
Abstract
Abstract
Nuclei from less differentiated stem cells yield high cloning efficiency. However, pluripotent stem cells are rather difficult to obtain from bovines. Skin-derived precursor (SKPs) cells exhibit a certain degree of pluripotency, which has been shown to enhance the efficiency of nuclear transfer (NT) in pigs. In this study, bovine SKPs were isolated and characterized. Results showed that bovine SKPs expressed nestin, fibronectin, vimentin, pluripotency-related genes, and characteristic neural crest markers, such as NGFR, PAX3, SOX9, SNAI2, and OCT4. Bovine SKPs and fibroblasts were used as NT donor cells to examine and compare the preimplantation developmental potential of reconstructed embryos after somatic cell nuclear transfer (SCNT). Bovine SKP-cloned embryos displayed higher developmental competence in terms of blastocyst formation rate and total cell number in blastocysts compared with the bovine embryonic fibroblast-cloned embryos. This study revealed that bovine SKPs may be considered excellent candidate nuclear donors for SCNT and may provide a promising platform for transgenic cattle generation.
Introduction
S
SKPs as a population of nonadherent cells are isolated from the dermis and then proliferate and self-renew as floating spheres in response to fibroblast growth factor 2 and epidermal growth factor (EGF) (Biernaskie et al., 2006). These cells share the characteristics of embryonic neural crest stem cells and cooperatively express pluripotency-related genes, including POU5F1, SOX2, NANOG, and STAT3, and neural crest marker genes, such as p75NTR, TWIST1, PAX3, SNAI2, SOX9, and SOX10 (Zhao et al., 2009).These cells also express nestin, fibronectin, and vimentin (Toma et al., 2005).
SKPs display multiple lineage potential and produce neural and mesodermal progenies in vitro; as such, SKPs exhibit properties distinct from other known precursors within the skin (Fernandes et al., 2008). SKPs also represent a highly accessible and potentially autologous source of adult precursors that can generate different functional cell types, such as glial cells, neurons, smooth muscle cells, and adipocytes; therefore, SKPs are a predominant autologous donor source for stem cell therapy (Bakhtiari et al., 2012; Gingras et al., 2007; Higashida et al., 2010; Lavoie et al., 2009; Toma et al., 2001). SKPs also differentiate into vascular and nerve cells, which are crucial components for wound repair. With easy accessibility and multipotency, SKPs have been proposed as potential therapeutic candidates for diabetic wound healing (Sato et al., 2015).
SKPs exhibit other differentiation potentials. For instance, SKPs can be differentiated into Schwann cells (SCs) (Biernaskie et al., 2006), which are most likely applied to clinical trials of SKP-SCs as a therapy for spinal cord injury (Sparling et al., 2015). Rodent SKPs can repair damaged muscles, bones, and nerves (Lavoie et al., 2009; Steinbach et al., 2011). Furthermore, SKPs can be directly differentiated into fibroblast-like cells, which may be a key fibroblast source for wound healing (Shu et al., 2014). Human SKPs can also undergo hepatic differentiation and differentiate into functional insulin-producing cells (De Kock et al., 2015). Therefore, SKPs may provide a safe cell source for future stem cell-based therapies (Mehrabi et al., 2015).
Fetal porcine skin-originated sphere (PSOS) cells are a viable alternative to fibroblasts as donor cells for NT and hence may improve the efficiency of NT in pigs (Zhu et al., 2004). PSOS cells are multipotent and capable of differentiating into cells of neuronal and mesenchymal lineages upon induced differentiation (Dyce et al., 2004). Before differentiation occurs, these cells express undifferentiated markers, including STAT3, Oct4, and Nestin; conversely, the expression of these markers is downregulated or absent in differentiated cells.
Previous studies suggested that the use of less differentiated donor cell types may improve the success rate of somatic cell nuclear transfer (SCNT) (Dinnyes et al., 2012; Faast et al., 2006; Jaenisch et al., 2002). However, stable embryonic stem (ES) cell lines have yet to be generated from bovine embryos. Although the induced pluripotent stem (iPS) cell technology has been used to generate bovine iPS cells with defining features similar to either human or mouse ES cells (Han et al., 2011), several important issues, including the low efficacy of generating fully reprogrammed bovine iPS cell lines and the safety of iPS cells, should be addressed. In our study, bovine SKPs were isolated from the skin and then characterized. The isolated cells expressed pluripotency markers and differentiated into ectodermal and mesodermal cells. After SCNT, embryos cloned from multipotent SKPs showed higher developmental competence than those generated from bovine embryonic fibroblasts (BEFs). Therefore, SKPs can be used to enhance the efficiency of bovine SCNT.
Materials and Methods
The whole experimental procedure was approved by the Animal Care Commission of the College of Veterinary Medicine, Northwest A&F University, P.R. China.
Cell isolation and culture
Bovine SKPs were isolated in accordance with a previously described method (Toma et al., 2001) with slight modifications. The fetal skin tissue was separated from the neck and back of a healthy 50-day-old fetus. The fetus was obtained through spontaneous abortion from a startled cow that was housed at our experiment center. The skin tissue was carefully dissected to avoid isolating other tissues, cut into 1-mm3 pieces, washed thrice with Ca2+/Mg2+-free phosphate-buffered saline (PBS), and digested with 0.1% trypsin for 40 minutes at 37°C, and 0.1% DNase was added and incubated for 3 minutes at room temperature. Trypsin was neutralized with 10 mL of neutralization solution DMEM/F12 (1:1; Invitrogen) and 10% fetal bovine serum (FBS; Hyclone, Logan, UT). The cells were washed thrice with a DMEM/F12 medium. Tissue pieces were washed twice with PBS and mechanically dissociated by vortexing and pipetting with 2 mL of the medium. The cell suspension was passed through a 40-μm strainer (Falcon), centrifuged at 1200 rpm for 5 minutes, and resuspended in a neurobasal medium (Cat. No. 21103049; Gibco) containing antibiotics, 20 ng/mL EGF (Sigma), 2% B-27 (Gibco), and 40 ng/mL bFGF (Sigma).
Cells were cultured in a 24-well ultra-low attachment cell culture plate (Cat. No. 3473; Corning) at 37°C in a 5% CO2 tissue culture incubator. To passage the floating clusters of cells, we centrifuged the medium containing spheres at 1200 rpm for 5 minutes, mechanically dissociated the pellet with a fire-polished Pasteur pipette, and reseeded the cells in fresh medium containing B-27, EGF, and bFGF. A volume equal to half of the initial culture medium was added to the dishes every 3 days, and the cells were passaged every 6–7 days. Primary bovine embryonic fibroblasts were obtained in accordance with the method described by Pan et al. (2015) and then cultured in DMEM/F12 plus10% FBS. BEFs were obtained from the same fetus in accordance with our previously described method (Yu et al., 2013).
Immunofluorescence staining
Before the cells were fixed, the SKP spheres or BEFs were plated onto 24-well Nunclon culture slides coated with poly-
The cells were washed thrice with PBS and incubated with Alexa Fluor 488 (donkey anti-mouse) or Alexa Fluor 546 (goat anti-rabbit)-conjugated secondary antibody (1:500; Chemicon) in the dark at room temperature for 2 hours. DNA was stained with DAPI for 5 minutes. BEFs served as negative controls and were similarly processed.
RNA extraction and quantitative real-time PCR (QRT-PCR)
Total RNA was extracted from a 60-mm dish of SKPs or BEFs using an RNA prep pure micro kit (Qiagen) in accordance with the manufacturer's instructions. First-strand cDNAs were prepared from 0.5 μg of RNA by using a PrimeScript RT reagent kit (TaKaRa, Dalian, China), and gene expression was analyzed. QRT-PCR was performed using SYBR Premix Ex Taq™ (TaKaRa) and a StepOnePlus Thermocycler (Applied Biosystems). The reactions were set in 25 μL reaction mixtures containing 12.5 μL of 1 × SYBR Premix Ex Taq (TaKaRa), 0.5 μL of sense primer (10 μM), 0.5 μL of antisense primer (10 μM), 11 μL of distilled water, and 0.5 μL of template (100 ng/μL). The reaction conditions were as follows: 95°C for 10 seconds, followed by 40 cycles at 95°C for 5 seconds, and 60°C for 30 seconds. The GAPDH gene was used as a reference.
For each sample, target and reference genes were amplified independently in the same plate and in the same experimental run in triplicate. The 2−ΔΔCT method was applied to quantify the relative mRNA levels. The BEF was used as the calibrator. In this test, real-time PCR assays were repeated thrice. Fold changes were then compared through one-way ANOVA, followed by the Newman–Keuls test. The primer sequences are described in Table 1.
Annealing temperature.
Forward primer.
Reverse primer.
PCR, polymerase chain reaction.
Western blot
Total H3K9me3 protein was collected by RIPA buffer containing protease inhibitors. The protein was separated by 10% SDS-PAGE and transferred onto a PVDF membrane (Millipore, Billerica, MA). The primary rabbit anti-H3 antibody (1: 1000, Cat. No. ab1791; Abcom) or primary rabbit anti-H3K9me3 antibody (1: 1000, Cat. No. ab8898; Abcom) was used to detect bovine H3 or H3K9me3. After incubating with horseradish peroxidase-conjugated goat anti-rabbit IgG (H + L) (1:1000) for 2 hours at RT, the antigen–antibody complexes were detected by using the Western Light kit (Advansta, Menlo Park).
In vitro differentiation
For differentiation, the spheres were collected and resuspended in neurobasal medium containing B-27, N2, and 10% FBS (Gibco). The spheres or the single cells that dissociated from the spheres were plated onto six-well Nunclon culture slides coated with poly-
Nuclear transfer
NT was performed in accordance with previously described methods (Wang et al., 2011b). Briefly, cumulus cells were denuded from matured oocytes by using 0.1% bovine testicular hyaluronidase in PBS. The selected oocytes were enucleated with a 20-μm (internal diameter) glass pipette for removing the first polar body and a small amount of surrounding cytoplasm. The result of enucleation was confirmed by Hoechst 33342 staining. BEFs and SKPs at passages 1–3 were used as donor cells for SCNT to investigate the development of cloned embryos in vitro.
A donor cell was injected into the perivitelline space of a successful enucleated oocyte, then a double closely spaced electrical pulse of 35 V for 10 μseconds was used for oocyte–cell fusion. Before activation, the reconstructed embryos were kept in mSOF containing 5 μg/mL cytochalasin B for 2 hours. Activation of all fused embryos was then performed in 5 μM ionomycin for 5 minutes, followed by 4 hours of exposure to 1.9 mM 6-dimethynopyridine in mSOF. The cloned embryos were cultured for 7 days to evaluate their developmental rate.
Counting of nuclei of cloned blastocysts
The counting assay was conducted in accordance with the methods described in our previous study (Wang et al., 2011a). The cloned embryos were cultured in vitro for 7 days, then 32 BEFs and 31 SKPs cloned blastocysts were collected. The number of blastocyst cells was estimated by counting the total number of nuclei stained with DAPI using Nikon NIS element D 8.0 software. Data were statistically analyzed through one-way ANOVA and LSD tests and reported as mean ± standard error (SPSS18.0; IBM Corporation, Somers, NY). p < 0.05 was considered statistically significant.
Results
Isolation and characterization of SKPs
Bovine SKPs and BEFs were successfully isolated from bovine fetal skin. The BEFs were explanted (Fig. 1A) and the SKP spheres of floating cells were formed within 5–7 days (Fig. 1B, C). Immunofluorescence analysis revealed that the SKP spheres were consistently positive for pluripotent markers, namely SOX2, NANOG, and OCT4. The SKP spheres also expressed the nestin marker for neural precursors, fibronectin protein produced by bone marrow mesenchymal stem cells, and vimentin. By contrast, BEFs expressed vimentin and fibronectin (Fig. 2). The gene expression levels of NGFR, SNAI2, PAX3, SOX9, SOX2, NANOG, and OCT4 were subjected to QRT-PCR analysis.
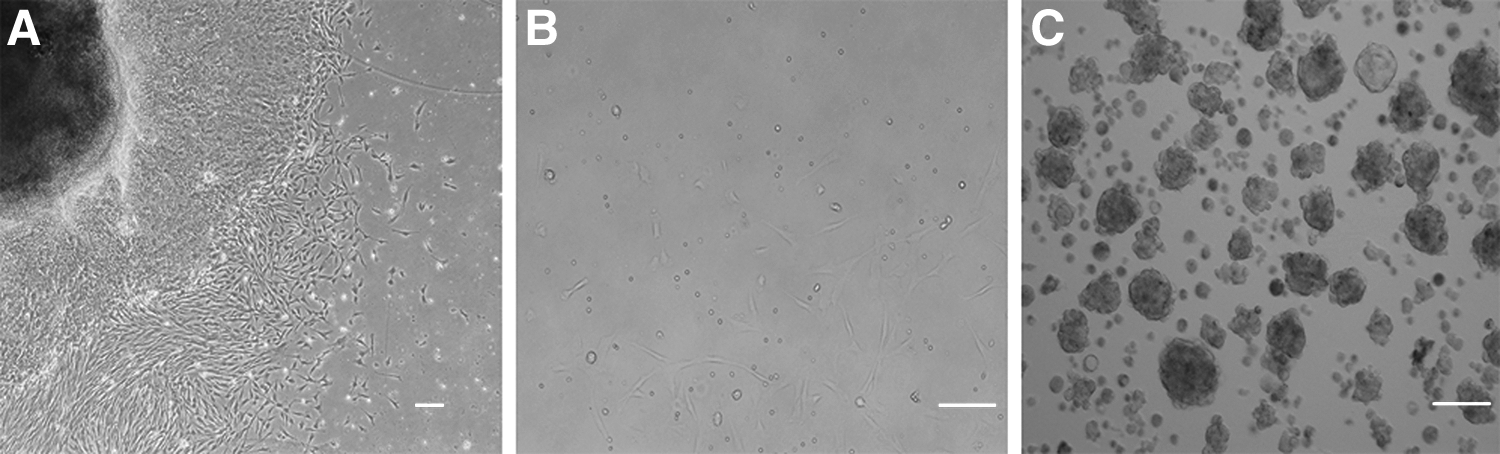
Culture of bovine SKPs and BEFs.

Immunofluorescence analysis of SKPs and BEFs. The expression of Vimentin, Fibronectin, Nestin, OCT4, SOX2, and NANOG was detected in SKPs and BEFs. The SKP spheres expressed Vimentin, Fibronectin, Nestin, OCT4, SOX2, and NANOG. BEFs were negative for Nestin, OCT4, SOX2, and NANOG. DNA was stained with DAPI.
The results revealed that SKPs could produce higher relative mRNA levels of neural crest stem cell markers (NGFR, SNAI2, PAX3, and SOX9) and pluripotent stem cell markers (SOX2, NANOG, and OCT4) compared with BEFs (p < 0.05) (Fig. 3A). H3K9me3 is considered one of the major barriers of reprogramming (Apostolou and Hochedlinger, 2013). Western blots demonstrated that the expression of H3K9me3 in SKPs was lower than that in BEFs (Fig. 3B). To confirm the undifferentiated state of SKPs, we tested the expression of the following lineage-related genes: GFAP for glial cells and tubulin β-III and MAP2 for neurons. Tubulin β-III was weakly expressed; MAP2 and GFAP did not obtain any detectable florescence (Fig. 4). These data suggested that SKPs may be more easily reprogrammed by NT.

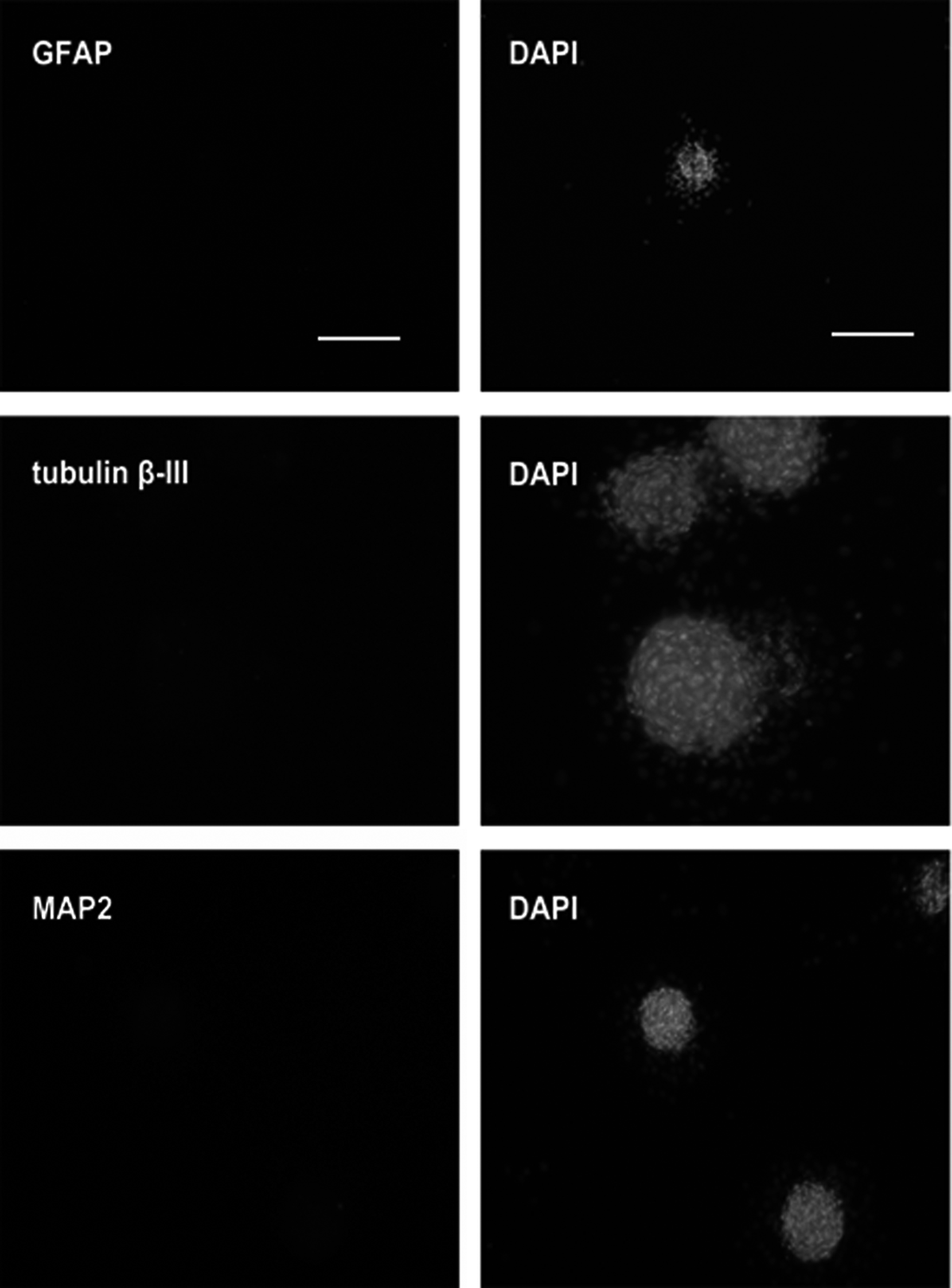
Immunofluorescence analysis of SKP clusters for GFAP, tubulin β-III, and MAP2. GFAP and tubulin β-III primary antibodies were detected with Alexa Fluor 546; MAP2 Alexa Fluor 488, DNA was stained with DAPI. Scale bar = 100 μm.
Differentiation potential of SKPs
The cell clusters at P3 were transferred to poly-
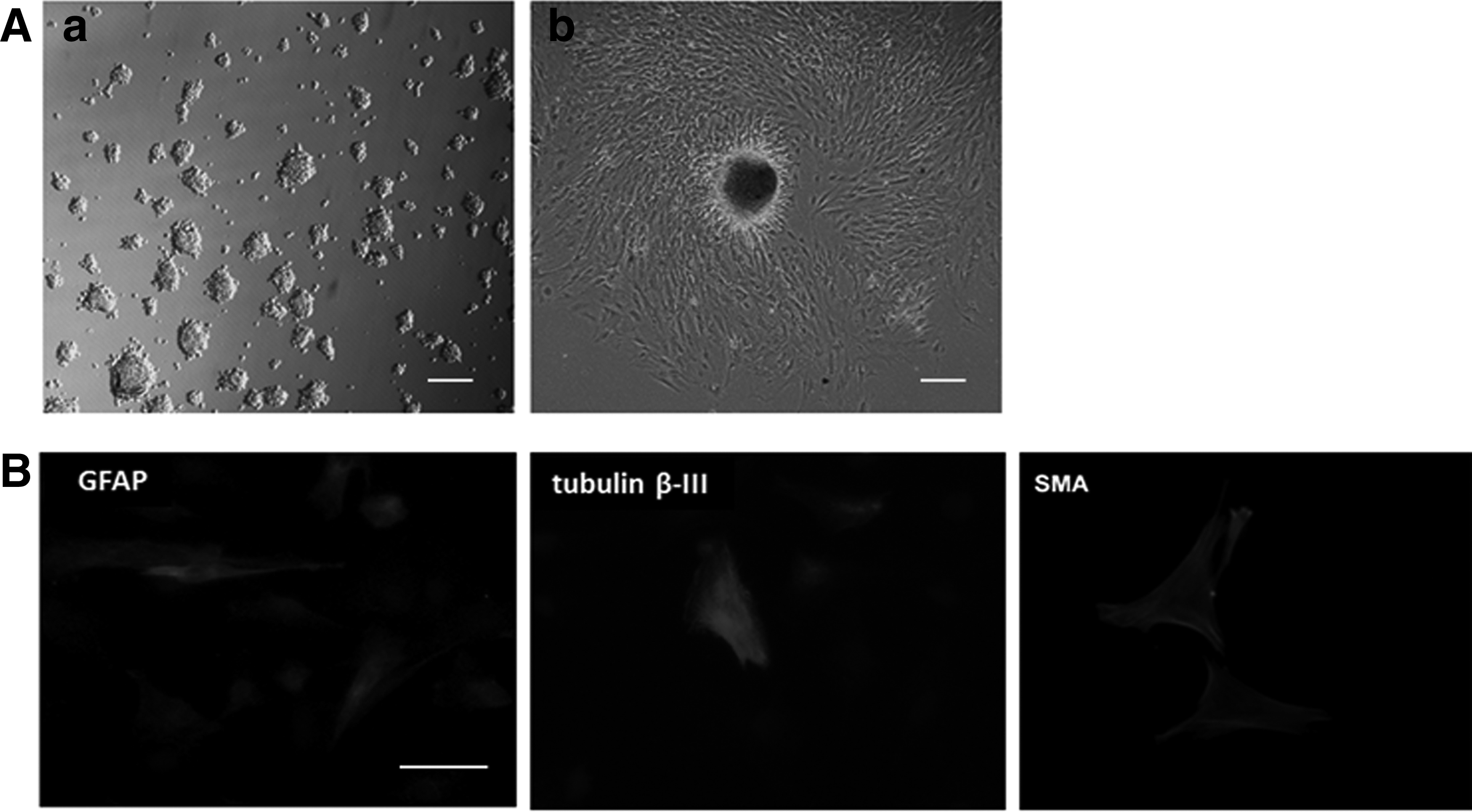
Immunocytochemical analysis revealed that the bovine SKPs generated numerous morphologically complex neurons that were positive for tubulin β-III and GFAP (Fig. 5B). For glial differentiation, the expression of GFAP was observed. For neuronal differentiation, a tubulin β-III-positive subpopulation was present after differentiation was completed. This finding indicated the neuronal potency of SKPs. In the same differentiation cultures for neurons, a small number of cell population expressed smooth muscle cell markers (SMA) (Fig. 5B). Therefore, bovine SKPs can generate neural and mesodermal cell types, including neurons, glia, and smooth muscle cells, in vitro. Our results further indicated the multiple lineage potential of SKPs.
Development of cloned embryos in vitro
The embryos from SKPs showed significantly higher cleavage rate than those from BEFs (Table 2; 79.50% ± 1.58% vs. 68.25% ± 1.71%, p < 0.01).The percentage of embryos that developed to the blastocyst stage was significantly higher in the SKP group than in the BEF group (Table 2; 41.25% ± 1.27% vs. 21.00% ± 2.94%, p < 0.01). The percentage of hatched SKP embryos was considerably higher compared with BEF embryos. Some representative photographs of preimplantation embryos (Fig. 6A–D) are provided. Moreover, the total number of cells of the SKP-cloned embryos in the blastocyst stage was significantly higher than that of the BEF-cloned embryos (Fig. 6E, F; Table 3; p < 0.05).
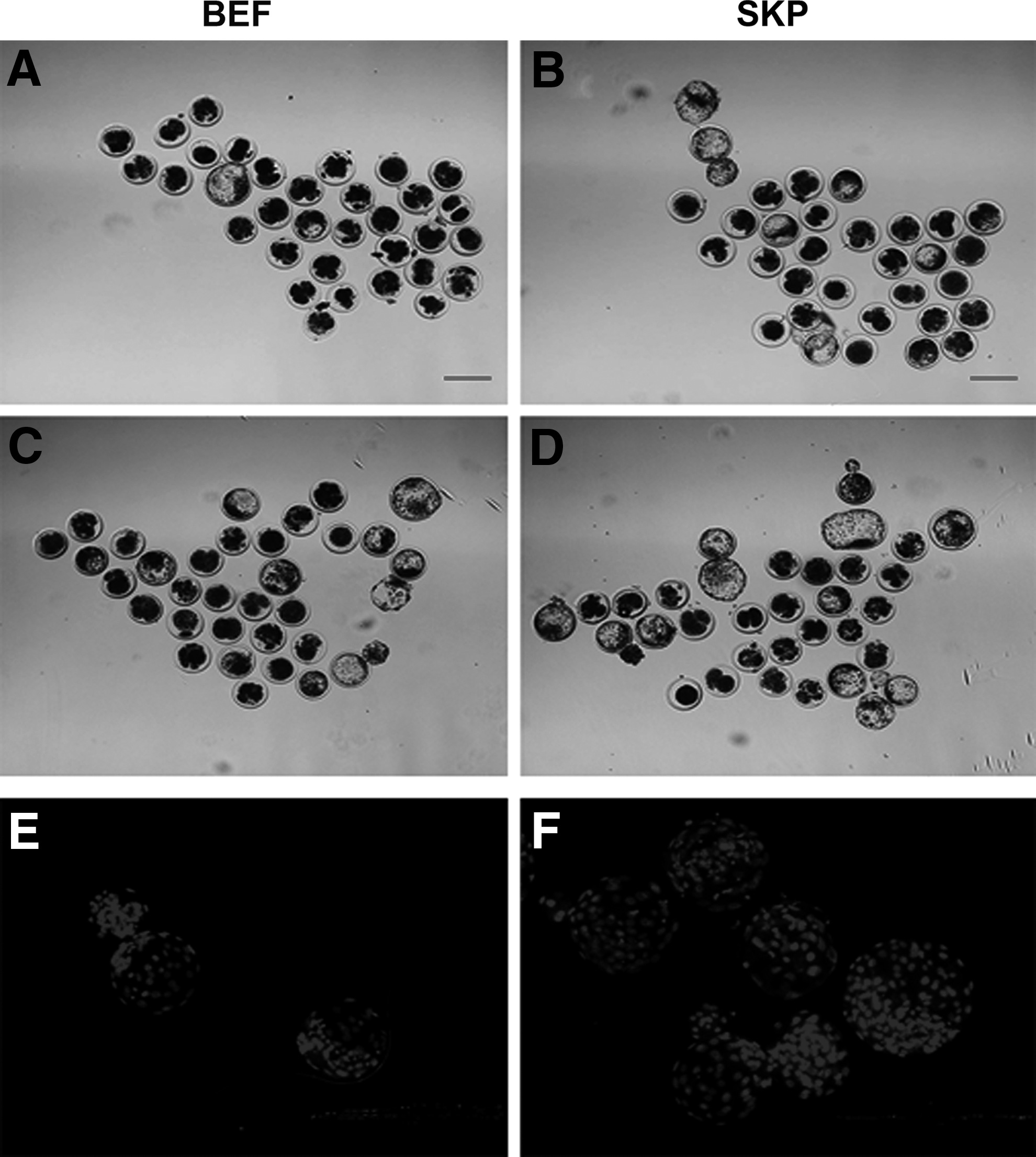
Representative photographs of preimplantation embryos.
Cleavage percentage: no. of embryos cleaved/no. of embryos cultured; Blastocyst percentage: no. of blastocysts/no. of embryos cultured; Hatched blastocyst percentage: no. of hatched blastocysts/no. of blastocysts; The experiments were repeated at least three times.
In the same column, values without a common superscript differed (p < 0.05).
BEF, bovine embryonic fibroblast; SKP, skin-derived precursor.
In the same column, values without a common superscript differed (p < 0.05).
Discussion
Thus far, this study is the first to isolate bovine SKPs (Fig. 1). Similar to human, rodent, and swine SKPs, bovine SKPs contained pluripotency-related genes simultaneously expressed with neural crest marker genes (Fig. 2) (Fernandes et al., 2004; Hunt et al., 2009; Toma et al., 2005; Wong et al., 2006; Zhao et al., 2009). The results of QRT-PCR showed that the bovine SKPs were consistently positive for neural pluripotent stem cell markers, namely OCT4, NANOG, SOX2, NGFR, SNAI2, PAX3, and SOX9 (Fig. 3A). This finding indicated that bovine SKPs are of neural crest origin and share a common transcriptional profile with neural crest stem cells.
SCNT is an efficient method for producing transgenic animals (Baguisi et al., 1999; Cibelli et al., 1998; Kato et al., 1998). SCNT has been successfully utilized for producing cloned horses, swine, mice, and rabbits (Choi and Hinrichs, 2003; Du et al., 2009; Gao et al., 2004; Onishi et al., 2000). Many types of bovine somatic cells, including fetal fibroblasts (Baguisi et al., 1999), oviduct cells (Kato et al., 1998), mural granulose cells (Wells et al., 1999), muscle cells (Shiga et al., 1999), ear fibroblasts (Zakhartchenko et al., 1999), and cumulus cells (Kato et al., 2000), have been used for nuclear transfer (NT).
However, the term development rate of blastocysts derived from ES cells (20%–40%) is higher than those from somatic cumulus cells (1%–3%) (Eggan et al., 2000). The cloning efficiency is significantly improved by using ES cells for NT (Wakayama et al., 1999). These results suggest that undifferentiated cells are more easily reprogrammed or may require less reprogramming after NT than differentiated cells (Oback and Wells, 2002; Stadtfeld and Hochedlinger, 2010). Therefore, we examined whether bovine SKPs can be used to improve somatic nuclear reprogramming and development of cloned bovine embryos in vitro.
Our results revealed that the embryos cloned from SKPs yielded significantly higher cleavage rates than those from BEFs (Table 2). Moreover, the total number of cells in SCNT blastocysts was increased by cloning with the use of SKPs (Table 3). These results showed that embryos cloned from SKPs exhibited better developmental rates than those from BEFs. H3K9 methylation has been considered as a major reprogramming barrier in somatic cells (Chen et al., 2013). Using Western blots, we found that the amount of critical epigenetic barrier, H3K9me3, in SKPs was much lower than that in BEFs (Fig. 3B). This difference may account for the higher cleavage rates of embryos cloned from SKPs.
In conclusion, we successfully isolated bovine SKPs. The embryos cloned from SKPs exhibited higher developmental potential than those from BEFs. SKPs may provide an excellent candidate for SCNT in bovines.
Footnotes
Acknowledgments
The authors are grateful to other colleagues in their laboratory for the instructive suggestions. This work was supported by the National Science and Technology Major Project in New Varieties Cultivation of Transgenic Organisms (No. 2013ZX08007-004).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.