
Abstract
Abstract
Use of histone deacetylase inhibitors (HDACis) is believed to improve the developmental competence and quality of cloned embryos produced. We examined the effects of treatment of buffalo fibroblasts with valproic acid (VPA), a HDACi on these cells and on embryos produced from them by hand-made cloning. VPA treatment (1.5, 3.0, or 4.5 mM) altered (p < 0.05) the growth characteristics and relative expression level of HDAC1, DNMT1, DNMT3a, P53, and CASPASE3, and the global level of H3K9/14ac, H4K5ac, and H3K18ac but not H3K27me3 in the cells. After the use of VPA-treated donor cells for producing embryos, the cleavage and blastocyst rate, and total cell number were not significantly affected; however, the apoptotic index was lower (p < 0.05) for 3.0 or 4.5 mM VPA group than for 1.5 mM VPA group or the controls. In the cloned blastocysts, the expression level of HDAC1 was higher (p < 0.05) and CASPASE3 was lower (p < 0.05), whereas that of DNMT1, DNMT3a, and P53 and the global level of H3K9/14ac were not significantly affected after VPA treatment of donor cells. In conclusion, these results suggest that VPA treatment of donor cells adversely affects their growth characteristics, increases histone acetylation, and alters gene expression but does not improve production rate of cloned embryos.
Introduction
A
Compared with a birth rate of more than 40% obtained with embryos produced through in vitro fertilization, the live offspring rate across most of mammalian species is much lower at <5% (Campbell et al., 2007). Moreover, a high incidence of abnormalities such as severe placental deficiency, prolonged gestation and dystocia, large offspring syndrome, short life span, and respiratory problems has been reported in the cloned offspring born (Yang et al., 2007; Young et al., 1998). This is believed to be primarily because of the abnormality of cloned embryos as a result of incomplete or incorrect epigenetic reprogramming of the donor somatic cell nucleus from its differentiated status into the totipotent state of the early embryo.
Epigenetic reprogramming involves establishment of appropriate epigenetic modifications such as DNA methylation and histone modifications in the cloned embryos. DNA function and gene expression are regulated by interactions between DNA methylations and acetylation or methylation of the core histones (Li et al., 2007). Epigenetic reprogramming leads to an erasure of the gene expression program of the respective donor cell and the establishment of the well-orchestrated sequence of expression of an estimated number of 10,000–12,000 genes regulating embryonic and fetal development (Niemann et al., 2008).
The major epigenetic errors, which include abnormal DNA methylation or histone modification, have a profound influence on the developmental competence of cloned embryos and the cloning efficiency (Santos et al., 2003). Therefore, improvement in the epigenetic status of the cloned embryos is considered to be one of the most important ways of enhancing the cloning efficiency. Two approaches have been used for achieving this. The first approach, which is aimed at reducing hypermethylation in the cloned embryos, involves use of drugs such as 5-aza-2′-deoxycytidine (5-aza-dC) to inhibit DNA methyltransferases, a class of enzymes responsible for DNA methylation.
The second approach involves inhibiting the activity of histone deacetylases, enzymes involved in removal of acetyl group from histones, with histone deacetylation inhibitors (HDACis) such as Trichostatin-A (TSA), scriptaid, and oxamflantin. There are several studies in cattle (Ding et al., 2008; Enright et al., 2003; Wang et al. 2011a, 2011b) and buffalo (Panda et al., 2012; Saini et al., 2016) on the use of these drugs for correcting abnormal epigenetic marks and improving reprogramming efficiency and developmental competence by treatment of donor cells or cloned embryos or both with them.
Valproic acid (VPA) is a short-chain fatty acid HDACi that improves the reprogramming of somatic cells for induction into induced pluripotent stem cells across several species. It was shown to induce reprogramming of mouse fibroblasts with only three transcription factors, with an efficiency higher than that with TSA (Huangfu et al., 2008a), and enabled reprogramming of primary human fibroblasts with only two factors, Oct4 and Sox2, without the need for the oncogenes c-Myc or Klf4 (Huangfu et al., 2008b). Treatment of reconstructed embryos with VPA has been shown to increase their developmental competence and improve their epigenetic status in many studies across several species such as mouse (Costa-Borges et al., 2010), pig (Huang et al., 2011; Kang et al., 2013; Kim et al., 2011), miniature pig (Miyoshi et al., 2010), and cattle (Song et al., 2014; Xu et al., 2012).
VPA treatment of reconstructed porcine embryos has been found to improve the live birth rate (Huang et al., 2011; Kang et al., 2013). However, in all these studies, reconstructed embryos were given VPA treatment. It has been shown in several studies that treatment of donor cells with HDACis has beneficial effects on the developmental competence, quality, or histone acetylation status of cloned embryos (Enright et al., 2003; Saini et al., 2016). There are very few studies in which the effects of treatment of donor cells with VPA on cloned embryos were examined (Sangalli et al., 2014; Selokar et al., 2013).
There is no report available on the effects of VPA treatment in buffalo. In this study, we treated buffalo fibroblast cells with VPA and examined its effect on their growth characteristics, global levels of some important histone modifications such as H3K9/14ac, H3K18ac, H4K5ac, and H3K27me3, and relative expression level of HDAC1, DNMT1, DNMT3a, P53, and CASPASE3. Then, we used the VPA-treated cells as donor cells for producing embryos by hand-made cloning (HMC) and examined the effect of VPA treatment on their developmental competence, level of apoptosis, global levels of H3K9/14ac, and the relative expression level of the already mentioned genes in cloned blastocysts.
Materials and Methods
All the chemicals were purchased from Sigma Chemical Co. (St. Louis, MO), the disposable plastic ware was from Nunc (Roskilde, Denmark), and the media were from GIBCO (Grand Island, NY) unless otherwise mentioned. In vitro culture of cells, oocytes, and embryos was carried out at 38.5°C in a CO2 incubator (5% CO2 in air).
Cell culture and VPA treatment
Buffalo ear skin fibroblasts that have been derived from an adult female (MU-5517), established in culture and cryopreserved earlier (Saini et al., 2016), were used in this study. Before each experiment, the cells were thawed and cultured in culture medium, which consisted of DMEM +10% fetal bovine serum (Gibco cat no #26140087) + 50 μg/mL gentamicin. The cells were seeded at a concentration of 1.5 × 105 cells/well in a six-well plate and were grown to 70%–80% confluence. For examining the effects of VPA, the cells were cultured for 24 hours in culture medium containing VPA (0, 1.5, 3, or 4.5 mM). To avoid difference in results caused by ageing, we have used cells between passages 5 to 8 for drug treatments.
Examination of morphology and growth characteristic of treated donor cells
For examination of morphological changes in the cells, images were taken on an inverted microscope (Nikon, Tokyo, Japan) at 200 × magnifications after 24 hours of culture in the presence of VPA. After culture of cells for 72 hours in the presence of VPA, the population doubling time was determined using the software (www.doubling-time.com). The cells were cultured for 120 hours in the presence of VPA for examining the effect of VPA on cell proliferation. The number of cells/well was counted using a TC20™ automated cell counter (Bio-Rad Laboratories, Hercules, CA) every 24 hours after trypsinization. The medium was changed with fresh medium containing the respective concentration of VPA at 72 hours.
Determination of cell viability of treated donor cells
Cell viability was determined by trypan blue exclusion test and MTT (3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide) assay after 24 hours of VPA treatment. For MTT assay, the treated cells were incubated with 5 mg/mL MTT at 38.5°C for 2 hours after which DMSO (diluted 1:1 with the culture medium) was added and the reactants were mixed until the formazan crystals were completely dissolved. The optical density of dissolved formazan was measured at 570 nm using a multiscan ascent plate reader. All the analyses were performed in at least three independent replicate cultures. Absorbance ratio of treated to nontreated control cells was calculated and presented as relative cell viability.
Immunofluorescence staining of donor cells for epigenetic markers
For examining the global level of H3K9/14ac, H4K5ac, H3K18ac, and H3K27me3 in VPA-treated donor cells by immunofluorescence staining, the cells were fixed in 4% paraformaldehyde for 1 hour at 38.5°C, washed three times with Dulbecco's phosphate-buffered saline (DPBS), and then permeabilized in 0.5% Triton X-100 for 20 minutes at 38.5°C. The cells were blocked for 1 hour in 3% bovine serum albumin (BSA) and then incubated overnight at 4°C with the respective rabbit primary antibody. Primary antibodies, which included anti-H3K9/14ac (1:1000; Santa Cruz Biotechnology, CA,, anti-H4K5ac (1:1500; Millipore, MA), anti-H3K18ac (1:1500; Millipore), and anti-H3K27me3 (1:1500, Millipore), were diluted in 3% BSA.
After washing five times with DPBS containing 0.1% Triton X-100 (DPBST), the cells were incubated with FITC-conjugated goat antirabbit secondary antibody diluted 1:700 in DPBS. After five washings with DPBST, the nuclei were counterstained with H33342 (10 μg/mL) and rinsed in DPBST. The cells were then mounted on slides in mounting medium (2.5% DABCO in glycerol). The slides were observed under a fluorescence microscope, and the images were captured keeping the same optical conditions. NIS-element basic research image processing software (Nikon, Tokyo, Japan) equipped with the microscope was used for image acquisition and quantitative measurements of the mean pixel intensity emitted by each individual nucleus. At least 10 images (200 nuclei from each image) were analyzed for each epigenetic marker.
Production of cloned embryos and assessment of quality
Somatic cells were treated with selected doses of VPA before SCNT. Preparation of recipient oocytes (maturation, cumulus/zona removal, and manual enucleation), and fusion, activation, and culture of embryos were performed according to the methods described previously (Selokar et al., 2012). For assessment of embryo quality, total cell number (TCN) and level of apoptosis in day 8 blastocysts were determined by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining as previously described (Selokar et al., 2014).
Gene expression analysis in donor cells and embryos
RNA was isolated from 1 × 104 cells or blastocysts (n = 3–4) using the RNAqueous micro kit (Ambion, Austin, TX) according to the manufacturer's instructions. The genomic DNA contamination was removed by DNase treatment at 37°C for 20 minutes. The RT reaction was achieved using the M-MLV RT provided in superscript reverse transcriptase III kit (Invitrogen). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using the optimized primer sets shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/cell) on CFX96 real time system (Bio-Rad) with maxima@SYBR Green master mix (Fermentas, St. Leon-Rot, Germany) at the following thermal cycling conditions: 95°C for 5 minutes, followed by 40 PCR cycles of 95°C for 15 seconds, 58°C for 30 seconds, and 72°C for 30 seconds.
Melting peaks were determined using melting curve analysis to ensure the specific amplification. Agarose (2%) gel electrophoresis analysis was carried out to determine the length of the amplified PCR products. The relative quantification of gene expression was conducted using the method described previously (Livak and Schmittgen, 2000). β-actin mRNA was employed as an internal standard for the analysis of relative transcript levels of each gene and in negative controls, H2O replaced cDNA in the reaction tubes. For comparison, the average expression level of each gene from nontreated control cells or embryos was set as 1. Three separate experiments were performed with three replicates for each gene.
Immunofluorescence staining of embryos for epigenetic markers
For examining the effect of treatment of donor cells with VPA on the global level of H3K9/14ac in the cloned embryos, immunofluorescence staining was carried out as already described for donor cells except that washing and incubation solutions contained 0.3% PVA to prevent sticking of blastocysts to the surface. A minimum of 15 blastocysts (50 nuclei/blastocyst) were analyzed for H3K9/14ac epigenetic marker.
Experimental design and statistical analysis
First of all, we subjected the donor cells to treatment with graded doses of VPA, after which they were examined for morphological changes, rate of proliferation, histone modifications, and gene expression. Then, we used these cells for production of cloned embryos and examined the effects of VPA treatment on their developmental competence and quality as indicated by the blastocyst rate and the apoptotic index, respectively. We also examined the effect of VPA treatment on the epigenetic status of cloned embryos as indicated by the global level of H3K9/14ac.
Results
VPA treatment altered the growth characteristics and decreased viability and proliferation of donor cells
We found that treatment of buffalo fibroblasts with VPA altered the morphological appearance of cells, making them vacuolated, enlarged, flattened, and elongated (Fig. 1). It resulted in a significant increase (p < 0.05) in population doubling time and a decrease in the proliferation rate in a dose dependent manner. Cell viability, examined by trypan blue dye exclusion test, was lower (p < 0.05) with 3 and 4.5 mM VPA than with 1.5 mM VPA or that in controls. Relative cell viability, examined by MTT assay, was significantly lower (p < 0.05) after treatment with 4.5 mM VPA than in the other three groups.
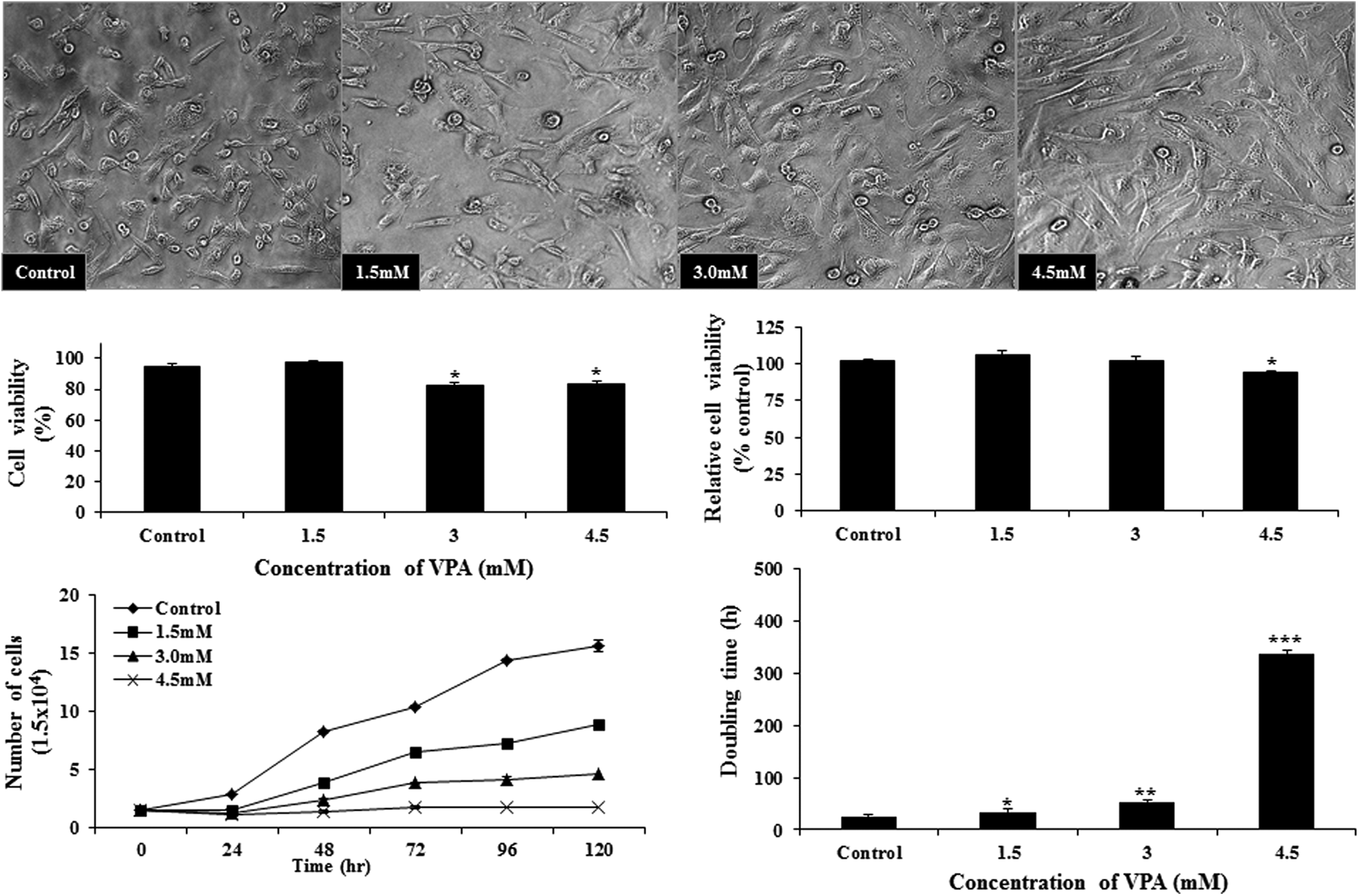
Effect of VPA treatment on the morphological appearance and growth characteristic of buffalo fetal fibroblast cells. Cell viability was measured by Trypan blue dye exclusion method, whereas relative cell viability was measured by MTT assay. Bars with different number of asterisks are significantly different (p < 0.05). VPA, valproic acid.
VPA treatment of donor cells improved the quality but not the developmental competence of cloned embryos
After use of VPA-treated donor cells, the cleavage and blastocyst rate and the TCN were not affected significantly at all the concentrations of VPA examined (Table 1). However, the level of apoptosis was significantly lower (p < 0.05) in blastocysts produced from donor cells treated with 3.0 or 4.5 mM VPA than in those produced from cells treated with 1.5 mM VPA or the controls.
Data from five trials. Values with different superscripts within the same column differ significantly (p < 0.05).
VPA treatment of donor cells increased their histone acetylation but not that of cloned embryos
As determined by the mean pixel intensity after immunofluorescence staining, VPA treatment of donor cells significantly increased (p < 0.05) the global level of H3K9/14ac, H3K18ac, and H4K5ac at all the concentrations of VPA examined compared with that in the controls (Fig. 2). However, the global level of H3K27me3 in the donor cells was not significantly affected by VPA treatment. In the cloned blastocysts, VPA treatment of donor cells did not affect the global level of H3K9/14ac at all the concentrations examined (Fig. 3).
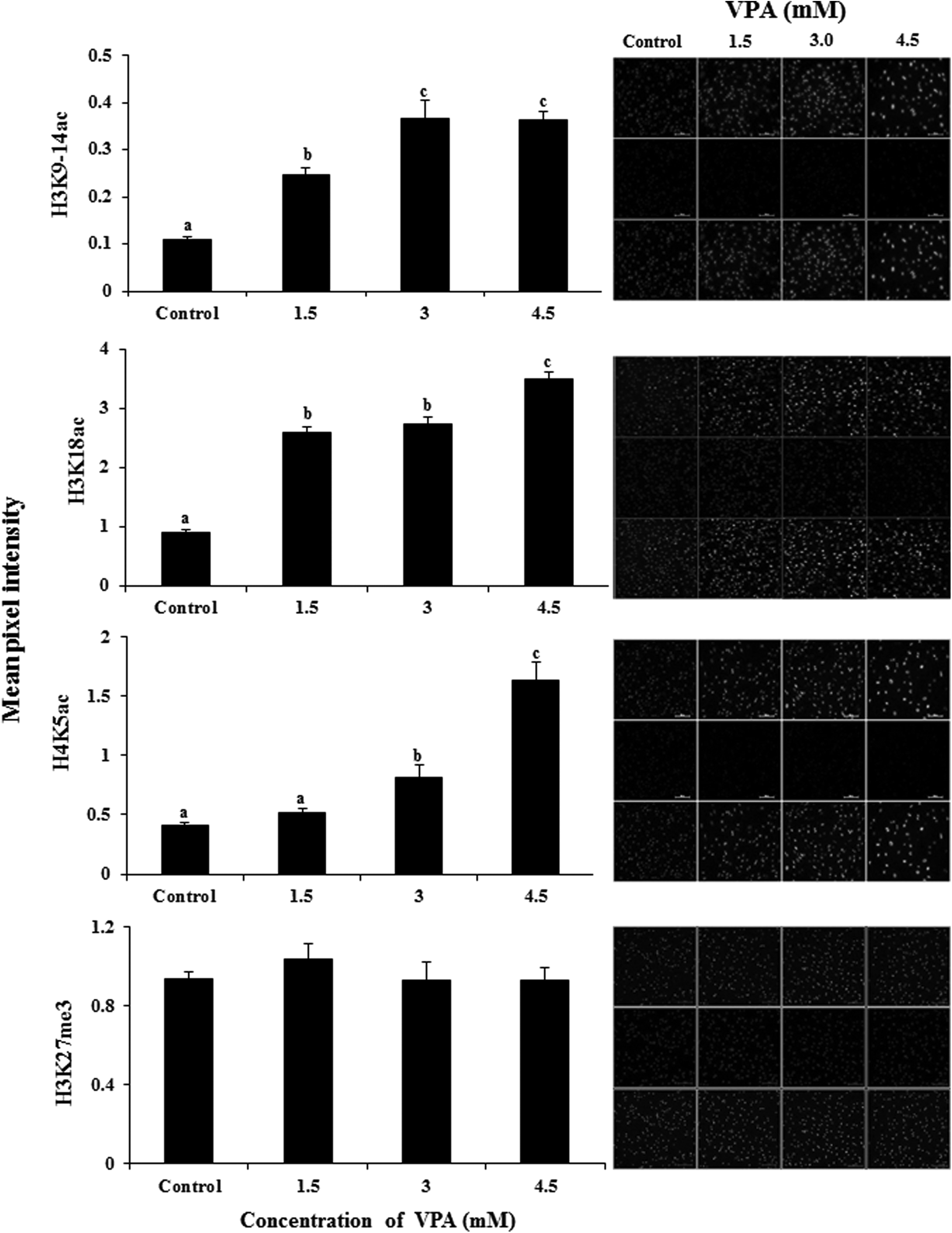
Effect of VPA treatment on the global level of epigenetic markers H3K9/14ac, H3K18ac, H4K5ac, and H3K27me3 in donor cells. Bars with different superscripts differ significantly (p < 0.05).
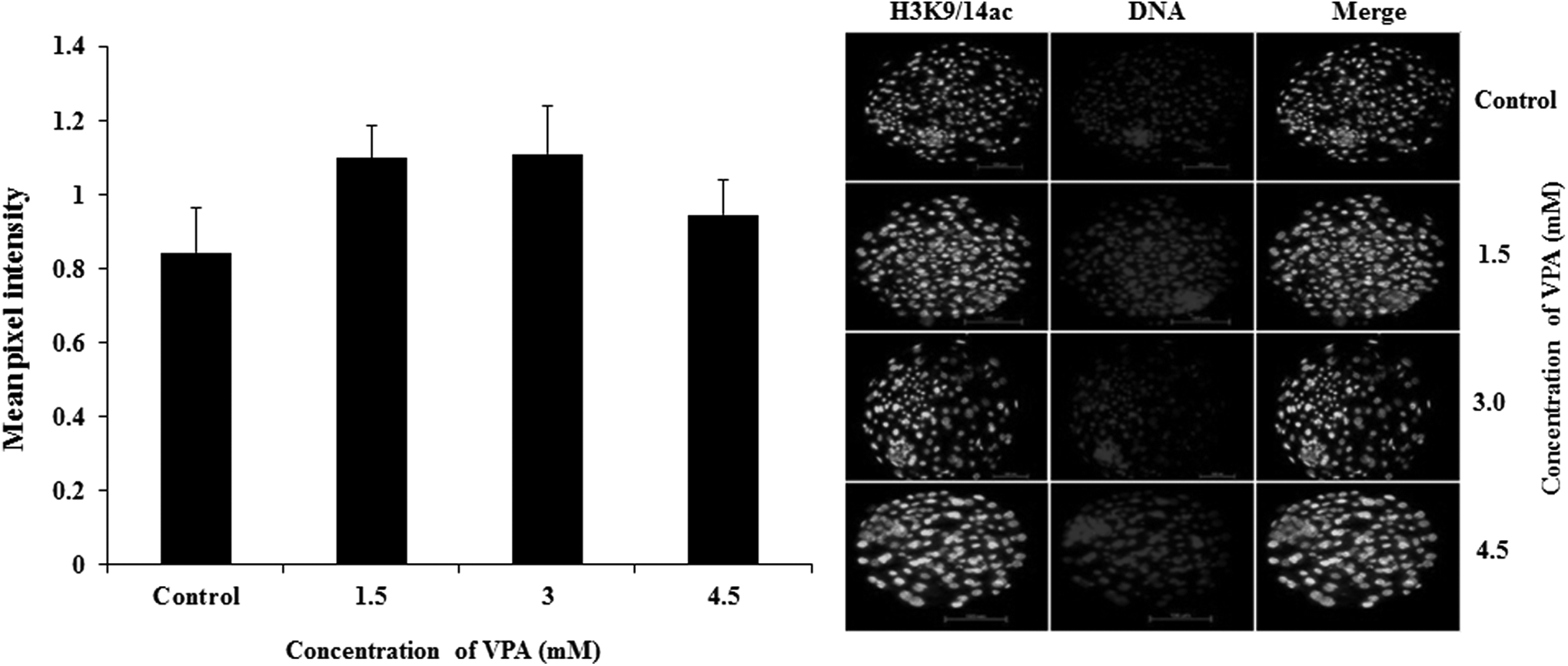
Effect of VPA treatment of the donor cells on the global level of H3K9/14ac in cloned blastocysts.
VPA treatment of donor cells altered their gene expression levels and that of cloned embryos
VPA treatment of donor cells decreased (p < 0.05) the relative mRNA abundance of HDAC1, DNMT1, DNMT3a, and P53 and increased (p < 0.05) that of CASPASE3 compared with that in the controls (Fig. 4). In the cloned blastocysts, VPA treatment of the donor cells increased (p < 0.05) the relative expression level of HDAC1 and decreased (p < 0.05) that of CASPASE3, whereas the expression level of DNMT1, DNMT3a, and P53 was not significantly affected.
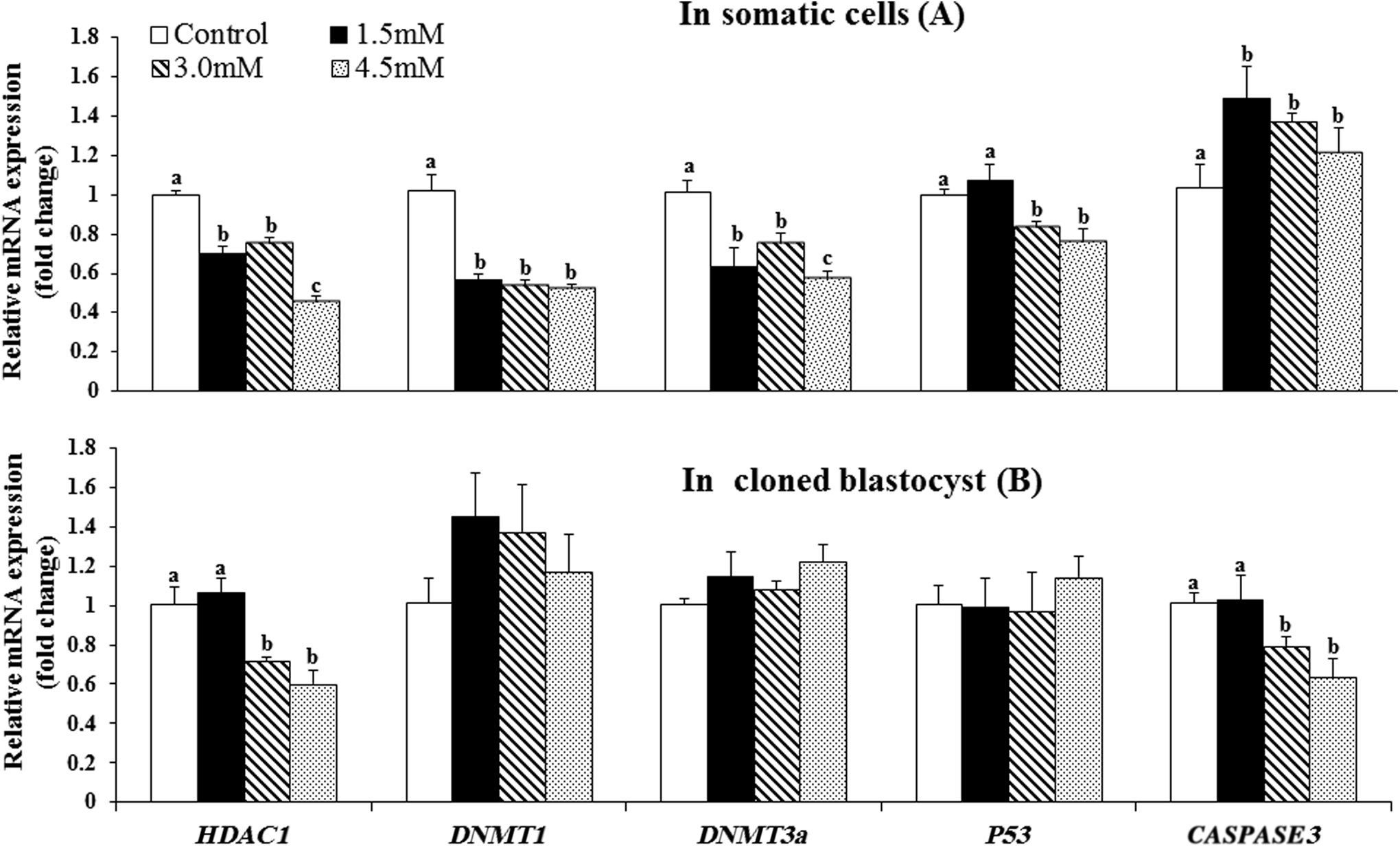
Effect of VPA treatment of the donor cells on the relative mRNA abundance of HDAC1, DNMT1, DNMT3a, P53, and CASPASE3 in the donor cells
Discussion
It has been demonstrated in many studies across several species including mouse (Costa-Borges et al., 2010), miniature pig (Miyoshi et al., 2010), pig (Kim et al., 2011), and bovine (Xu et al., 2012) that treatment of cloned embryos with VPA improved their in vitro developmental competence and increased their histone acetylation status. There are very few studies that have investigated the effects of treatment of donor cells with VPA (Sangalli et al., 2014; Selokar et al., 2013). In this study, we examined the effects of treatment of buffalo fibroblasts with VPA on the developmental competence of cloned embryos produced using these cells. We also examined the effect of VPA treatment on the epigenetic status and gene expression levels in treated cells and further investigated whether these effects are reflected in the cloned embryos produced from them.
We have earlier demonstrated that treatment of buffalo fibroblast donor cells with chromatic modifying agents such as TSA and 5-aza-dC affects the viability and growth characteristics of these cells (Saini et al., 2016). Therefore, before their use as donor cells for HMC, we examined the effects of increasing concentrations (1.5, 3, and 4.5 mM) of VPA on the morphology, viability, and growth characteristics of buffalo fibroblasts. We found that VPA treatment not only altered the morphological appearance of these cells making them vacuolated, enlarged, flattened, and elongated but also decreased the cell proliferation rate in a dose dependent manner.
Cell viability and relative cell viability, examined by trypan blue exclusion test and MTT assay, respectively, were also adversely affected by VPA treatment. These results are in agreement with those of an earlier study in which treatment of bovine fibroblasts with 2.5–10 mM VPA was found to alter their morphological appearance, increase the population doubling time in a dose-dependent manner, decrease the Ki-67 proliferation index, and induce apoptosis at doses ≥5 mM (Selokar et al., 2013).
In contrast, Sangalli et al. (2014) observed an increase in the proliferation and relative cell viability of bovine fibroblasts treated with 1, 2, or 5 mM VPA, which they attributed to a side effect of VPA on cell metabolism. In view of the adverse effects of VPA on the donor cells observed in this study and by others (Selokar et al., 2013), the highest concentration of VPA used in this study was kept at 4.5 mM.
In the next experiment, we used VPA-treated donor cells for producing cloned embryos by HMC. We found that treatment of donor cells with VPA did not affect the in vitro developmental competence of the cloned embryos produced from them as indicated by the cleavage and blastocyst rate. This agrees with an earlier report, in which treatment of bovine fibroblasts with 2 mM VPA was not found to have any significant effect on the preimplantation and postimplantation development of cloned embryos (Sangalli et al., 2014).
However, in another study, treatment of bovine fibroblasts with 2.5 mM VPA was found to significantly increase (19.9% vs. 41.2%) the blastocyst rate (Selokar et al., 2013). Lack of effect of VPA treatment of donor cells on the in vitro development of cloned embryos is in contrast with the beneficial effects of VPA treatment observed in several earlier studies, in which the reconstructed embryos, instead of the donor cells, were given VPA treatment. Costa-Borges et al. (2010) reported that treatment of cloned mouse embryos with 2 mM VPA for 8–9 hours immediately after reconstruction improved the blastocyst rate (38.3% vs. 22.9%).
Treatment of bovine embryos for 24 hours after activation with VPA increased the blastocyst rate significantly at 4 mM (39.9% vs. 34.8%; Xu et al., 2012) or 3 mM concentrations (37.7% vs. 53.3%; Song et al., 2014). Miyoshi et al. (2010) reported that VPA (4 mM) treatment of reconstructed miniature pig embryos for 48 hours after activation increased the blastocyst rate significantly (20.8% vs. 7.7%). Treatment of reconstructed porcine embryos with VPA for 24 hours increased the blastocyst rate significantly at 2 mM (21.5% vs. 10.5%; Kang et al., 2013) and 5 mM concentrations (40.8% vs. 23.4%; Kim et al., 2011). In another study, reconstructed porcine embryos treated with 1 mM VPA for 14–16 hours after activation were found to exhibit significantly higher rate of blastocyst formation (31.8% vs. 11.4%; Huang et al., 2011). Our results together with those of these studies indicate that although VPA treatment of reconstructed embryos is beneficial, VPA treatment of donor cells does not improve the in vitro development of cloned embryos.
Then, we examined the quality of embryos produced from VPA-treated donor cells by determining their TCN and level of apoptosis by TUNEL assay. We found that VPA treatment of donor cells improved the quality of cloned blastocysts produced as indicated by lower level of apoptosis although the TCN was not significantly affected. This is consistent with the lower transcript level of proapoptotic gene CASPASE3 in embryos produced from VPA-treated donor cells observed in this study.
Interestingly, VPA treatment increased the CASPASE3 transcript level in the donor cells. Similar results have been reported in bovine and miniature pig, wherein VPA treatment of donor cells (Selokar et al., 2013) or reconstructed embryos (Miyoshi et al., 2010; Song et al., 2014; Xu et al., 2012) reduced the number of apoptotic nuclei in the blastocysts without a significant effect on the blastocyst cell number. However, in other studies, VPA treatment of reconstructed embryos was observed to increase the number of inner cell mass (ICM) cells (Kim et al., 2011) or both ICM and trophectoderm cells (Song et al., 2014) in blastocysts. Together, these results demonstrate that VPA treatment improves the quality of cloned embryos irrespective of whether the donor cells or the reconstructed embryos are treated with VPA.
Among various histone modifications such as acetylation, methylation, phosphorylation, and ubiquitination, acetylation of histones, especially histone H3, is considered to be a very important modification. Acetylation level of H3 in early embryonic stages affects subsequent development (Yamanaka et al., 2009) and the acetylated form of H3 lysine 9 (H3K9ac) is associated with active chromatin configurations (Rice and Allis, 2001). DNA methylation and acetylation of H3K18 are correlated with the developmental potential of SCNT embryos and reflect nuclear reprogramming of nuclear donor cells (Santos et al., 2003).
Reprogramming involves erasure and re-establishment of the epigenetic marks of the donor cells. To find out whether VPA treatment affected the epigenetic marks of the donor cells, we examined the global levels of some important epigenetic marks. We found that the global level of H3K9/14ac, H3K18ac, and H4K5ac, but not that of H3K27me3, was increased after VPA treatment of donor cells. However, this increased acetylation in donor cells was not translated into an increase in histone acetylation in cloned embryos, at least in terms of the global level of H3K9/14ac, which was found to be similar between cloned embryos produced from VPA-treated donor cells and those produced from untreated controls.
In agreement with our results, the VPA treatment-induced increase in global level of H3K9ac in bovine donor cells was not found to be maintained after nuclear transfer in an earlier study (Sangalli et al., 2014). Our results indicate that although VPA treatment of donor cells improves the level of acetylation and alters gene expression in the treated donor cells, most of these beneficial effects are not passed on to the cloned embryos produced using these cells and are lost during reprogramming.
The mechanism behind the beneficial effects of VPA on reprogramming remains to be elucidated. The beneficial effects could be because of upregulation of embryonic stem cell-specific genes and downregulation of embryonic fibroblast-specific genes (Huangfu et al., 2008a) or maintenance of the ability of cloned embryos to express Oct-3/4 (Miyoshi et al., 2010). VPA treatment of four cell embryos has been reported to improve Oct4 expression in cloned mouse blastocysts (Isaji et al., 2013). However, the expression level of Oct4, Sox2, and Klf4 was not found to be significantly changed between VPA-treated and VPA-untreated groups at the blastocyst stage in pig (Huang et al., 2011). VPA might enhance the developmental competence of SCNT embryos by alleviating the endoplasmic reticulum (ER) stress and its associated developmental damage because VPA markedly reduced the transcript levels of ER stress markers, sXBP-1 and CHOP, and significantly increased those of GRP78/BiP, an ER stress-alleviating gene (Song et al., 2014).
In conclusion, the results of this study suggest that treatment of donor cells with VPA adversely affects their growth characteristics, increases histone acetylation, and alters gene expression but does not improve production rate of cloned embryos. This study is an exploratory study to see effect of well-known HDACi, VPA, on buffalo-cloned embryonic development, and results of this study will pertain to only one specific cell line. Therefore, further experimentation on more number of cell lines are needed.
Footnotes
Acknowledgments
This work was funded by National Agriculture Innovative Project (NAIP) grant to S.K.S. (C 2-1-(5)/2007) and M.S.C. (C-2067 and 075). Naresh Selokar and Monika Saini were recipients of CSIR-SRF fellowship.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.