation of proliferation, cytodifferentiation, apoptosis, maintenance of normal tissue homeostasis, embryo development, and somatic cell reprogramming. Klf 4 is a bifunctional transcription factor, which can act as a transcriptional activator or a repressor, depending on the target gene, but utilizes different mechanisms when doing so (Evans and Liu, 2008). It is also associated with both tumor suppression [such as in cervical cancer (Mao et al., 2015) and colorectal cancer (Lee et al., 2014)] and oncogenesis [such as in skin squamous cell carcinoma (Chen et al., 2008)] depending on the type of cancer involved. Due to its oncogenicity, it is a controversial factor in the generation of iPSCs and several reports have indicated that Klf 4 was not required for the process of reprogramming. Thus, replacement of Klf 4 is a critical step in the eventual application of iPSCs as a therapeutically viable strategy.
Klf 2 and Klf 5
In addition to Klf 4, two other Klf family members (Klf 2 and Klf 5) are also important mediators of embryonic development and ESC identity. Absence of Klf 2 is detrimental for ESC self-renewal under dual inhibition (2i) of prodifferentiation of MEK/ERK and Gsk3/Tcf3 pathway conditions (Yeo et al., 2014). Klf 5−/− embryos fail to develop beyond the blastocyst stage in vivo and fail to produce ESC lines in vitro (Ema et al., 2008). Depletion of Klf 2, Klf 4, and Klf 5 by RNA interference (RNAi) was shown to result in differentiation and the impaired self-renewal of mouse ESCs, whereas single knockdown of each gene did not lead to an apparent change of phenotype, suggesting a functional redundancy among Klf 2, Klf 4, and Klf 5 in the maintenance of self-renewal of ESCs. The depletion of one Klf can be compensated for by the binding of other Klfs to the common target sites (Jiang et al., 2008). Klf 2 and Klf 5 are also capable of replacing Klf 4 in the generation of iPSCs from mouse fibroblasts by retroviral transduction of Oct 3/4, Sox 2, and c-Myc (OSM) (Nakagawa et al., 2008).
Mitochondrial E3 ligase: March5
Ubiquitination is a highly important mechanism for posttranslational control of protein functionality. Membrane-associated ring finger (C3HC4) 5 (March5) has been recognized as an E3 ubiquitin ligase located in the mitochondria. It is involved in promoting ESC stemness. Knockdown of March5 resulted in the decreased expression of some pluripotency markers, strongly inhibiting the clonogenicity of ESCs and OSKM-induced iPSCs, while promoting the differentiation of PSCs.
Klf 4 can be replaced by March5 for the generation of iPSCs; OSM and March5 (OSMM)-derived colonies successfully expressed ESC marker genes, indicating that March5 and Klf 4 have at least partially overlapping functions in regulating pluripotency. However, these cells were not able to form three germ layers-containing teratomas, thereby March5 has only the ability to partially replace Klf 4 to induce reprogramming. As a direct transcriptional target of Klf 4, March5 interacts and ubiquitinates Prkar1a, a negative regulatory subunit of protein kinase A (PKA). This activates PKA, thereafter leading to the inhibition of ERK activity and promoting stem cell properties of mouse ESCs. Therefore, March5 regulates mouse ESC pluripotency through the Klf 4-March5-Prkar1a/PKA- ERK pathway (Gu et al., 2015).
Esrrb
Orphan NR Esrrb can reprogram MEFs in the presence of OSM. The number of iPSC colonies generated using Esrrb substituted for Klf 4 is ∼50% of that obtained by introduction of OSKM. Esrrb may upregulate endogenous Klf 4, but can also replace Klf 4 during reprogramming in the absence of c-Myc as cells reprogrammed from MEFs transfected with Oct 4, Sox 2, and Esrrb are genuinely pluripotent. Apart from Klf 4, Esrrb can functionally substitute for a deficiency of Klf 2 and Klf 5 in ESCs. Esrrb tends to colocalize with Klf 4, and possesses a high degree of similarity with the Klf 4 transcriptional regulatory circuits in ESCs (Feng et al., 2009a).
Bone morphogenetic proteins
Factor-driven reprogramming is a multistep process, generally including initiation, maturation, and stabilization phases. The initiation phase of fibroblast reprogramming is characterized by a coordinated MET. TGF-β is a potent inducer of epithelial to mesenchymal transition (EMT) in some cells, and inhibition of TGF-β can accelerate or improve the reprogramming by promoting MET. BMPs, members of the TGF-β superfamily, have been involved in MET by reversing TGF-β-mediated EMT and reported to enhance MET (Samavarchi-Tehrani et al., 2010). Klf 4 also participates in MET by upregulating genes that are important for epithelial development and activating robust expression of epithelial markers, such as E-cadherin, Occludin, Ep-CAM, and Krt 8 (Chen et al., 2011; Li et al., 2010a). When epithelial cells are used as donor cells, Klf 4 can be omitted from the reprogramming cocktail (Li et al., 2010a). Therefore, Klf 4 also mainly works at the initial phase of reprogramming to trigger MET, it can be functionally substituted by BMPs.
MEFs and tail tibial fibroblasts can be efficiently reprogrammed into iPSCs in the presence of single-factor Oct 4 and BMPs. The effect of BMPs is beneficial only in the absence of Klf 4, suggesting that BMPs are functionally redundant with Klf 4 on inducing MET, or Klf 4 can inhibit the function of BMPs (Chen et al., 2011). While BMPs don't increase the endogenous expression of other reprogramming factors including OSKM and Esrrb (Feng et al., 2009a) during reprogramming, it can replace Klf 4 functionally by inducing MET through a mechanism different from the activation of endogenous reprogramming factors (Chen et al., 2011).
Kenpaullone
Kenpaullone is a potent inhibitor of Cdk1/cyclin B, Cdk2/cyclin A, Cdk5/p35, and GSK-3β, and exhibits an inhibitory activity toward various other kinases at higher concentrations (Knockaert et al., 2002). It could increase the reprogramming efficiency of MEFs transduced with OSKM; furthermore, it was able to substitute for Klf 4 in the reprogramming of MEFs into iPSCs in the presence of OSM, but the reprogramming efficiency was 10-fold lower than that after transduction with a Klf 4 vector. The reprogramming kinetics of kenpaullone-treated iPSC formation was delayed, indicating that kenpaullone was less efficient in epigenetic reprogramming than Klf 4 and might not completely recapitulate the role of Klf 4. Kenpaullone did not directly increase the expression of endogenous Klf 4; CHIR99021 (an inhibitor of GSK-3β) and purvalanol A (an inhibitor of Cdks) were unable to replace Klf 4 in the reprogramming of MEFs. Therefore, kenpaullone functions do not result from its well-documented role as a GSK-3β or cell cycle inhibitor, but through an entirely novel mechanism (Lyssiotis et al., 2009).
Others
miR 302–367 could substitute for the two oncogenic factors Klf 4 and c-Myc in the original OSKM combination for the successful generation of human iPSCs (Deng et al., 2015). Valproic acid (VPA) was found to allow for 2-factor (Oct 4 and Sox 2) reprogramming of neonatal human foreskin or dermal fibroblasts in the absence of oncogenes c-Myc and Klf 4. However, the efficiency of reprogramming by Oct 4, Sox 2, and VPA was significantly lower (about 200-fold) than that by three factors OSK with VPA treatment (Huangfu et al., 2008b). Therefore, Klf 4, although dispensable for reprogramming, plays an important role in facilitating somatic cell reprogramming.
Substitutes for c-Myc
c-Myc is the product of the c-Myc proto-oncogene, which plays a crucial role in cell growth, cellular metabolism, proliferation, differentiation, cell adhesion, apoptosis, senescence, embryonic development, stem cell self-renewal, and somatic cell reprogramming (Boxer and Dang, 2001). However, mutations in the c-Myc gene have been shown to be a contributing factor in many kinds of cancers, including prostate (Sarveswaran et al., 2015), pancreatic (Shukla et al., 2015), breast (Wu et al., 2015b), lung (Pan et al., 2015), and colon (Kugimiya et al., 2015) carcinomas. Due to the potential oncogenicity of c-Myc, it is also a controversial transcription factor in iPSC generation (Guo et al., 2009b). Therefore, searching for other substances to replace c-Myc will be very important for generating safer iPSCs for future clinical applications.
L-Myc and N-Myc
The Myc proto-oncogene family consists of three members: c-Myc, N-Myc, and L-Myc. c-Myc functions as a “double-edged sword,” promoting both iPSC generation and tumorigenicity (Nakagawa et al., 2010), N-Myc and L-Myc can substitute for c-Myc during the generation of iPSCs (Blelloch et al., 2007; Nakagawa et al., 2008). N-Myc is similar to c-Myc regarding its length, domain structure, and frequent association with human cancers (Malynn et al., 2000). In contrast, the L-Myc protein is shorter in N-terminal amino acid sequence, which contributes to the transformation activity than the other two members, and thus possesses a significantly lower transformation activity in cultured cells (Cole and Cowling, 2008). Consistent with this property, the aberrant expression of L-Myc in human cancers is less frequently reported (Okita and Yamanaka, 2011).
L-Myc can efficiently generate both mouse and human iPSCs (Drozd et al., 2015; Liu et al., 2015a; Nakagawa et al., 2010), and was more effective than c-Myc or N-Myc in the human fibroblasts (Okita and Yamanaka, 2011). Importantly, tumorigenicity is hardly observed in chimeric mice produced with L-Myc-induced iPSCs; L-Myc is the least tumorigenic of the three paralogs. Therefore, the tumorigenicity of c-Myc-induced iPSCs can be solved by using L-Myc instead of c-Myc for the iPSC generation. The iPSCs induced with L-Myc are safer, of high quality, and therefore are considered to be useful for future clinical applications (Nakagawa and Yamanaka, 2012).
Signaling pathways
Inhibiting TGF-β signaling
Repsox, also known as E-616452, is a TGF-β signaling inhibitor. It could enhance the number of iPSC colonies by 20-fold in the absence of c-Myc and resulted in a significant upregulation of the Wnt, Notch, and Hedgehog pathways, as well as c-Myc, which is a known target of the Wnt pathway (Zhang et al., 2012). These changes are contributing factors to the increase of reprogramming efficiency. Therefore, c-Myc can be omitted in the reprogramming cocktail when RepSox is used (Larsen, 2013). In addition, a structurally distinct TGF-β inhibitor SB431542 and a TGF-β-specific neutralizing antibody both increased the reprogramming efficiency in the absence of c-Myc. Therefore, the inhibition of TGF-β signaling sufficiently substitutes for c-Myc during cellular reprogramming (Ichida et al., 2009). Furthermore, iPSCs were generated from head-derived primary culture of mouse embryonic cells using small chemical inhibitors of MEK (PD0325901) and TGF-β pathways (SB431542) without delivery of exogenous transcription factors (Vrbsky et al., 2015).
Wnt signaling pathway regulators
The Wnt signaling pathway is intimately connected to the core transcriptional regulatory circuitry of pluripotency and plays a significant role in self-renewal of ESCs and cellular reprogramming (Fritz et al., 2015; ten Berge et al., 2011). Soluble Wnt3a could promote the generation of iPSCs from murine fibroblasts in the absence of c-Myc retrovirus. Wnt could serve to activate endogenous c-Myc directly, thereby substituting for exogenous c-Myc. Wnt signaling and c-Myc have overlapping, but not completely redundant roles in the induction of pluripotency (Marson et al., 2008). Therefore, signal transduction pathways and transcription factors can act coordinately to reprogram differentiated cells to a pluripotent state.
The CHIR99021 molecule, an inhibitor of GSK-3β, could strongly enhance the induction of pluripotency (Bar-Nur et al., 2014) and transit “pre-iPSCs” into fully reprogrammed pluripotent cells (Silva et al., 2008). CHIR99021 could induce the reprogramming of MEFs and human primary keratinocyte into iPSCs by overexpressing Oct 4 and Klf 4, thereby eliminating the use of c-Myc (Li et al., 2009). Also more dramatically, the combination of CHIR99021, Repsox, DZNep, and Forskolin was able to induce iPSCs from both MEFs and mouse adult fibroblasts, and replace OSKM (Hou et al., 2013). It reveals that the endogenous pluripotency program can be established using only small molecules rather than by exogenously provided “master genes.”
MicroRNAs
MicroRNAs (miRNAs) are small, single-stranded, and noncoding RNAs with an average length of ∼22 nucleotides, which modulate a large number of cell functions, including those related to stemness maintenance, differentiation, cell cycle progression, and cellular reprogramming (Bartel, 2004; Liao et al., 2011). Elucidation of targets of key miRNAs that interfere with the efficacy of iPSC generation might uncover new molecular mechanisms underlying cellular reprogramming.
miR-290 cluster
A subset of the miR-290 cluster (includes miR-291-3p, miR-294, and miR-295) is highly expressed in ESCs and rapidly downregulated upon differentiation (Chen et al., 2007). These miRNAs maintain the quick proliferative state of ESCs by accelerating the G1 to S phase transition (Wang et al., 2013) and enhance the efficiency of OSK-induced pluripotency. miR-294 can substitute for c-Myc in the generation of iPSCs, but does not further enhance c-Myc's contribution to reprogramming efficiency. Both c-Myc and n-Myc bind to the promoter region of the miR-290 cluster, suggesting that these miRNAs are downstream of c-Myc. This binding induces the direct expression of the miR-290 cluster, but requires epigenetic remodeling before they can be expressed (Cao et al., 2015). Unlike c-Myc, miR-294 did not induce proliferation of MEFs during the early phase of dedifferentiation and was not involved in MET during OSKM-based iPSC generation (Judson et al., 2009). In the early phase, its genomic region is epigenetically nonpermissive and thus remains silent. Instead, miR-294 is highly induced by c-Myc during the late phase of reprogramming (Moradi et al., 2014).
miR-302/367 cluster
The miR-302/367 cluster is a polycistronic miRNA, including five mature miRNAs (miR-302 b/c/a/d and miR-367). Reprogramming of human foreskin fibroblasts deficient in the miR-302/367 cluster was dramatically impaired and was largely rescued by the overexpression of an exogenous miR-302/367 cluster (Zhang et al., 2013b). The miR-302/367 cluster can substitute for c-Myc during the generation of mouse and human iPSCs induced by OSK. Further pathway analysis indicated that the miRNA-302/367 cluster increased iPSC generation efficiency, in part, by targeting TGF-β receptor 2, repressing TGF-β signaling, promoting E-cadherin expression, and accelerating the MET, which is a critical step during iPSC formation (Liao et al., 2011; Zhang et al., 2013b).
The expression of the miR-302/367 cluster rapidly and efficiently reprogrammed mouse and human somatic cells to an iPSC state without a requirement for exogenous transcription factors; miRNA and Hdac-mediated pathways could cooperate in a powerful way to reprogram somatic cells to pluripotency (Anokye-Danso et al., 2011). On the superiority of nanoparticulate gene delivery systems and miRNA for cellular reprogramming, miR-302/367 was used to replace the two oncogenic factors Klf 4 and c-Myc and reprogram human umbilical cord mesenchymal stem cells into a pluripotent state (Deng et al., 2015), which provides a novel, simple, safe, and effective alternative for generating iPSC-suitable clinical application.
miR-130/301/721
Nucleotides 2 through 7 in miR-130/301/721 are identical, which are important for target recognition within the 3′-UTR, and enhances the efficiency of iPSC generation from OSK-transduced MEFs in the absence of c-Myc. Meox2 plays a regulatory role in cell cycle progression by directly activating the tumor suppressor p21 (Chen et al., 2007). Overexpression of Meox 2 could significantly block the enhancing effects on reprogramming efficacy mediated by miR-130b/301b/721, however, the number of iPSC colonies in OSK-transduced MEFs could be increased after downregulating Meox2; miRNA-resistant Meox2 overexpression abolished the effect of miR-130/301/721 on iPSC formation. Although it is unclear how Meox2 interferes with reprogramming, Meox2 might inhibit TGF-β signaling or help overcome p21-mediated cell cycle arrest (Pfaff et al., 2011), which was described as a roadblock for iPSC generation (Hong et al., 2009).
Epigenetic modifiers
Epigenetic modifiers are used most commonly in various iPSC generation protocols because the epigenetic changes (e.g., DNA demethylation and histone acetylation) of pluripotency genes are the key features of reprogramming.
HDAC inhibitors
VPA (Huangfu et al., 2008a), suberoylanilide hydroxamic acid (Pandian et al., 2011), trichostatin A (Huangfu et al., 2008a), and butyrate (Zhang and Wu, 2013a) are HDAC inhibitors, which can improve somatic cell reprogramming. Especially, VPA could increase the reprogramming efficiency of OSKM-infected MEFs by more than 100-fold over the control, it also could efficiently induce the generation of iPSCs from MEFs (Huangfu et al., 2008a) and primary human fibroblasts (Huangfu et al., 2008b) infected with OSK, without introduction of c-Myc.
The effect of VPA on reprogramming may be due to the collective effects of upregulation of ESC-specific genes, downregulation of MEF-specific genes, and promotion of epigenetic remodeling. Chromatin modification is a key step in reprogramming fibroblasts into pluripotent cells (Huangfu et al., 2008a). VPA treatment before the introduction of exogenous Oct 4 was more effective and significantly increased the expression of endogenous Oct 4, Nanog, Klf 4, c-Myc (p < 0.01), Pax6, and Sox 1 (p < 0.001). VPA could be the best enhancer of pluripotency especially when applied before pluripotency induction by Oct 4 (Asadi et al., 2015). In addition, VPA is a US Food and Drug Administration-approved treatment for epilepsy (Leeman and Cole, 2008) and may therefore be easily applied in clinical medicine.
Histone methyltransferase inhibitors
iPSCs could be generated from primary fetal neural progenitor cells (NPCs) by Oct 3/4-Klf 4 viral transduction with simultaneous BIX-01294 (BIX) treatment, a histone methyltransferase (HMT) G9a inhibitor, BIX, could functionally replace Sox 2 and c-Myc during the reprogramming of NPCs, and meanwhile improve the reprogramming efficiency ∼10-fold over controls and ∼50% over OSKM-reprogrammed cells (Shi et al., 2008b). BIX had also been shown to be capable of reprogramming MEFs into iPSCs in combination with Oct 4 and Klf 4, as well as the small molecule BayK8644 with approximately half the efficiency of OSKM reprogramming (Shi et al., 2008a).
Histone H3K4 demethylation inhibitors
Tranylcypromine (TCP), a monoamine oxidase inhibitor used as an antidepressant drug, showed a potent effect on inhibiting the H3K4 demethylation and significantly facilitated iPSC generation induced by OSK in the absence of c-Myc (Li et al., 2011). When TCP was combined with CHIR99021, human neonatal keratinocytes were also reprogrammed into iPSCs by transduction of only Oct 4 and Klf 4 (Li et al., 2009). A combination of the small molecules TCP, VPA, CHIR99021, and VC6T was sufficient to induce reprogramming of mouse fibroblasts with only Oct 4, thus replacing SKM (Li et al., 2011). TCP was also reported to activate endogenous Pou5f1/Oct 4 expression and inhibit demethylation in P19 ECCs in vivo, which resulted in transcriptional derepression of Oct 4 and global increase in H3K4 methylation (Lee et al., 2006). Therefore, H3K4 demethylation can be a critical epigenetic barrier to reprogramming, which may repress the establishment of a pluripotency transcriptional network.
DNA methyltransferase inhibitors
Establishment of an “open” chromatin state involved in direct reprogramming to a pluripotent state correlates with DNA demethylation (Mikkelsen et al., 2008). DNA methyltransferase (DNMT) inhibitors are able to reactivate epigenetically silenced genes. Therefore, treatment with DNMT inhibitors can improve the overall efficiency of the reprogramming process. 5′-azacytidine (5′-aza-dC) is a DNMT inhibitor and treatment with it during days 8–10 resulted in a 4-fold increase in the number of ESC-like colonies from chimaera-derived Nanog-GFP MEFs being affected with OSKM over untreated controls; it induced a rapid and stable transition from a partially reprogrammed state to a fully reprogrammed iPSC state (Mikkelsen et al., 2008). 5′-aza-dC and OSK reprogrammed MEFs into iPSCs without the need for c-Myc, and improved the efficiency of iPSC colony formation by 3-fold at 10 days postinfection (Huangfu et al., 2008a). Full reprogramming involves upregulation of proliferative genes with functions such as DNA replication and cell cycle progression, which may be consistent with the expression of c-Myc (Adhikary and Eilers, 2005).
RG108 is also a DNMT inhibitor that enhanced reprogramming of Oct 4 and Klf 4-transduced MEFs in the presence of BIX (Shi et al., 2008a). Mouse skeletal myoblasts (SMs) also could be efficiently reprogrammed into iPSCs with RG108 by induction of Oct 3/4, RG108 treatment of SMs not only upregulated Oct 3/4 but also increased the expression of other already present factors, including SKM (Pasha et al., 2011). Therefore, 5′-aza-dC or RG108 can functionally replace c-Myc and support efficient reprogramming.
Gli-like transcription factor Glis1 (Glis family zinc finger 1)
Glis1 is enriched in unfertilized oocytes and one-cell-stage embryos. Glis1 markedly increased the number of iPSC colonies when it was cointroduced with OSK into fibroblasts. This effect of Glis1 was comparable to that of Myc; furthermore, Glis1 could counteract the undesired effect of Myc, which increased the number of partially reprogrammed cell colonies or transformed cell colonies. Glis1 promotes iPSC generation by activating multiple proreprogramming pathways; the expressions of multiple genes (including Esrrb, Wnt3, Wnt6, Wnt8a, Wnt10a, Lin28a, Nanog, Mycn, Mycl1, and Foxa2) whose products have been shown to enhance iPSC generation were markedly increased.
However, the expression of Myc was suppressed by Glis1 (Maekawa et al., 2011). N-Myc and L-Myc predominantly increased the numbers of ESC-like colonies (Nakagawa et al., 2010). N-Myc, L-Myc, and c-Myc are direct transcriptional targets of Glis1. The altered balance between N-Myc/L-Myc and c-Myc should contribute, at least in part, to the specific promotion of iPSC generation by Glis1. Endogenous Glis1 may have a supportive role during the generation of iPSCs by OSK. Glis1 can associate with OSK by a protein-protein interaction and thereby may promote the activation of OSK target genes (Maekawa et al., 2011).
Poly (ADP-ribose) polymerase 1
Poly (ADP-ribose) polymerase 1 (Parp1), a member of the Parp family of proteins, is a highly conserved DNA-binding protein that is abundant and ubiquitous in the nucleus. It catalyzes a process called poly (ADP-ribosylation) (PARylation). Parp1 and PARylation participate in the establishment and maintenance of a genome methylation pattern (Caiafa et al., 2009). Their activities were detected in iPSCs and ESCs at very high levels, but they were lower in differentiated cells (Chiou et al., 2013).
Furthermore, Parp1-PARylated proteins interacted significantly with Oct 4, Nanog, c-Myc, Klf 4, CTNNB1, WDR5, SUZ12, EZH2, DNMT3A/B, and JARID2 in the core network of nuclear reprogramming and pluripotent status. Parp1 and PARylation could enhance the expression of Oct 4 and Nanog in OSK-transfected reprogramming cells. Parp1 was able to replace c-Myc or Klf 4 to increase the efficiency of iPSC generation from MEFs. c-Myc is a direct regulator of Parp1 and PARylation, endogenous c-Myc promotes reprogramming by its upstream regulation of Parp1 and subsequent PARylation (Chiou et al., 2013). Therefore, Parp1 and PARylation can effectively promote iPSC production and help to maintain a pluripotent state in the absence of c-Myc.
Cellular Reprogramming Without Transfection of Transcription Factor Genes
To facilitate the transition of iPSCs to clinical practice, a variety of substances such as small molecules (Hou et al., 2013; Kimura et al., 2015), mRNA (El-Sayed et al., 2014; Liu and Verma, 2015), miRNAs (Miyoshi et al., 2011; Sandmaier and TeluGu, 2015), and proteins (Kim et al., 2009; Nemes et al., 2014; Zhou et al., 2009) have been developed for transgene-free reprogramming of pluripotency. Small molecules have advantages in reprogramming because they can be cell permeable, nonimmunogenic, controllable, more cost-effective, and more easily synthesized, preserved, and standardized, and they have the potential to steer reprogramming toward a faster, safer, more efficient, and directed process.
In mRNA reprogramming, iPSCs could be generated only using synthetic modified mRNA encoding the reprogramming factors; notable advantages of this approach include high efficiency, rapid kinetics, a very low aneuploidy rate, a low requirement of donor cell, a complete absence of integration, and obviation of a clean-up phase to purge the vector. However, due to the very short half-life of mRNAs, repeated transfections are required to induce iPSCs, so this method is relatively laborious.
miRNAs bind target mRNAs through complete or incomplete sequence complementarity and play a crucial role in the posttranscriptional regulation of gene expression, but without risk of genome integration. Specific miRNA mimics or miRNA inhibitors can completely replace exogenous transcription factors in the reprogramming. miRNAs remain stable for several days and function longer than the coding RNAs, however, attributing to the transient action, multiple transfections are also required for complete reprogramming.
Another way to avoid introducing any exogenous genetic modifications to target cells would be to deliver the reprogramming proteins directly into cells by conjugating them with a short peptide, such as polyarginine and human immunodeficiency virus-1 transcription activator (HIV-1 TAT). This method provides a simpler strategy for generating safer iPSCs and does not require repeated transfer or prolonged exposure to reprogramming materials; it also maintains genomic integrity better than viral reprogramming methods (Park et al., 2014). However, the main difficulty is how to transport proteins across cytomembrane, thus initiating reprogramming procedures and increasing its low reprogramming efficiency.
Generally speaking, each transcription factor of OSKM is not indispensable to generate iPSCs from somatic cells, and it can be replaced by members of the same subgroup of transcription factors, activator or inhibitor of signaling pathways, miRNAs, epigenetic modifiers, or even differentiation-associated factors (Fig. 1). Generation of transgene-free and mutation-free iPSCs is critical for future clinical applications. The screening and identification of substitutes for Yamanaka's four transcription factors will be beneficial for us to develop many novel, safe, and more efficient cocktails of chemicals or/and protein factors to generate fully reprogrammed iPSCs.
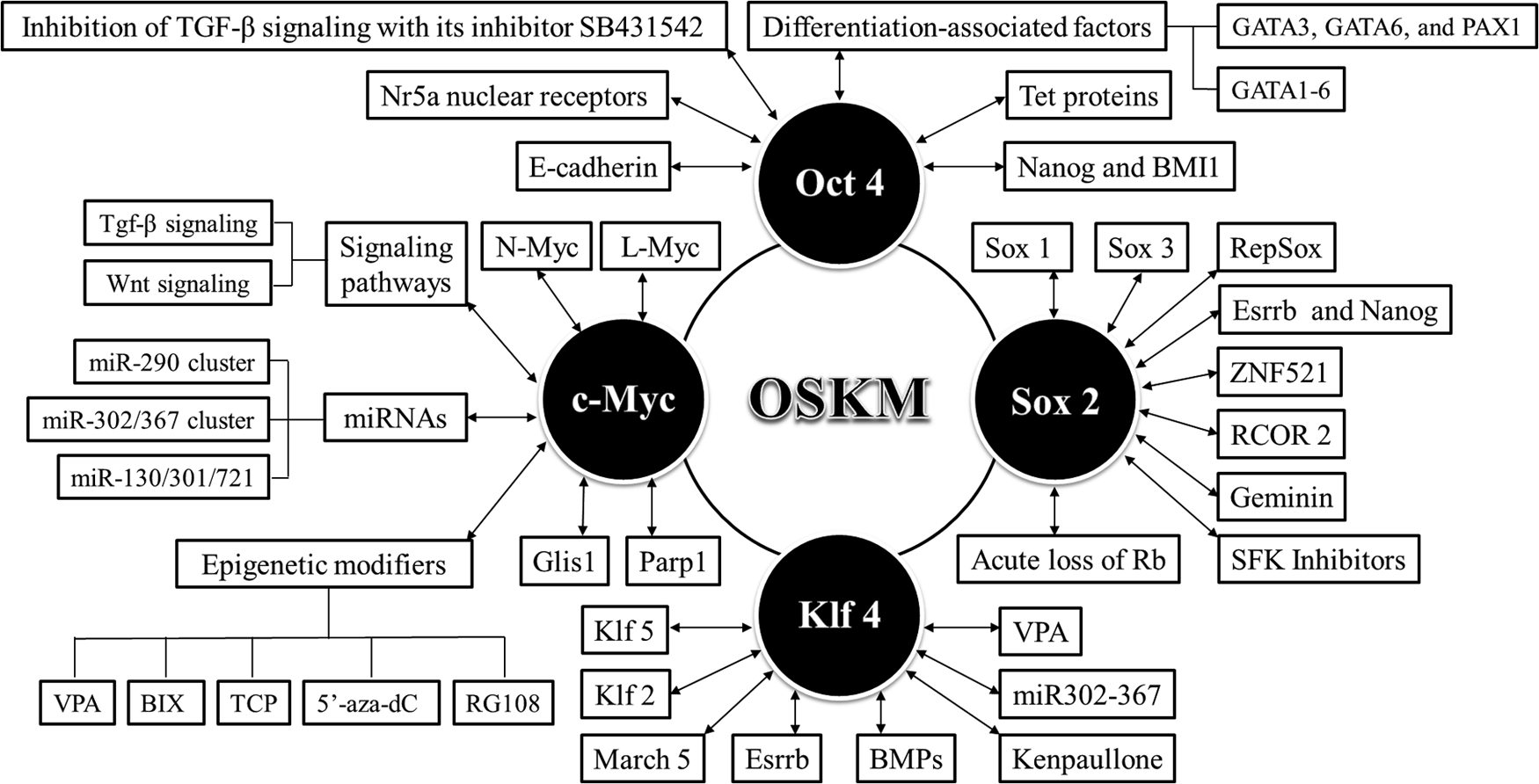
Substitutes for Yamanaka's four transcription factors (OSKM) during the generation of iPSCs. iPSCs, induced pluripotent stem cells.
Footnotes
Acknowledgments
This contribution is funded by the National Natural Science Foundation of China (General Program) (31572488), the Based and Advanced Research Projects of Chongqing, China (cstc 2013jcyjA10110), the Fundamental Research Funds for the Central Universities, China (XDJK2014C022), the PhD. Programs Foundation of Southwest University, China (SWU113002), and the National Undergraduate Innovation and Entrepreneurship Training Programs (201510635015). We thank M.S. Edward Trope for proof reading.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.