
Abstract
Abstract
Induced pluripotent stem (iPS) cells are generated by reprogramming of differentiated somatic cells. These cells are identical to human embryonic stem cells (hESCs) in gene expression pattern and the ability to differentiate. iPS cells can be used in in vitro modeling of diseases, testing drugs, assessing gene therapy methods, and cell therapy. Yet, the most important and promising application of iPS cells is in regenerative medicine. Regenerative medicine is a novel area in medicine aiming at the treatment of impaired or lost tissues by replacing them with functional and healthy ones. Currently, organ transplantation, which is considered the only treatment and cure for a number of diseases, is limited by shortage of organ donors and availability of the right match. Therefore, utilization of an alternative source of cells and tissues is critical in transplantation therapy. In this study, we review recent advances in therapeutic application of iPS cells in diseases where organ transplantation remains the only solution and will discuss the potential and usage of iPS cells in different areas of regenerative medicine. The primary theory of using iPS cells in regenerative medicine has brought lots of promises due to its potential for solving the immunological, social, and ethical problems of using ESCs. Nevertheless, several issues and problems have to be resolved before applying iPS cells in therapeutic applications.
Introduction
S
ASCs, adult stem cells; CSC, cancer stem cell; ESCs, embryonic stem cells; ICM, inner cell mass; iPS, induced pluripotent stem; TS, trophoblast stem.
Induced pluripotent stem (iPS) cells are generated by reprogramming of a differentiated somatic cell. These cells are identical to human embryonic stem cells (hESCs) in gene expression pattern, surface proteins, and the ability of differentiation as pluripotent cells (Patel and Yang, 2010).
Using iPS cells has many advantages in comparison with ESCs. For example, they can be obtained from any human somatic tissues, even those with inaccessible or undetected stem cells, or patients with incurable diseases. iPS cell technology also provides an in vitro model for studying diseases (Kawamura et al., 2012), testing drugs (Xu and Zhong, 2013), and assessing gene therapy methods (Devine et al., 2011). It can also be considered a future cell source for cell therapy (Alvarez et al., 2012). Besides, there are some advantages in using iPS cells for human organ regeneration, for example, the unlimited source of cell donors, possibility of using any available cells for producing iPS cells, and elimination of invasive cell-collecting methods (Takahashi and Yamanaka, 2013).
In 2006, the first mouse iPS cell was generated using Oct4, Sox2, Klf4, and c-Myc factors (direct reprogramming) (Takahashi and Yamanaka, 2006). One year later, the first human iPS (hiPS) also was generated (Takahashi et al., 2007). Since the first discovery of iPS cells, many studies have been performed to increase the efficiency and safety of the reprogramming process for clinical applications, for example, developing new methods for delivery of the reprogramming factors instead of retroviral vectors: polycistronic vectors (Kaji et al., 2009), transient transfection (Okita et al., 2008), using small molecules (Lin et al., 2014), transposon vectors (Kaji et al., 2009), adenoviral vectors (Stadtfeld et al., 2008), episomal vectors (Yu et al., 2009), minicircle vectors (Jia et al., 2010), and recombinant proteins (Zhou et al., 2009).
Plasmid transfection (Kim et al., 2016), Sleeping Beauty (Talluri et al., 2014) and PiggyBac transposon system (Woltjen et al., 2009), reprogramming techniques based on microRNAs (Deng et al., 2015), and CRISPR/Cas9 system (Grobarczyk et al., 2015) are some of other nonviral methods that have been used for iPS cell generation.
In addition, new studies try to design a feeder-free (Caxaria et al., 2016), scaffold-free (Kamao et al., 2014), and safe culture condition for iPS generation. Figure 1 shows some of the common methods for iPS cell generation, their advantages, and disadvantages. However, despite increasing new hopes for using iPS cells in organ regeneration, the use of iPS cell technology in regenerative medicine is still limited.
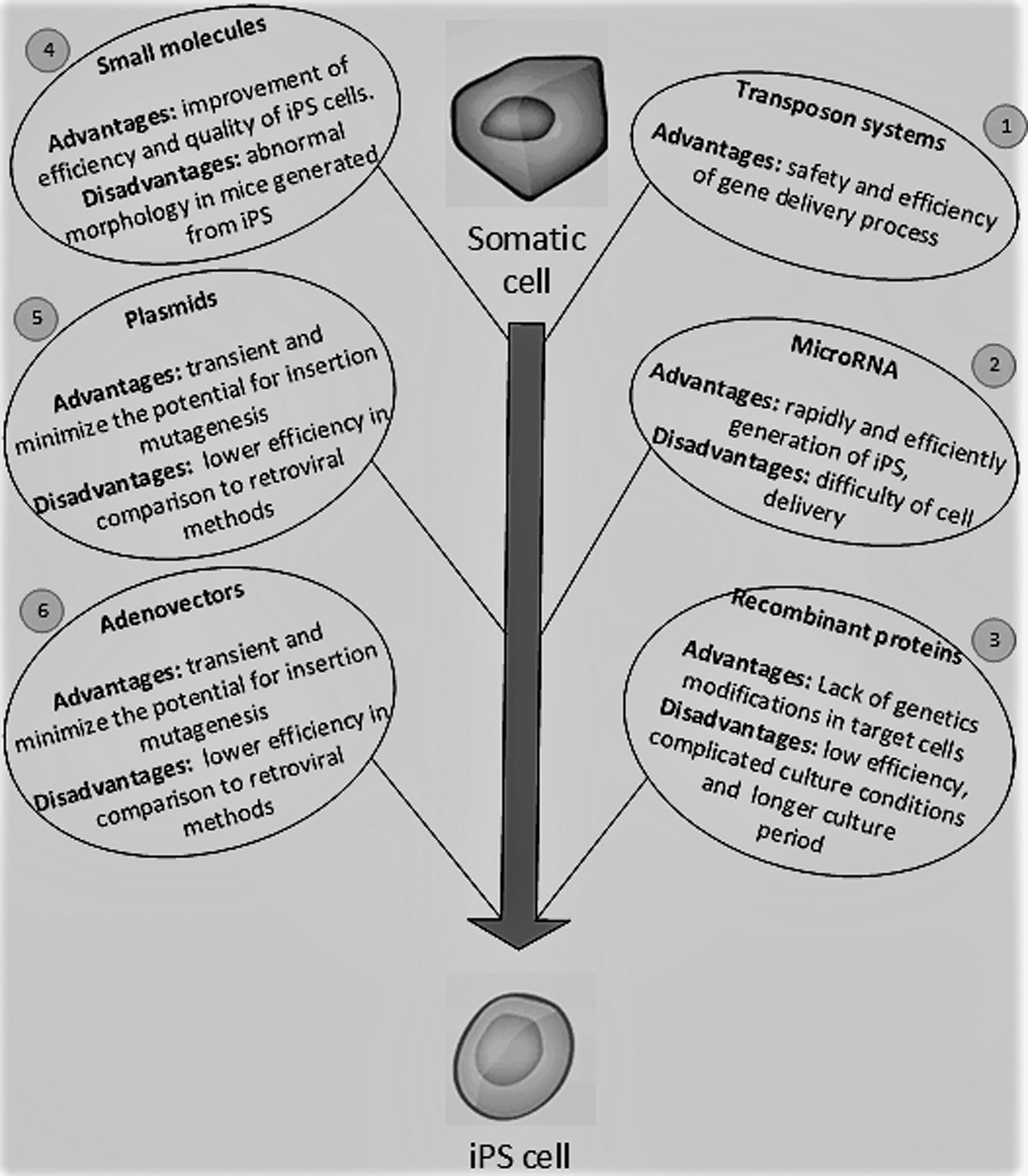
Common methods for iPS cell generation. (1) Using transposon vectors, for example, Sleeping beauty and Piggy Back (Woltjen et al., 2009), (2) using MicroRNAs, for example, microRNA 302–367 (Deng et al., 2015), (3) using recombinant proteins (Zhou et al., 2009), (4) using small molecules, for example, PD 98059 (inhibitor of ERK 1) and SC1 (pluripotin) (inhibits the RasGAP and ERK pathways) (Lin et al., 2014), (5) using plasmid-based vectors (Okita et al., 2008), and (6) using adenovirus-based vectors (Stadtfeld et al., 2008). ERK, extracellular signal-regulated kinase; iPS, induced pluripotent stem; PD, Parkinson's disease; ST, stem cell.
In this article, we reviewed new advances in the use of iPS cells, which could eventually lead to treatment of some organ-destructive diseases by regenerative medicine.
iPS Cell Applications in Regenerative Medicine
Regenerative medicine is a novel area in healthcare and its main goal is the treatment of impaired or lost tissues by replacing them with functional and healthy ones, which are created from stem cells or engineered biomaterials (Heine et al., 2011; Svendsen, 2013). Figure 2 shows some factors needed for differentiation of iPS cells into different cell types.
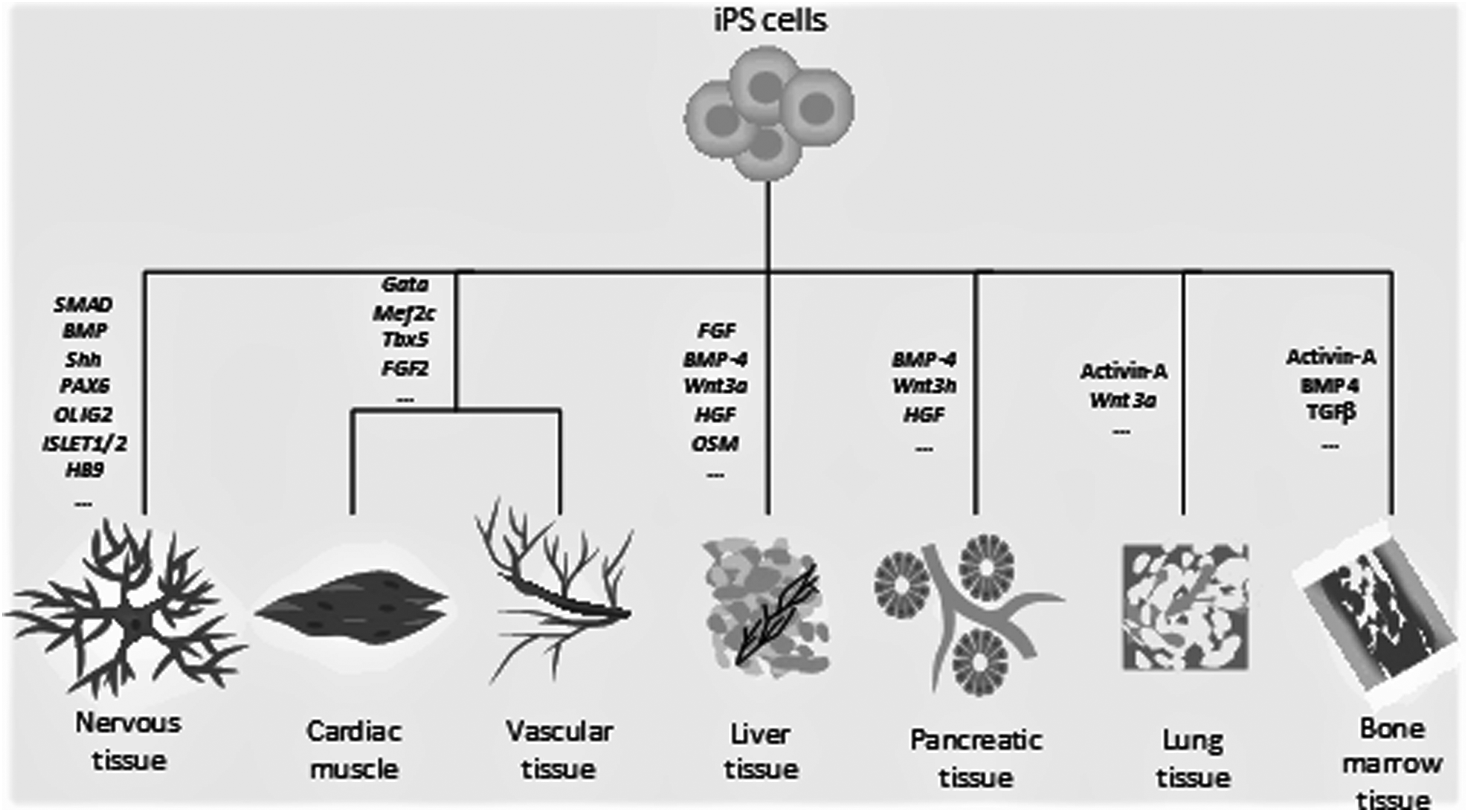
Different factors needed for iPS cell differentiation.
Currently, several diseases can be cured by organ transplantation, although lack of available organ donors causes limitations. Therefore, this medical field is in extreme need of new sources of cells and tissues for transplantation therapy (Nicolas et al., 2016). Application of iPS cells can solve the immunological rejection problem of cell transplantation process. In addition, these cells do not have the social and ethical controversies associated with hESCs (Svendsen, 2013; Teoh and Cheong, 2012).
iPS cell applications in brain and nervous system diseases
Neurodegenerative disorders pathologically are illustrated by the progressive and specific loss of highly specialized neuronal and glial populations that lead to memory, cognitive, and motor coordination defects and finally death of the patient. Among the wide range of hereditary and sporadic neurological disorders, Alzheimer's disease, Parkinson's disease (PD), Huntington's disease, amyotrophic lateral sclerosis, and spinal muscular atrophy (SMA) are the most common ones (Jung et al., 2012). These disorders are responsible for about 6.3% of the global burden of disease and as the population of the world grows older, their occurrence is anticipated to be increased (Jung et al., 2012; Peng and Zeng, 2011; Saporta et al., 2011).
The mechanisms of neuronal cell death in these diseases are not completely understood yet and therefore there are no efficient therapeutic methods for many of them. Lack of knowledge about these diseases is mainly due to lack of human models (Devine et al., 2011; Imaizumi and Okano, 2014). Despite this limitation, various studies have been performed on ESC differentiation into different types of neurons, which made it possible to follow the stages of neuron formation in vitro; early studies also demonstrated that coculture of ESCs with stromal cells (such as PA6) can be directed to form neural cells (Okada et al., 2004).
There are several techniques that efficiently convert iPS cells to desired cell lines. New techniques such as dual SMAD inhibition (through using Noggin, which restrains bone morphogenetic protein [BMP]) and SB-431542 (through blockage of transforming growth factor beta) generate neuroepithelial cells in a more powerful and specific manner (Chambers et al., 2009). Using appropriate factors such as Sonic Hedgehog (Shh) and Wnt8 (for midbrain dopaminergic neurons) (Fasano et al., 2010), retinoic acid (RA) and Shh (for spinal cord motor neurons) (Li et al., 2005), and Shh (for forebrain c-aminobutyric acid, GABA interneurons) (Liu et al., 2013) creates neuroepithelial cells that can be converted to region-specific central nervous system cells.
Depending on the specific process, this procedure lasts about 3–6 weeks. Afterward, accuracy of differentiation can be examined using a set of markers, including PAX6 (neuroepithelia), OLIG2 (motor neuron progenitors), ISLET1/2 and HB9 (motor neurons), acetylcholine transferase, and synapsin (mature motor neurons) (Karumbayaram et al., 2009).
Ebert et al. (2009) succeeded in generating iPS cells from a type 1 SMA patient and illustrated that these cells hold the ability to generate differentiated neural tissues and motor neurons in the absence of SMN1 expression. In culture medium, these cells strongly increased and produced cells with disease genotype and generated motor neurons that (compared with those derived from the unaffected control) had desired deficiencies, so they can be used as a helpful resource to model the disease and study its mechanisms and drug screening for SMA (Ebert et al., 2009).
They generated iPS cells from fibroblasts of SMA patients using lentiviral transduction systems expressing OCT4, SOX2, NANOG, and LIN28 genes. In the next step, these iPS cells were used to derive motor neurons. Immunostaining for the nonphosphorylated neurofilament SMI-32 and acetylcholine transferase indicated that these SMA motor neurons have developed normally, but they are more susceptible to degeneration than normal motor neurons (Ebert et al., 2009).They also indicated that the administration of valproic acid and tobramycin can increase the number of SMN-rich structures (gems) in the SMA iPS cells (Ebert et al., 2009; Xu and Zhong, 2013).
The relationship between the SMN gene defects and the selective loss of motor neurons was further demonstrated by Chang et al. (2011) who reported that ectopic SMN expression in five SMA iPS cell lines restored normal motor neuron differentiation and rescued the delayed neuritis outgrowth. Activation of endogenous SMN2 is another therapeutic plan since its activation seems to alleviate disease signs (Mack et al., 2014).
PD is one of the late-onset neurodegenerative disorders and is the second most common neurodegenerative disorder in the world (Devine et al., 2011). Two important characteristics of this chronic and progressive movement disorder are severe affection of midbrain dopaminergic neurons in the substantia nigra pars compacta and formation of Lewy bodies (abnormal fibrillar cytoplasmic inclusions), which usually develop in surviving neurons throughout the brain (Devine et al., 2011; Saporta et al., 2011). However, recent data on PD (especially in groups with nonmotor PD-related symptoms) indicate that other kinds of neuronal cells also are affected (Heine et al., 2011; Saporta et al., 2011).
Since there is no reliable method of treatment for PD and as affected people display the first signs of disease in sixth and seventh decades of their lives (Sundberg et al., 2013), the management of PD is an important issue in countries facing an aging population. Previous studies showed that cell replacement therapy using transplantation of fetal dopaminergic (DA) neurons in PD patients can be useful, but since fetal tissues are so limited, hESCs or hiPS cells are regarded as a substitute source for derivation of specialized DA neurons for the future cell therapy of PD (Sundberg et al., 2013).
The curative potential of iPS cells in regenerative medicine has been revealed for PD. Wernig et al. (2008) effectively produced differentiated neuronal precursor cells from patient-derived iPS cells, which had the capability to form neuronal and glial cells in vitro. Transplantation of these differentiated neuronal precursor cells into fetal rat brain led to functional and differentiated glia and neurons. More importantly, differentiated dopaminergic neurons could recover behavioral defects in rat model of PD (Wernig et al., 2008).
In modeling late-onset disease such as PD, it is necessary to not neglect effects of relevant biological, chemical, or environmental factors (Saporta et al., 2011; Xu and Zhong, 2013).
iPS cell applications in cardiovascular diseases
Heart is a mosaic organ with various types of differentiated cells, including cardiomyocytes, smooth muscle cells, fibroblasts, endothelial cells (ECs), autonomic ganglia, and pacemaker cells.
Recent studies also have revealed that there are endogenous cardiac progenitors and cardiac stem cells within an adult mammalian heart. Differentiation of these cells into vascular smooth muscle cells (VSMCs), cardiomyocytes, and ECs can be used in cardiovascular treatment using regenerative medicine (Teng et al., 2012). There are four different noncardiac stem cell sources in adults: bone marrow stem cells (both hematopoietic and mesenchymal cell lineages), skeletal myoblasts, and adipose mesenchymal stem cells (MSCs) that also have been shown to have the potential of generating cardiomyocytes in animal experiments and clinical trials (although MSC is mostly used as a cancer cell therapy tool) (Mirzaei et al., 2016; Teng et al., 2012).
Myocardial infarction, an ischemic heart disease with great losses of cardiac tissue, is one of the most common causes of death throughout the world (Segers and Lee, 2008; Zheng et al., 2016). Fortunately, medical advances significantly decreased the mortality rate of the disease, although current treatment methods do not have the ability to restore the damage to the heart. In this case, recent stem cell studies have submitted a new hope for this kind of regenerative therapy (Jang et al., 2016; Passier et al., 2008).
One of the best cell sources for cardiac tissue regeneration is ESC and the high similarity between iPSC and ESC makes it a potential replacement for ESCs in therapeutic applications. hiPS cells, generated from different somatic cell sources (mostly fibroblast cells), can be used for obtaining cardiomyocytes (Kawamura et al., 2012). Recently, other cell types have been used to generate cardiomyocytes as well, for example, human aortic VSMCs (Lee et al., 2010) and cord blood cells (Iachininoto et al., 2015).
Reprogramming of a mature cell type into cardiac cell types has been performed by introducing several transcription factors, which are often carried out using viral methods. In the first studies, miPS (mouse iPS) lines differentiated into cardiomyocytes using the Yamanaka factors (Ohno et al., 2013). It is also indicated that some other transcription factors, such as Gata4, Mef2c, and Tbx5, can be effective in more rapid and efficient generation of cardiomyocyte-like cells (Ieda et al., 2010). In these studies, iPS cell generation using Nanog revealed higher efficiency in comparison with the past studies (Ohno et al., 2013). The cardiac differentiation of hiPS cells was initially reported by Zhang et al. (2009b). The study showed that both hiPS cells and hESCs have a similar capacity for differentiation into nodal-like, atrial-like, and ventricular-like phenotypes (Zhang et al., 2009b).
iPS colonies can be differentiated into functional cardiomyocytes using several methods, which are very similar to those used for generating cardiomyocytes from hESCs. The three most frequently used methods are (1) coculture with mouse visceral endoderm-like (END-2) stromal cells (usually this strategy has low efficiency for generating cardiomyocytes) (Mummery et al., 2003), (2) spontaneous embryoid body (EB) differentiation in suspension in the presence of differentiation signals (e.g., TGF-β and Wnts), which is the most common method for generating cardiomyocytes from iPS cells (Lian et al., 2012; Ohno et al., 2013), and (3) two-dimensional monolayer differentiation that allows the differentiation of pluripotent stem cells under fully defined condition without formation of EBs, which derives specific mature cell types in large quantities (Chen et al., 2014).
BMP4 and activin A are potent factors that are used for differentiation of stem cells into cardiomyocytes (Laflamme et al., 2007). Some other inducers that have been used in cardiac differentiation of ESCs and iPS cells are trichostatin, FGF2, ascorbic acid, ITS, cyclosporin-A, and VEGF (AKaichi et al., 2010; Ye et al., 2013).
In recent years, there has been a significant increase in the number of publications on modeling cardiovascular diseases using in vitro iPS cell technology. LEOPARD syndrome (Lin et al., 2012), catecholaminergic polymorphic ventricular tachycardia (Fatima et al., 2011), and dilated cardiomyopathy (Sun et al., 2012a) are some examples of these diseases. In these diseases, fibroblast cells have been used as source of somatic cells for iPS generation. Generation of hiPS cells from patients with advanced heart failure also represented a good candidate population for future hiPS-based cell therapies (Zwi-Dantsis and Gepstein, 2012).
iPS cell applications in liver diseases
Liver is the largest internal organ in the human body with an essential role in metabolism. It also has several other special functions, including glycogen storage, plasma protein synthesis, and detoxification of toxic metabolites. Because of these important functions, it is essential to find an effective replacement for repairing its defects (Yanagida et al., 2013). Today, the only proven treatment for end-stage liver diseases is orthologous organ transplantation. However, as mentioned before, lack of donors and immunorejection are some limitations of this method (Alison et al., 2009; Liu et al., 2009). Therefore, cell-based hepatocyte transplantation is a simpler and less invasive procedure than whole organ transplantation (Liu et al., 2009) and the generation of iPS-derived hepatic cells may be effective in the treatment of severe liver diseases (Yanagida et al., 2013).
Existing protocols for generation of hepatocytes from iPS cells include using activin A for induction of endodermal progenitor cells and FGF, BMP-4 (Yanagida et al., 2013), Wnt3a (Sullivan et al., 2010), hepatocyte growth factor, and oncostatin M for differentiation toward hepatocytes (Cai et al., 2007).
Functional hepatocytes can be detected in vitro using surface asialoglycoprotein receptor expression (Basma et al., 2009).
For the first time, Sullivan et al. (2010) demonstrated that iPS cells have the ability of efficient differentiation to hepatic endodermal lineage. In following studies, these hepatic cells demonstrated to have high similarity to primary hepatocytes in gene expression profiles, secreted proteins, and metabolism (Chen et al., 2012; Liu et al., 2011). They also had the potential for long-term proliferation and differentiation to both hepatocyte and cholangiocytic cells (Yanagida et al., 2013). Afterward, these cells have been engrafted into several animal models such as rodents (Espejel et al., 2010; Chan et al., 2012). The studies demonstrated that the transplanted cells have the ability of maturation, normal functions, and treatment of the liver failure signs in animals (Espejel et al., 2010).
In the study by Espejel et al. (2010), miPSCs generated by nonviral method successfully differentiated to functional hepatocytes. In addition, a corrected point mutation in the α1-antitrypsin gene in hiPS cell-derived α1-antitrypsin-deficient liver cells has shown restoration of normal cell function (Yusa et al., 2011). Although efficient generation of hepatocytes from iPS cells has been demonstrated in many of these cases, more studies are still required for an effective and safe method to use in trial studies.
iPS cell applications in diabetes mellitus
Diabetes mellitus (DM) is a worldwide progressive disease. Already two distinct types of DM exist: T1DM with progressive β cell destruction (mostly due to autoimmunity) and T2DM mainly caused by factors involved in insulin resistance and inadequate insulin secretion (Soejitno and Prayudi, 2011). As the development of both forms of diabetes is mostly related to deficiency of pancreatic beta cells and the diabetic state can be reversed using normal beta cells, advances in stem cell biology can be effective in cell replacement and treatment of both types of DM (Weir et al., 2011).
The stem cell populations that have been used for generation of insulin-secreting cells are tissue stem cells [pancreatic stem cells (Yatoh et al., 2007), bone marrow (BM) stem cells (Hess et al., 2003), and adipose tissue-derived stem cells (Kim et al., 2010)], ESCs, and iPS cells (Tateishi et al., 2008; Zhang et al., 2009a). The first iPS-derived functional β cell was created in 2008 by Ferber et al. (2000). In this study, human dermal fibroblast-derived iPS cells could successfully differentiate into functional islet-like clusters (Ferber et al., 2000).
Stem cell is a distinct cell source for treatment of T1D patients, but because of insulin resistance and relative insulin deficiency of T2D, its treatment is more difficult. Although disease-specific hiPS cells provide a good model in investigation of new drugs for treatment of T2D in preclinical studies (Burkart et al., 2016; Liew, 2010), the efficiency of this process is still very low (Harding and Mirochnitchenko, 2014).
iPS cell applications in bone marrow failure syndromes
Recent advances in iPS cell technology and its applications in cell and gene therapy provided a new hope for treatment of inherited bone marrow failure (BMF) syndromes (the syndromes in which the production of peripheral blood cells decreases as a result of progressive decrease in hematopoietic stem cells [HSCs]). In this case, the generation of disease-free hematopoietic progenitor cells from genetically engineered reprogrammed cells may provide novel curative opportunities (Raya et al., 2009).
Fanconi anemia (FA) is the most common BMF syndrome (Shukla et al., 2012). It manifests in early childhood (first decade of life) with pancytopenia, developmental delay, increased incidence of solid tumors and leukemia, physical abnormalities (short stature and microcephaly), abnormal skin pigmentation, and other malformations (1983; Shukla et al., 2012). There are 14 known genes and their mutations play a role in FA (Muller et al., 2012; Shukla et al., 2012). Products of these genes play an important role in repair of DNA damages (1983; Shukla et al., 2012).
At the moment, administration of hematopoietic growth factors and bone marrow transplantation from human leukocyte antigen (HLA)-identical siblings are the only available therapeutic options for FA treatment. A new method also proposes to use autologous HSCs with some genetic corrections for treatment of FA (Raya et al., 2009; Shukla et al., 2012). However, the low efficiency of this process and the small number of the treated HSCs have caused uncertainty in using this method (Raya et al., 2009).
In recent years, generation of genetically corrected iPS-derived HSCs by the reprogramming of nonhematopoietic somatic cells has led to the production of large numbers of autologous HSCs that can restore the hematopoietic function in FA patients (Raya et al., 2009). In this case, Müller et al. (2012) displayed that FA-deficient somatic cells can be reprogrammed to iPS cells with a decreased efficiency (Muller et al., 2012). In contrast, Raya et al. (2009) reported that four samples of FA-A and two samples of FA-D2 patients could not be reprogrammed.
These results indicated that correction of the FA pathway is a requirement for iPS cell generation from somatic cells of FA patients (Muller et al., 2012; Shukla et al., 2012). Despite all this progress, the exact role of the FA pathway in reprogramming process remains unclear.
iPS cell applications in lung diseases
In recent years, the therapeutic potential of stem cells in clinical trials of lung diseases has been cleared (Weiss, 2014). Especially iPS cells with an unlimited potential of differentiation might provide an alternative source of stem cells for therapeutic uses (Somers et al., 2010). Since murine models of lung diseases are not often the same as human lung diseases, generation of disease-specific lung epithelial cells from hiPS cells is particularly important (Mou et al., 2012). In addition, deriving lung progenitor cells from patient-specific pluripotent cells is a key step in producing differentiated lung epithelium for disease modeling and transplantation (Mou et al., 2012; Sun et al., 2012b).
Recently, Somers et al. (2010) have generated iPS cells for a variety of lung diseases, including cystic fibrosis, alpha-1 antitrypsin deficiency-related emphysema, and scleroderma; However, due to difficulties in deriving endodermal cells from iPS cells, studying and working with this cell type are not as common as other cell types such as heart or central nervous system (CNS) (Wetsel et al., 2011). Besides, no defined condition for differentiation of iPS cells into an alveolar epithelial lineage with high homogeneity has been reported yet (Ghaedi et al., 2013).
iPS cell applications in eye diseases
Retinitis pigmentosa (RP) and age-related macular degeneration (AMD) are two irreversible severe retinal degeneration diseases that can cause visual impairment and blindness (Kamao et al., 2014; Sun et al., 2015). Although there is no effective treatment for these patients, cell therapy serves as a new hope (Jin et al., 2011; Okamoto and Takahashi, 2011). In recent years, several studies were performed on RP and AMD patients using iPS cells to either detect the exact mechanism of the diseases or find an effective cure for them. Most of these studies differentiated iPS cells into the retinal pigment epithelium (RPE) that is essential for function and survival of photoreceptor cells (Carr et al., 2009); however, direct iPS cell differentiation into photoreceptor and other retinal cells has also been observed (Carr et al., 2009; Mandai et al., 2010).
For this purpose, different combinations of factors such as Dkk1, Lefty A, FCS, activin, aFGF, bFGF, taurine, Shh, and RA are used (Mandai et al., 2010), although other methods such as using small molecules (the casein kinase I inhibitor CKI-7, the ALK4 inhibitor SB-431542, the Rho-associated kinase inhibitor Y-27632, RA, and taurine) were indicated to have a similar effect for distinct retinal cell-type differentiation from iPS cells (Osakada et al., 2009).
The results of a study performed by Buchholz et al. (2009) indicated that the RPE cells obtained from the differentiation of iPS cells, ESCs, and the fetal human RPE are functionally similar. Sugita et al. (2015) showed that secretion of TGF-β by iPS-RPE cells can result in suppression of T cell immune response in the culture medium. In the study by Carr et al. (2009), transplantation of iPS-RPE cells into the dystrophic retinal rat models resulted in the improvement of their visual function. Besides, the iPS-RPE cells were detectable until 13 weeks after transplantation (Carr et al., 2009).
Jin et al. (2011) generated iPS cells from fibroblasts obtained from five RP patients, differentiated them into photoreceptor cells, and used the resulted cells as human RP models with distinct mutations for drug testing. The study by Sun et al. (2015) on the rd1 mouse model indicated that hiPS-RPE has a more effective protection effect on photoreceptor cells and a longer survival time in comparison with the MSC and neural stem cell. The authors also believe that because of severe degenerating condition used for this study, the results can be useful for AMD condition as well (Sun et al., 2015).
Kamao et al. (2014) reported generating iPS-RPE cells for clinical applications. They generated a monolayer hiPS-RPE sheet without any artificial scaffold that had normal function, quality, quantity, consistency, and safety requirements for clinical use. Their transplantation improved the visual function of the animal models and was detectable for 9 weeks in rat, for 1 year in monkey, and for 60 weeks in mouse AMD models (Kamao et al., 2014). The study by Li and colleagues also showed the efficiency and safety of iPS-RPE cells for restoring visual function in Rpe65 rd12 /Rpe65 rd12 mice (Li et al., 2012).
Zhang et al. (2014) used iPS-derived conditioned medium (iPSC-CM) for treatment of radiotherapy-induced lacrimal gland injury, which results in impaired tear secretion and dry eyes, in the mouse model. However, MSC has shown to be the best choice for lacrimal gland injury regenerative treatment (Dietrich et al., 2016).
Conclusions
iPS cell has demonstrated the potential of generating patient-specific tissues for regenerative medicine applications (Geti et al., 2012). This cell can be generated from many accessible sources and has the ability to differentiate like an ESC. It also can be modified for each patient and there is no need to find a compatible donor to avoid immunological rejection risks.
However, before the use of iPS cells in clinical trials, several related problems must be solved, for example, finding an ideal somatic cell type for reprogramming, low efficiency of reprogramming process, lack of defined protocols for effective reprogramming and differentiation, and improvement of safety and effectiveness of iPS-derived cells (Geti et al., 2012; Harding and Mirochnitchenko, 2014). Furthermore, reprogramming human cell has also appeared to be more difficult than a murine cell (Somers et al., 2010). Considering these factors, further studies will indicate the real potential of iPS cells for use in human regenerative medicine.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.