
Abstract
Abstract
The therapeutic potential of multipotent stromal cells (MSCs) largely depends on the isolation and expansion methods used. In this study, we propose a laminin-based technique to select and enrich for MSCs isolated from the mouse testis. Primary cell cultures were prepared from juvenile mouse testes and the capacity to generate colony forming units together with population doubling time (PDT) during expansion were determined. The identity of MSCs was assayed using reverse transcription–polymerase chain reaction (RT-PCR) and flow cytometry for the active expression of cell surface markers, such as CD44, CD73, and CD29; absence of the CD45 hematopoietic cell marker; and in vitro differentiation of the cells into osteoblasts and adipocytes. Testis-derived MSCs (tMSCs) displayed self-renewal properties and in the early passages, exhibited high proliferation patterns with an average PDT of 44.1 hours. The lack of Vasa expression implied that the tMSCs were not of germ cell origin. The RT-PCR data, which were confirmed by immunophenotyping, revealed high expression of CD44 and the absence of CD45 expression in tMSCs. The strong Alizarin Red stain in tMSCs that were stimulated into making bone cells was indicative of the presence of calcium-producing cells (osteoblasts). Likewise, the adipogenic potential of tMSCs was demonstrated based on Oil Red O staining of lipid vacuoles in differentiated cells. Loss of fibroblast-like morphology in late passage cells along with the increase in PDT and the decrease in the mRNA levels of CD73 and CD29 suggested that the tMSCs developmental program is reformed at this stage.
Introduction
S
Adherence to plastic is the first and simplest method used to isolate MSCs from different sources (Lennon and Caplan, 2006). Advanced techniques have been utilized to generate more homogenous populations early on in culture. The ability of MSCs to bind to extracellular matrix (ECM) proteins has been recently utilized to enrich for MSCs. In fact, in 2012, collagen type IV was used to enrich and characterize human dermal stem/progenitor cells (Shim et al., 2012).
Spermatogonial stem cells (SSCs) are considered to be the most common source of stem cells in the testis, and these cells are only committed to generate sperm (Singh et al., 2011). However, a new source of stem cells that resembles MSCs in nature was recently isolated. There are a few studies on testis-derived MSCs from humans and monkeys (Chikhovskaya et al., 2014; Eildermann et al., 2012; Gonzalez et al., 2009). These reports postulated that testis-derived MSCs are not of germ cell origin and agreed on their multipotency differentiation potential and expression of MSC surface markers.
On the other hand, studies on testis-derived MSCs are insufficient. There is inconsistent data regarding the types of surface markers expressed, and there has been little emphasis on the expansion patterns of cultured cells in vitro. The main objective of our proposed study was to utilize the mouse model to isolate and culture testis-derived mesenchymal stromal cells (tMSCs) for the first time. In addition, we aimed to validate MSC-related properties such as in vitro proliferation patterns, surface marker expression profiles, and differentiation potential. We also assessed the growth kinetics of the isolated cells and the effect of serial passaging on the expression of MSC surface markers.
Materials and Methods
Animals
Male CD-1 Swiss albino mice (4–5 weeks old) were obtained from the Theodor Bilharz Research Institute (TBRI, Giza, Egypt). Animals were bred on a standard commercial pellet diet at the institute (El-Kahira Company for Oils and Soap, Cairo, Egypt). Animal experiments were carried out according to internationally valid guidelines.
Primary cell cultures
Preparation of testicular single cell suspensions
Three independent enrichment experiments were performed utilizing a total of 12 male mice. Animals were euthanized and dissected. The testes were subsequently harvested and washed in sterile filtered phosphate-buffered saline. For each experiment, the testes of four mice were pooled together. In sterile conditions, the testes were cut into small pieces and exposed to a two-step enzymatic digestion procedure (Guan et al., 2009) using sterile filtered 1 mg/mL collagenase type IV (Worthington Biochemical Corp.) (4–5 mL/mouse) followed by trypsin (GIBCO) (3 mL/mouse). The resulting cell suspension was filtered using a sterile 70-μm pore size Nylon mesh (BD Bioscience).
Enrichment for tMSCs and cell culture
Cells were adhered to laminin to enrich for tMSCs. Testicular cell suspensions were plated on laminin-coated dishes (Sigma-Aldrich Co.) at a density of ≈2.25 × 105/cm2. After incubation, the adherent cells were trypsinized and seeded on 6-well plates coated with 0.1% gelatin solution (Sigma) at a seeding density of 70,000 cells/cm2 in tMSC culture media. The steps for tMSCs enrichment have been summarized in Figure 1.
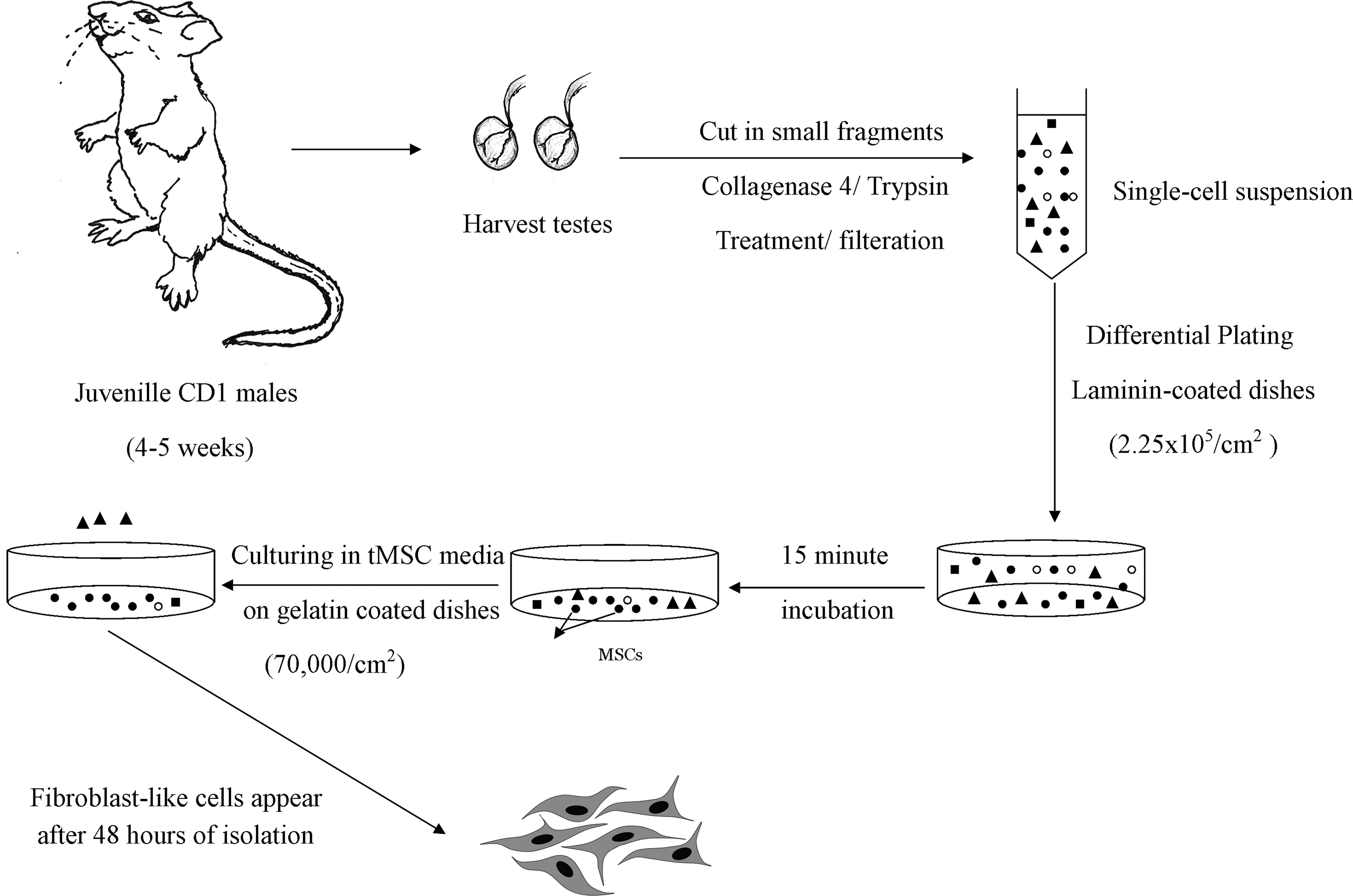
Representation of cell isolation procedures, culture conditions, and plating densities for the isolation of tMSCs. tMSCs, testis-derived multipotent stromal cells.
Primary cells were cultured on gelatin-coated dishes in complete media composed of minimum essential medium (α-MEM, GIBCO) supplemented with 1% fetal bovine serum (FBS; GIBCO), 5% penicillin/streptomycin (GIBCO), 1X nonessential amino acids (GIBCO), 1X β-mercaptoethanol (10 mM, Serva, Germany), 0.2% bovine serum albumin (GIBCO), 1X Bottenstein's N2 supplement (Life Technologies), 20 ng/mL recombinant rat glial cell line-derived neurotrophic factor (R&D Sciences), 10 ng/mL basic fibroblast growth factor (Life Technologies), and 1000 U/mL of recombinant human leukemia inhibitory factor (Life Technologies).
Colony formation assay
To test the self-renewal capacity of tMSCs, Cells at passage 3 (P3) were seeded on gelatin-coated 35 mm dishes at 500 cells/dish. Cells were incubated at 37°C for 14 days to develop colonies. Then, the plates were stained with 0.1% gentian violet in methanol (Oxford Lab Chem- Vasai Maharashtra, India). Groups of cells of 20 or more were considered as colonies. The colony forming units (CFU) were visualized using inverted microscope (Olympus 1 × 70) and cells were counted using ImageJ software (NIH). Experiments were performed in duplicates.
Population doubling time
To analyze the growth kinetics of tMSCS, the population doubling time (PDT) was calculated at every passage according to the following equation: TD = tplog2/(logNt-logNo), where Nt is the number of cells harvested, whereas No is the number of cells seeded, and t is the time in culture (in hours). The trypan blue exclusion test was used to count the number of cells with a hemocytometer (Hausser Scientific).
Characterization of tMSCs
Reverse transcription–polymerase chain reaction
Total RNA was extracted from cultured tMSCs at different passages (P0, P1, P2, P3, P4, P5, and P6) using TRIzol® (Invitrogen) as per the manufacturer's protocol. RNA was extracted from the pellets from the testicular cell suspension before enrichment for comparison. cDNA was synthesized from 1 μg of RNA using RevertAID™ First Strand cDNA synthesis kit (Fermentas).
The cDNA subsequently served as a template for PCR using gene-specific primers. Table 1 lists the sequences of the primers used to analyze the following genes: Vasa, CD44, CD73, CD29, CD45, and the loading control β-actin. A negative control was prepared for each run without the template cDNA. Finally, the amplified PCR products were electrophoresed on a 1.5% agarose gel. Densitometric analysis of the bands was performed using ImageJ software (NIH) and each band was normalized against the loading control.
Flow cytometry
Testicular cell suspensions and cultured tMSCs at P1 and P2 were stained with antibodies for immunophenotyping. Briefly, cells were incubated for 30 min at 4°C in the dark with a PE-conjugated CD44 antibody and FITC-conjugated CD45 antibody (Beckman Coulter, Mexico). At least 10,000 events were acquired on a COULTER Epics XL flow cytometer (Beckman Coulter) and analyzed using XL SYSTEM II™ software (Beckman Coulter). To adjust the threshold positivity, all experiments included unstained cells that served as negative controls.
Osteogenesis differentiation assay
To induce osteogenesis, tMSCs at P5 were cultured in osteogenesis differentiation media consisting of MEM alpha media (GIBCO) supplemented with 10% FBS (GIBCO), 5% penicillin/streptomycin (GIBCO), 1X L-Glutamine (GIBCO), 10−7 M dexamethasone (Sigma-Aldrich), 10 mM β-glycerophosphate (Sigma-Aldrich), and 50 μM ascorbic acid (Sigma-Aldrich). After 21 days, cells were fixed with 4% paraformaldehyde (Mensing et al., 2011) and stained using a 2% Alizarin Red S solution (Biostain Ready Reagents Ltd., United Kingdom). Using ImageJ software, calcification was quantified by determining the amount of stain incorporated into the cells. Noninduced cells were cultured in regular growth media and stained to serve as the negative control.
Adipogenesis differentiation assay
To induce adipogenesis, tMSCs at P3 were seeded, at 10,000 cells/cm2, on gelatin-coated dishes and grown in adipogenesis differentiation media. The differentiation media consisted of MEM alpha media (GIBCO) supplemented with 10% FBS (GIBCO), 5% penicillin/streptomycin (GIBCO), 0.5 μM dexamethasone (Sigma-Aldrich), 0.5 μM isobutylmethylxanthine (Sigma-Aldrich), 50 μM indomethacin (Sigma-Aldrich), and 5000 nM insulin (Sigma-Aldrich). The cells were cultured in adipogenesis differentiation media for 14 days. To evaluate the adipogenic differentiation, cells were fixed with 4% PFA and stained using 0.5% Oil Red O (Oxford Lab Chem- Vasai Maharashtra, India).
Statistical analysis
Data are presented as the mean ± standard deviation (SD). Comparisons were carried out using one-way analysis of variance followed by Tukey–Kramer's test for post hoc analysis. p < 0.05 was considered to be statistically significant. All statistical analyses were performed and plotted using GraphPad Prism software, version 5.00 (GraphPad Software).
Results
Enrichment for tMSCs
Enzymatic digestion of testes yielded an average cell count of 2.34 ± 0.49 ( × 106 ± SD)/paired testes. Incubating the testicular cell suspension on laminin-coated dishes yielded an average of 3.71 ± 0.81 ( × 105 ± SD) cells/paired testis adherent cells that constituted ∼16% of the initial cell suspension. Following enrichment, spindle-shaped fibroblast-like cells started to appear 24–36 hours after plating (Fig. 2A, B). After ∼4 days from the start of the enrichment experiments, tMSCs reached 80%–90% confluency and were comprised of around 50,000 cells/cm2. At this stage, tMSCs were termed “P0” and were ready to be propagated.

Characteristics of tMSCs in culture.
Culturing tMSCs
At P0, cells were morphologically found to be highly heterogeneous consisting of both round and spindle-shaped cells (Fig. 2A). The heterogeneity of the population appeared to decrease with subsequent passaging, and the cell population became nearly homogeneous at P2. Cells cultured in vitro for four to nine passages, adhered to plastic and during the early passages from P1 to P5 (Fig. 2C), maintained their bipolar spindle-shaped morphology. Interestingly, colonies composed of cells that resembled embryonic stem cells appeared in confluent cultures at P1 and P2 (Fig. 2D). However, they disappeared with later passages.
Starting at P6, a change in the morphology started to appear that was more prominent in later passages (P7–P9). Gradually, cells showed an irregular morphology, became flattened and enlarged with circumscribed nuclei, and the amount of debris increased in the medium (Fig. 2C).
Colony formation assay showed a number of stained colonies (Fig. 2E); some were large while others were compact and small with sizes ranging from 70.64 to 231.48 μm. The tMSCs of early passages (P1–P4) exhibited a relatively high proliferation pattern with an average PDT of 44.1 ± 6.5 hour (Fig. 2E). However, the PDT increased with later passages until it became significantly different at P7 compared to P4.
The investigation of the cellular origin of tMSCs
Reverse transcription–polymerase chain reaction (RT-PCR) data of the germ cell marker at P1 and P3 showed that Vasa was not expressed in tMSCs (Fig. 3A). Vasa displayed significantly low or no expression in tMSCs at P1 compared to the high levels of expression in the testis. Vasa was also absent in the subsequent passages.
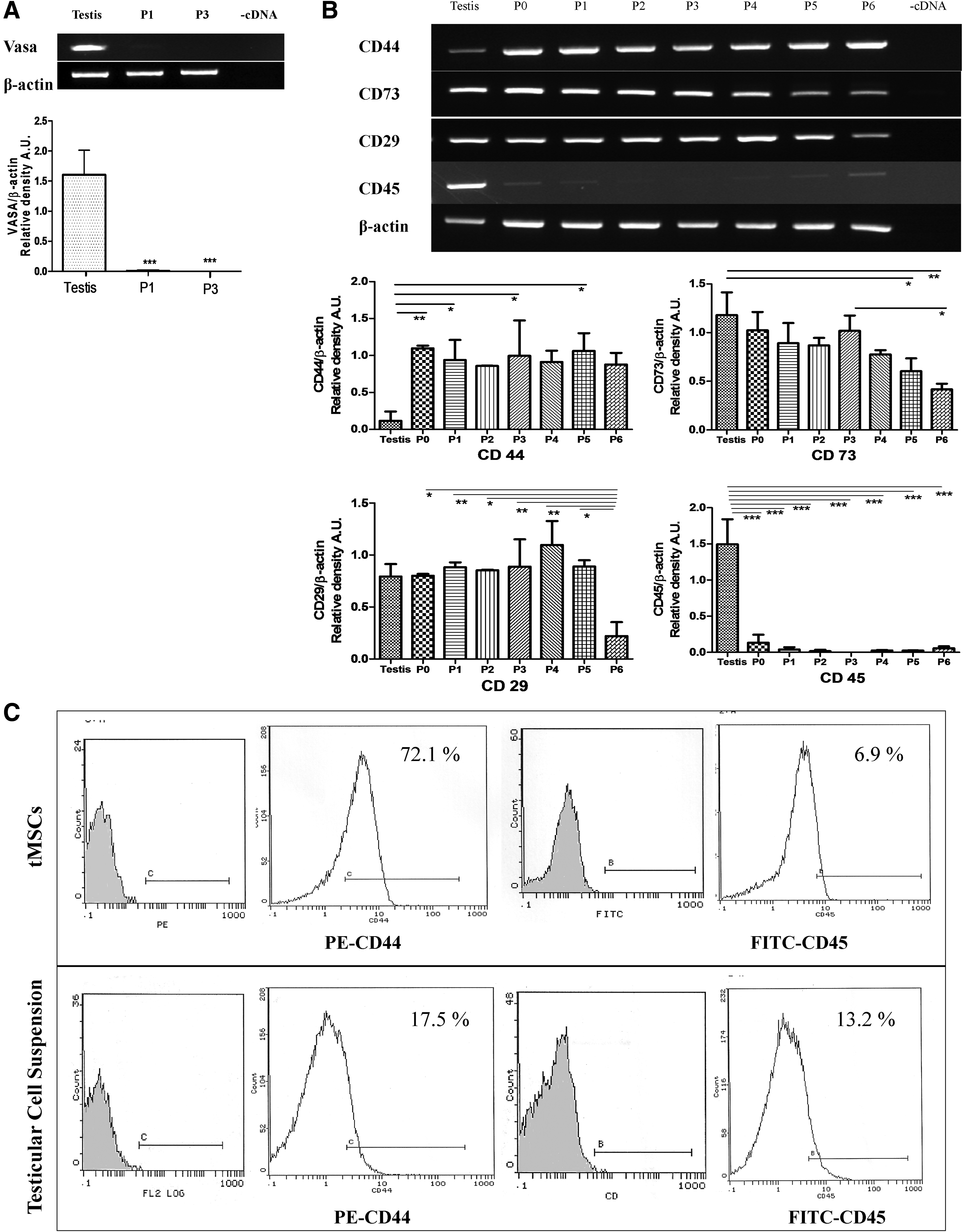
Expression of germ cell, mesenchymal stem cell, and hematopoietic stem cell markers in tMSCs.
Molecular characterization of tMSCs
RT-PCR data revealed that CD44 mRNA was highly expressed at steady state levels in the tMSCs from P0 to P6 (Fig. 3B). Interestingly, the expression levels were significantly higher than in the testis, with ∼9.6-folds greater expression at P0. The CD73 and CD29 transcript levels revealed the positive expression of both markers (Fig. 3B). Both markers displayed no significant changes in their expression in the testis. A gradual decrease in the expression of CD73 was noted until it became significantly low at P6 compared to P3.
CD29 maintained a relatively elevated expression pattern throughout different passages until it sharply decreased at P6 and became significantly different from the previous passages. The steady state RNA levels of the CD45 cell surface marker in tMSCs were negligible among all passages (Fig. 3B). The expression levels of the testicular cell suspension were ∼11.25-fold higher than in P0 tMSCS.
The surface marker expression profile of tMSCs
The immunophenotyping data showed that a high percentage of tMSCs expressed CD44 (>70%) while a low percentage of cells expressed CD45 (<7%) (Fig. 3C). On the other hand, a low percentage of cells in the testicular cell suspensions expressed both markers: CD44 < 18% and CD45 < 14% (Fig. 3C). When comparing the percentage of cells expressing both markers in the tMSCs and in the testis, CD44 was found to be higher in the tMSCs by ∼4.12-fold, while CD45 was found to be lower in the tMSCs by ∼0.5-fold.
The differentiation potential of tMSCs
tMSCs cultured in osteogenesis media exhibited mineralization in the form of calcium deposits that stained bright orange red following Alizarin Red staining (Fig. 4A, B). Mineralization was absent in noninduced control tMSCs, which stained negative for Alizarin Red (Fig. 4C, D). The quantification of mineralization using ImageJ confirmed a higher degree of mineralization in induced tMSCs compared to noninduced controls (Fig. 4E). Similarily, tMSCs cultured in adipogenesis media for 14 days exhibited intracellular lipid vesicles in the cytoplasm that were stained red following Oil Red O staining (Fig. 4C, D).
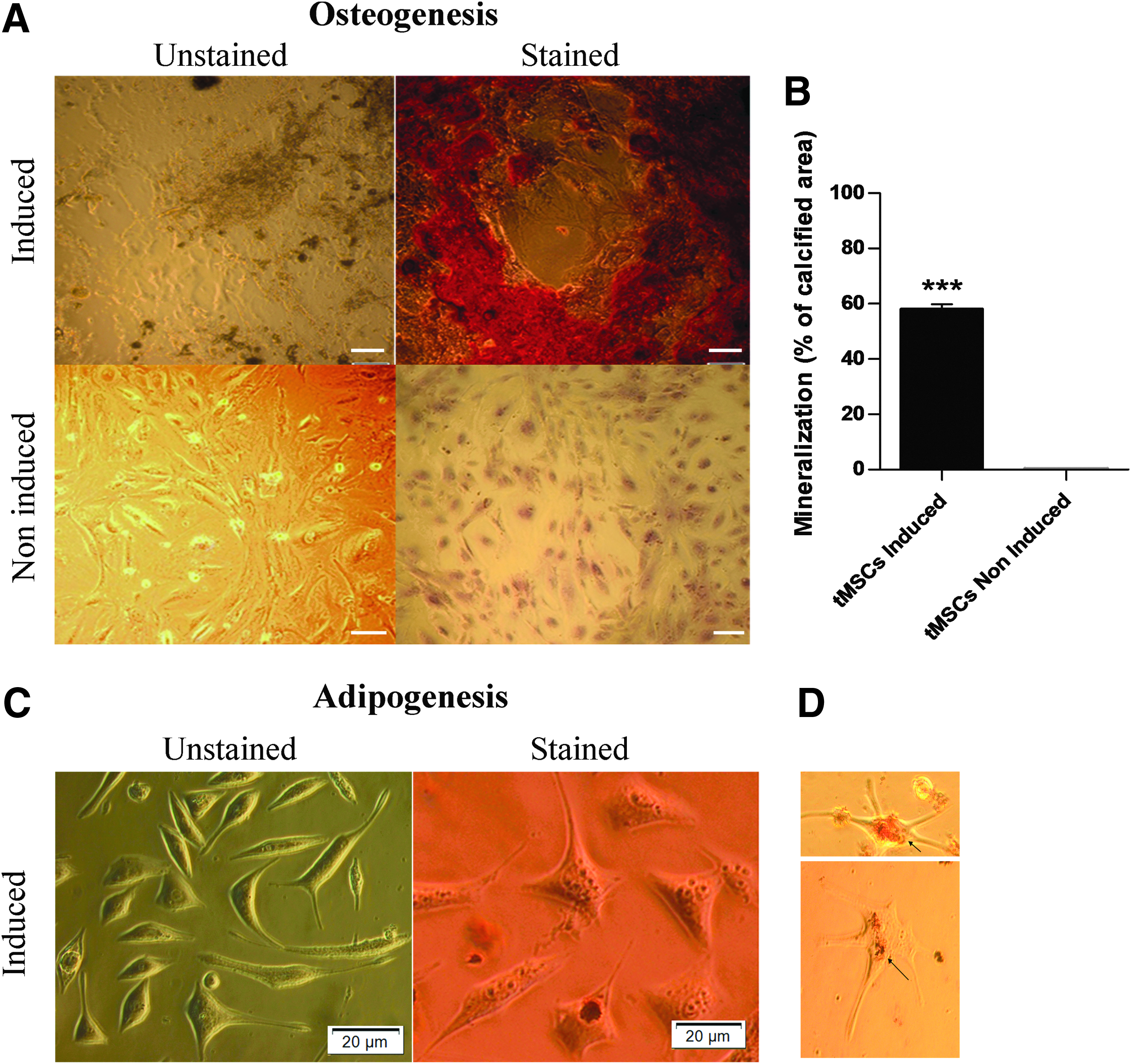
Osteogenic and adipogenic differentiation of tMSCs.
Discussion
The mouse is a simple and efficient model to isolate and culture tMSCs because of its close genetic similarity to humans and its easy manipulation. However, because the testis is composed of a highly heterogeneous cell population (Desjardins and Ewing, 1993), we introduced a simple method to enrich for MSCs from testicular cell suspensions based on positive selection by the ECM ligand laminin. This method is based on the cell binding capacity of laminin to the surface integrins on MSCs (Roncoroni et al., 2013). Furthermore, filtering the testicular suspension through a small pore size filter allowed for better selection of tMSCs.
Our enrichment technique generated successful tMSC cultures that could be propagated for up to nine passages. The homogenous population observed with early passages (P2) suggests that the enrichment technique was efficient. In addition, the cell yield (∼16%) generated from the laminin selection step is considered to be high compared to the yield generated from plastic adherence, as several studies showed that MSCs represent only 0.01%–0.001% of nucleated cells in bone marrow aspirates isolated from plastic adherence (Marcantonio et al., 2009; Muschler et al., 2001).
However, laminin-based enrichment is not considered to be a highly specific method that only selects for the desired cell population; instead, this method generates other cell types, although at lower ratios from the original testicular cell population. In fact, laminin was found to bind to SSCs (Guan et al., 2009). However, because SSCs do not bind to gelatin, they are eliminated upon subsequent culturing on gelatin-coated dishes (Wu et al., 2009). Therefore, laminin enrichment may be regarded as a simple, rapid, and cost-effective method for generating cell populations enriched with a high proportion of MSCs.
The cultured cells exhibited a morphological profile identical to that of MSCs reported in the literature (Ahrari et al., 2013; Dehghani, 2015). Upon reaching confluency, the monolayer exhibited a constant orientation and whirling tendency. In addition, the formation of colonies upon reaching confluency is a feature that was previously reported in testicular multipotent stromal cells (MSCs) isolated from the marmoset monkey (Eildermann et al., 2012). Interestingly, tMSCs showed an initial short lag or adaptation phase compared to cells isolated from the human testis, which took ∼7–10 days to reach confluency (Gonzalez et al., 2009).
CFU is a standard assay for self-renewal, where a cluster of 20 cells or more could qualify as a colony (Mensing et al., 2011). The demonstrated clonogenic capacity of tMSCs suggests that single cells gave rise to a homogenous population of cells clustered together forming colonies.
It is well established that the proliferation patterns of MSCs vary among different protocols due to variations in the culture media, source of the cells, and culture conditions. During early passages, tMSCs displayed a high proliferation rate of ∼44 hours. Although the MSCs derived from the adult human testis displayed a shorter PDT of 33.8 hours (Gonzalez et al., 2009), tMSCs displayed high proliferation patterns compared to other reports from different sources. In 2012, it was found that bone marrow-derived mesenchymal stem cells (BM-MSCs), adipose tissue-derived mesenchymal stem cells, and periosteum-derived cells at passage 2 displayed a high PDT of 3.99, 4.65, and 3.55 days, respectively (Tawonsawatruk et al., 2012).
The fast establishment of tMSC cultures and their high proliferation patterns in early passages might be attributed to several factors. Gelatin-coated matrices have been previously reported to promote the mass production of mesenchymal stem cells (Park et al., 2013). One other possible interpretation is the young age of the mice that we used, as several studies have shown that there is an inverse relationship between the age of the mice and the proliferation pattern (Kretlow et al., 2008; Shi et al., 2005). Additionally, beta-FGF was found to be associated with high proliferation capacities of testis-derived MSCs isolated from the marmoset monkey (Eildermann et al., 2012).
The change in morphology observed with later passages (starting from P6) along with the significant increase in PDT at P7 suggests that the cells may have entered a senescent or stationary phase of proliferation. The morphological changes that occurred fall in line with the changes observed in aged MSCs (Wagner et al., 2008). The time at which senescence occurs varies among different reports. However, several studies have reported results similar to ours; AT-MCSs and BM-MSCs were found to show signs of senescence and a marked increase in PDT at P6 (Christodoulou et al., 2013; Lu et al., 2006).
Our results show that tMSCs express MSC surface antigens. In fact, the tMSC surface marker profile falls in line with that of monkey testis-derived stem cells (Eildermann et al., 2012). Comparing the expression of MSC surface markers in the testicular cell suspension and the tMSCs revealed that CD73 and CD29 were not specific markers for tMSCs in the testis. In fact, several studies have reported that both markers are expressed in germ cells that are present in the testis (Martín-Satué et al., 2010; Schaller et al., 1993). On the other hand, our data revealed the specifity of CD44 to tMSCs because we found CD44 to be significantly higher in tMSCs than in the testis. This finding suggests the possible use of CD44 as a specific marker to isolate testis-derived MSCs.
Flow cytometry data revealed that a high percentage of cells expressed CD44, but the value was not above 90%. This finding could be attributed to the fact that the analysis was performed during early passages (P1 and P2). Although this stage in culture should have reached a high level of homogeneity, they have not “suffered” from long permanence in culture. However, testis-derived MSCs isolated from the monkey showed similar expression patterns, where 62%–94% of the cells expressed CD44 (Eildermann et al., 2012). As for CD45, the lack of expression in tMSCs was expected because MSCs are not known not to express hematopoietic cell markers.
Our studies on the effect of serial passaging on the expression of mesenchymal stem cell markers showed that CD44 expression is not affected by passaging. However, the expression of CD73 and CD29 is downregulated as the cells age, especially at P6. This finding appears to be consistent with morphological and growth kinetics data. Reports have shown contradicting data regarding which markers are affected by senescence. However, our data may be in line with the data of Cheng C et al. (2012) who showed that the expression of some markers, such as CD73, CD166, and Sca-1, decrease with senescence, while other markers such as CD44, CD29, and CD105, maintain constant levels of expression.
VASA is known to be a specific marker of germ cells in mice and humans (Castrillon et al., 2000; Tanaka et al., 2000). Our data on the lack of expression of Vasa in tMSCs fall in line with previously published data on testis-derived MSCs isolated from humans and marmoset monkey. In fact, these reports postulate that testis-derived MSCs originate from the interstitial stroma of the testis (Chikhovskaya et al., 2014; Eildermann et al., 2012).
Osteogenic and adipogenic differentiation capacities are considered among the key criteria to prove the multipotency potential of mesenchymal stem cells (Dominici et al., 2006). Following induction, tMSCs were proven to have both osteogenic and adipogenic potentials. This differentiation potential is consistent with MSCs isolated from the testes of humans and monkeys (Chikhovskaya et al., 2014; Eildermann et al., 2012; Gonzalez et al., 2009).
In conclusion, to the best of our knowledge, this is the first study that reports the isolation of mesenchymal stromal cells from the mouse testis. In our study, we introduced a simple and rapid laminin-based technique to enrich for tMSCs. The isolated cells were successfully cultured in vitro and displayed morphological properties, marker profiles, and differentiation capacities similar to those reported for MSCs. In addition, our study sheds some light on the proliferation patterns of testis-derived MSCs and the possible stages at which senescence may start. However, further studies need to be conducted to confirm our hypothesis on cellular aging.
The establishment of successful tMSC cultures sets the stage for further research to be conducted. Investigating the differentiation capacity of tMSCs toward germ cells could provide a new source of cells for the treatment of male infertility.
Footnotes
Acknowledgments
This study was supported by the American University in Cairo Internal Faculty Research Grant to A.A. and the Graduate Student Research Grant to Mai Abdul Rahman. We would like to thank Ms. Nancy Hassanein for her technical support.
Author Disclosure Statement
No competing financial interests exist.