
Abstract
Abstract
Specific transcription factors are sufficient to reprogram fully induced pluripotent stem cells or other types of cells. These findings raise the question of whether chemical molecules or proteins can replace transcription factors to alter the defined cell fate. In this study, we treated mouse skin fibroblasts (MSFs) with bone morphogenetic protein 4 (BMP4) and examined intermediate reprogramming of MSFs into stem-like cells. Putative epidermal stem cells isolated from the ventral skin epidermis of an adult mouse were used to confirm the reprogramming activity of BMP4, which increased the proliferation of these cells. After these cells formed spheroids, they were treated with BMP4 and cultured for 5 days. Following BMP4 treatment, the characteristics of these cells changed, and they expressed Oct-4 and its target transcripts Nanog, Sox2, and alkaline phosphatase. To confirm the stem cell potency of these cells, we induced their differentiation into cardiomyocytes. Stem-like cell-derived cardiomyocytes exhibited mRNA expression of cardiac mesoderm markers such as Nk2 transcription factor-related locus 5 and connexin 40, and the cardiomyocyte marker troponin T. These differentiated cells exhibited contracting masses. These results suggest that BMP4-mediated somatic stem cell reprogramming may become an alternative approach for cell therapy.
Introduction
E
Adult stem cells can be derived from different parts of the body and, depending on where they are derived from, have different properties. These cells have been isolated from several tissues such as bone marrow (Pittenger et al., 1999; Prockop, 1997; Wilmut et al., 1997), the central nervous system (Gage, 2000; Reynolds et al., 1992), skeletal muscle (Gussoni et al., 1999; Jackson et al., 1999), and the retina (Tropepe et al., 2000). Several studies suggested that adult stem cells uniquely produce differentiated cells of an identical lineage as their tissue of origin and, moreover, can develop into many different cell types (Bjornson et al., 1999; Brazelton et al., 2000; Clarke et al., 2000; Ferrari et al., 1998; Galli et al., 2000; Gussoni et al., 1999; Mezey et al., 2000; Petersen et al., 1999). Therefore, adult stem cells are not only tissue biased but also “adaptable.” This characteristic of adult stem cells may provide new insight into cellular differentiation and has potential therapeutic significance.
Although they are associated with few ethical problems and are multipotent of differentiation, adult stem cells are obtained through an invasive procedure and their numbers significantly decrease as an individual ages (Rao and Mattson, 2001). To overcome these problems and ensure a plentiful supply of cells, many new alternative trials have been performed, such as somatic cell reprogramming. There are several ways to reprogram a somatic cell into the pluripotent or precursor state such as somatic cell nuclear transfer (Byrne et al., 2003; Gurdon et al., 2003; Wilmut et al., 1997), hybridization of the somatic cell with ESCs (Cowan et al., 2005; Flasza et al., 2003; Pells and McWhir, 2004; Tada et al., 2001), and permeabilization of a cell-free extract prepared from pluripotent cells into somatic cells (Collas et al., 2006). Although these procedures reset the genome of a somatic cell to its initial state, their efficiency remains open to question.
The skin is the largest organ in the body. Adult skin consists of the epidermis, the dermis, and appendages such as hairs and glands that are linked to the epidermis, but project deep into the dermal layer. Stem cells from the epidermis and hair follicles have been well studied in the past several decades, and multipotent cell populations from dermal tissues have been identified (Janes et al., 2002; Lako et al., 2002; Toma et al., 2001; Young et al., 2001). Considering the potential application of adult stem cells in regenerative medicine, the stem cell populations in skin tissues may also have extensive therapeutic implications for the replacement of skin and may be an alternative source of stem cells for several organs other than the skin.
Several articles reported the identification of multipotent cells from adult dermis (Lako et al., 2002; Toma et al., 2001; Young et al., 2001). These cell populations have the potential to produce neural cells (neurons and glia), mesoderm derivatives (osteocytes, chondrocytes, smooth muscle cells, and adipocytes), and even blood cells. Although the origin of these multipotent cell populations remains uncertain, hair follicle dermal cells and nonfollicle dermal cells are regarded as candidates. The follicle dermal sheath may be the strict origin of these multipotent cells. However, scattered multipotent cells may also exist in the nonfollicle dermis because multipotent cells were identified in marrow stroma and adipose tissue, which have a similar mesodermal origin in embryonic development as the dermis (Prockop, 1997; Zuk et al., 2001).
Recent studies showed that mouse ESC-conditioned medium (CM) contains several biologically active interleukins, colony-stimulating factors, cytokines, and other growth-modulatory proteins, which enhance the survival of murine bone marrow myeloid progenitors (Guo et al., 2006). Goat embryonic stem-like cells remain undifferentiated when cultured in ESC-CM, indicating that mouse ESCs secrete autocrine or paracrine factors capable of maintaining pluripotency (Tian et al., 2006).
Among these factors, Wnt3a (Singla et al., 2006) and bone morphogenetic protein (BMP) 4 (Qi et al., 2004; Ying et al., 2003) are independently capable of maintaining murine ESCs in their undifferentiated state. BMPs are present in a wide variety of stem cell niches that maintain both embryonic and somatic stem cells (Li and Neaves, 2006). BMP4 acts synergistically with leukemia inhibitory factor to support ESC self-renewal through the inhibition of mitogen-activated protein kinase (MAPK) pathways and to decrease upon ESC differentiation (Palmqvist et al., 2005; Qi et al., 2004).
This study was based on two hypotheses. One is that skin, especially the epidermal region, may be a local repository of diverse adult stem cell populations, including committed and noncommitted stem cell populations. The other is that BMP4 may affect the potency of this adult stem cell population. In this study, the effect of BMP4 treatment on epidermal fibroblast-like cells was examined. The results are as follows: (1) BMP4 treatment significantly amplified the proliferation of putative epidermal stem cell populations, and (2) these BMP4-treated stem cells had a similar potency as both pluripotent and multipotent stem cells. A new simple method for generating a sphere-like organoid body (OB) was also developed that is quite different from the traditional method (Reynolds et al., 1992; Singla et al., 2006; Toma et al., 2001).
Based on the results, the adult epidermal region contains somatic stem cells, and these cells have both multipotent and pluripotent stem cell characteristics when treated with BMP4 in vitro. It is also suggested that BMP4 treatment promotes proliferation, inhibits lineage-committed differentiation, and, furthermore, alters the potency of putative epidermal stem cells to make them more developmentally potent in vitro. This could increase the therapeutic importance of somatic cells, especially those in skin.
Materials and Methods
Cell culture of adult somatic tissues
Primary cultures were prepared by somatic explant culture of tissue. Tissue was isolated from the ventral epidermal region of a 10-week-old adult mouse (Fig. 1A). The tissue was isolated under sterile conditions, washed twice in phosphate-buffered saline (PBS) containing antibiotics, cut into 2 ± 3 mm long slices, which were placed on the bottom of culture dishes (Falcon No. 3002; Becton Dickinson, Franklin Lakes, NJ), and left to dry at room temperature. About 1 hour later, a murine ESC culture medium, namely, Dulbecco's modified Eagle's medium (DMEM) supplemented with 1 mM glutamine (Invitrogen, Grand Island, NY), 1% (v/v) MEM-nonessential amino acids (NEAA; Invitrogen), 10 μM 2-mercaptoethanol (Sigma, St. Louis, MO), 100 U/mL penicillin (Invitrogen), and 0.5 mg/mL streptomycin (Invitrogen) containing 20% ESC-qualified fetal bovine serum (FBS; HyClone, Logan, UT), was gently added, taking care not to detach the ventral tissue from the bottom of each dish.
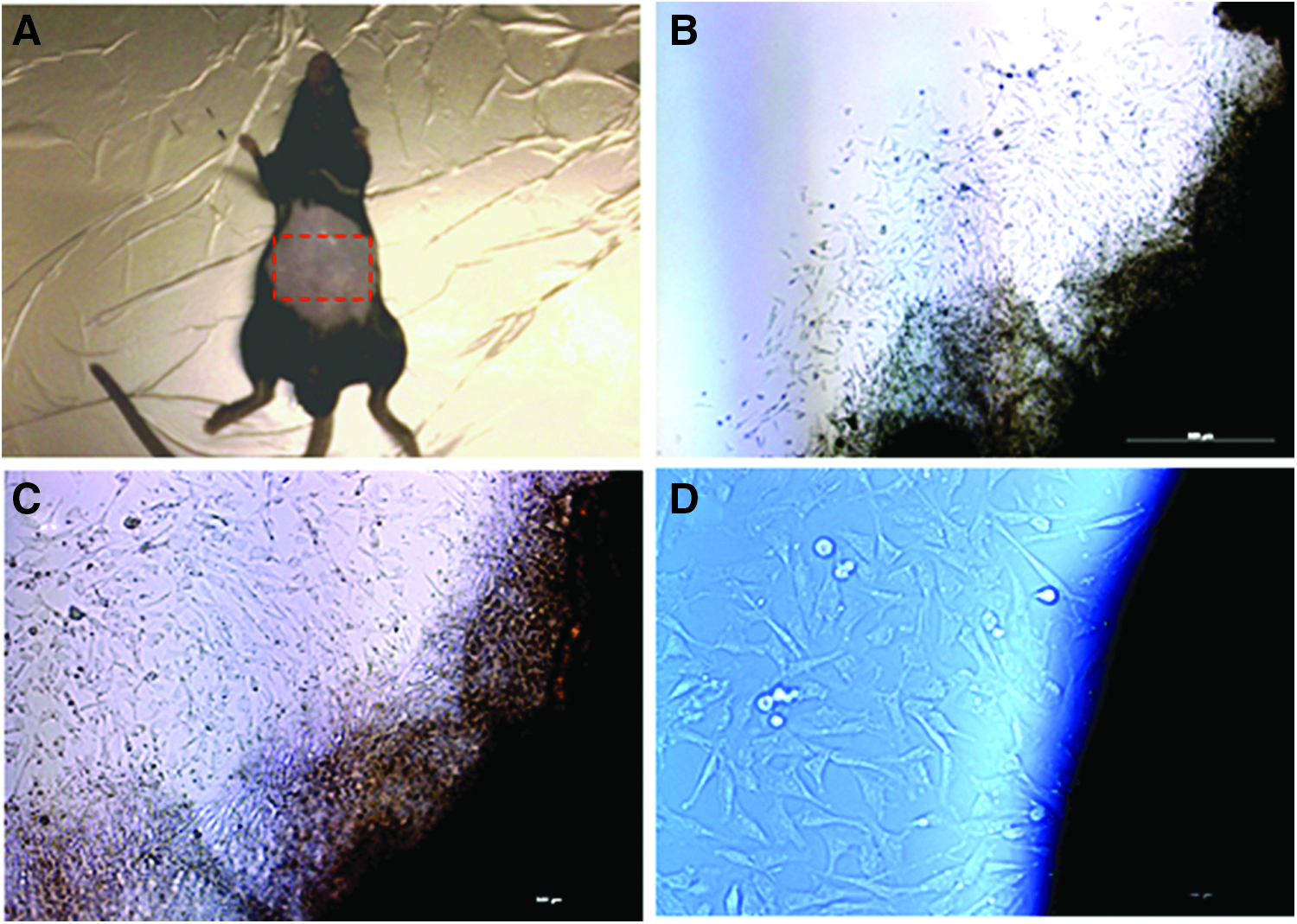
Isolation of epidermal cells from adult mouse ventral epidermis region.
The tissue was cultured in the same medium in a humidified atmosphere of 5% CO2/95% air at 37°C for 5 ± 8 days until cells developed at the bottom of the dish. To ensure a sufficient number of cells, outgrowing cells were trypsinized and subpassaged. Typically, the first outgrowing cells from the tissue explants became visible within 48–60 hours of incubation, and subpassaging was performed after 7–10 days of primary culture.
BMP4 study and staining for endogenous alkaline phosphatase activity
To determine the optimal concentration of BMP4, various concentrations were tested (0, 1, 5, 10, 15, 20, 30, and 50 ng/mL) during subculture of cells derived from the epidermis. To evaluate the differences in responses to these concentrations of BMP4, cells at two densities, low (standard) and high, were prepared in 12-well cell culture plates. Cells were cultured in the murine ESC culture medium as described above, and a specific concentration (0–50 ng/mL) of recombinant human BMP4 (R&D Systems, MN) was added. The culture medium was changed every other day. After 5 days of culture, the alkaline phosphatase (AP) activity was examined in each culture to identify the presence of stem cells and determine their numbers. AP is the most widely used stem cell marker. AP histochemistry was performed using the Sigma 86C-1 kit according to the manual provided with the kit. Briefly, cultures were washed with PBS, fixed in 80% (v/v) alcohol for 15 minutes, washed in PBS, and then stained with Fast Blue BB salt/naphthol AS-BI for 60 minutes (Sigma) at room temperature.
Sphere-like OB formation
Sphere-like OBs were prepared as described elsewhere (Reynolds et al., 1992; Toma et al., 2001) with major modifications. Briefly, to generate sphere-like OBs, cultured cells were trypsinized and pipetted gently to generate single cells and then cultured in suspension with 5 × 105 cells per 100 mm bacterial culture dish. All cells were cultured in DMEM/F12 containing 20% (v/v) knockout serum replacement (KSR; Invitrogen), 1% (v/v) MEM-NEAA, 10 μM 2-mercaptoethanol, 100 U/mL penicillin, 0.5 mg/mL streptomycin, and 30 ng/mL BMP4 for 24 hours. At 24–36 hours of culture, the AP activity was examined.
Embryoid body formation and cardiac cell differentiation
For cardiac cell differentiation, to generate the sphere-like OBs, cultured cells were collected and cultured in DMEM containing 20% (v/v) FBS (HyClone), 1% (v/v) MEM-NEAA, 10 μM 2-mercaptoethanol, 100 U/mL penicillin, and 0.5 mg/mL streptomycin. After 4 days, small embryoid bodies (EBs) were transferred to the Microvascular Endothelial Cell Basal Medium (EBM-2; Lonza, Walkersville, MD) and cultured for 4 additional days. On day 8, cystic EBs were plated onto 0.1% gelatin-coated tissue culture dishes and allowed to differentiate in DMEM containing 20% (v/v) FBS (HyClone), 1% (v/v) MEM-NEAA, 10 μM 2-mercaptoethanol, 100 U/mL penicillin, and 0.5 mg/mL streptomycin for 15 days.
Immunofluorescence
Immunocytochemistry was performed of cells plated on a gelatin-coated coverslip inserted in a culture dish. To stain for Oct-4, stage-specific embryonic antigen-1 (SSEA-1), and CD90, cells were fixed in 4% paraformaldehyde for 30 minutes at room temperature, and then rinsed with 1 × PBS before being permeabilized with 0.1% Triton X-100 for 20 minutes at room temperature. Following permeabilization, cells were rinsed with 1 × PBS and blocked in nonimmune goat serum or 12% bovine serum albumin for 1 hour at room temperature. Cells were then incubated with the following primary antibodies: monoclonal anti-Oct-4 (Chemicon, Temecula, CA), monoclonal anti-SSEA-1 (Chemicon), and monoclonal anti-CD90 (Chemicon). Primary antibodies were localized using an FITC-conjugated IgG secondary antibody. Samples were mounted on the VECTASHIELD mounting medium containing 1.5 μg/mL DAPI (Vector Laboratories, Burlingame, CA) and observed under an Olympus BX 61 fluorescence microscope.
RNA isolation and reverse transcription–polymerase chain reaction
Total RNA isolated from cells using TRIzol® (Invitrogen) was converted into cDNA using oligo dT and RT PreMix (Bioneer, Daejeon, Korea). Polymerase chain reaction (PCR) was performed with recombinant Taq polymerase (TaKaRa Bio, Inc., Otsu, Shiga, Japan). The cycling parameters were as follows: denaturation at 94°C for 30 seconds, annealing at 55°C for 30 seconds, and extension at 72°C for 30 seconds. PCR products were separated on 1.5% agarose gels and visualized by ethidium bromide staining. Amplification of the gene encoding glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control. The following primers were used to amplify the genes: mouse Oct-3/4, sense 5′-ACCCAGGCCGACGTGGGGCT-3′ and antisense 5′-TTC TGGCGCCGGTTACAGAACCA-3′ (365 bp); mouse Nanog, sense 5′-TACCTCAGCCTCCAGCAGAT-3′ and antisense 5′-CCTCCAAATCACTGGCAG-3′ (460 bp); mouse Nestin, sense 5′-GTCTTCACCACCATGGAGAAG-3′ and antisense 5′-CAAGGGGGAAGAGAAGGATGTCG-3′ (694 bp); mouse Sox2, sense 5′-TTCGGTGATGCCGACTAGA-3′ and antisense 5′-TGCGAAGCGCCTAACGTA-3′ (100 bp); mouse AP, sense 5′-TTGTGCCAGAGAAAGAGAGAGA-3′ and antisense 5′-GTTTCAGGGCATTTTTCAAGGT-3′ (453 bp); mouse Nk2 transcription factor-related locus 5 (Nkx2.5), sense 5′-CAAGTGCTCTCCTGCTTTCC-3′ and antisense 5′-CGGCTTTGTCCAGCTCCACT-3′ (137 bp); mouse connexin (Cx) 40, sense 5′-GGTCTGGCTCACTGT CCTGT-3′ and antisense 5′-GAAGGCGTGGACACAAA GAT-3′ (198 bp); mouse troponin T (TnT), sense 5′-GAGG AACAGGAAGAGCAAGA-3′ and antisense 5′-ACCAAGT TGGGCATGAAGAG-3′ (191 bp); and mouse GAPDH, sense 5′-GTCTTCACCACCATGGAG AAG-3′ and antisense 5′-GCCATCCACAGTCTTCTG GGT-3′ (270 bp).
Western blot analysis
For Western blot analysis of BMP receptor type 1 (BMPR1), BMP receptor type 2 (BMPR2), Smad, and Oct-4, cells were lysed for 60 minutes at 48°C in a lysis buffer (50 mM Tris-HCl, pH 7.4, 1% Triton X-100, 150 mM NaCl, 5 mM EDTA, 0.2 mM sodium orthovanadate, 0.2 mM sodium fluoride, 1 mM phenylmethylsulfonyl fluoride, 1 mg/mL leupeptin, and 1 mg/mL aprotinin). The lysates were clarified by centrifugation (12,000 g for 10 minutes at 48°C), and the total protein concentration in the supernatant fraction was quantified by the modified Bradford protein assay (Bio-Rad, Hercules, CA).
For immunoblotting, equal amounts of proteins were separated by SDS-PAGE and transferred to nitrocellulose membranes (Amersham Biosciences, Piscataway, NJ) using a semidry transfer apparatus (Bio-Rad). The membranes were blocked in 20 mM Tris, pH 7.5, 150 mM NaCl, and 0.1% Tween-20 containing 5% skimmed milk powder. The primary antibodies were monoclonal anti-Nestin (Chemicon), monoclonal anti-neurofilament M (Chemicon), and, as a loading control, monoclonal anti-α-tubulin (Sigma). Immunoreactive bands were detected by enhanced chemiluminescence (ECL Plus Western blotting kit; Amersham Biosciences) using an anti-mouse IgG horseradish peroxidase-conjugated secondary antibody (Chemicon).
Statistical analysis
All experiments consisted of at least three replicates. Statistical analysis was performed using SAS software (Statistical Analysis System, Inc., Cary, NC). The means of developmental data and nuclei counts were compared by the Student's t-test or analysis of variance where appropriate. Data are presented as mean ± standard error of the mean. p < 0.05 was considered significant.
Results
Isolation of cells from adult mouse skin epidermis
To prevent putative stem cell loss and isolate primary cells from the skin directly, adhesive culture of a small piece of epidermal explant was performed in the murine ESC culture medium because enzymatic digestion of skin tissue may lead to loss or damage of putative stem cells residing in the epidermis, and a low amount or no serum supplementation might lead to progressive loss of characteristics of putative stem cells. Several small follicle-like outgrowing cells became visible around the explants after 24 hours of incubation. These follicle-like cells became fibroblast-like cells and developed at the bottom of the dish within 60–72 hours (Fig. 1B–D). Cells were harvested after 7–10 days of primary culture, and sufficient numbers of cells were obtained after one to two subcultures.
Measurement of AP activity following treatment of cells derived from the epidermis with different concentrations of BMP4
The proportion of AP-positive cells was gradually increased by BMP4 treatment in a dose-dependent manner both in low-density (standard, Fig. 2A) and high-density (Fig. 2B) cultures, and was highest for cells treated with 30 and 50 ng/mL BMP4. However, the increase was not significant for low-density cultures, in contrast to high-density cultures. This result revealed that stem cells exist in the epidermis and BMP4 treatment might affect cell proliferation and characteristics. The AP activity was dramatically increased by BMP4 treatment in a dose-dependent manner (Fig. 2C).
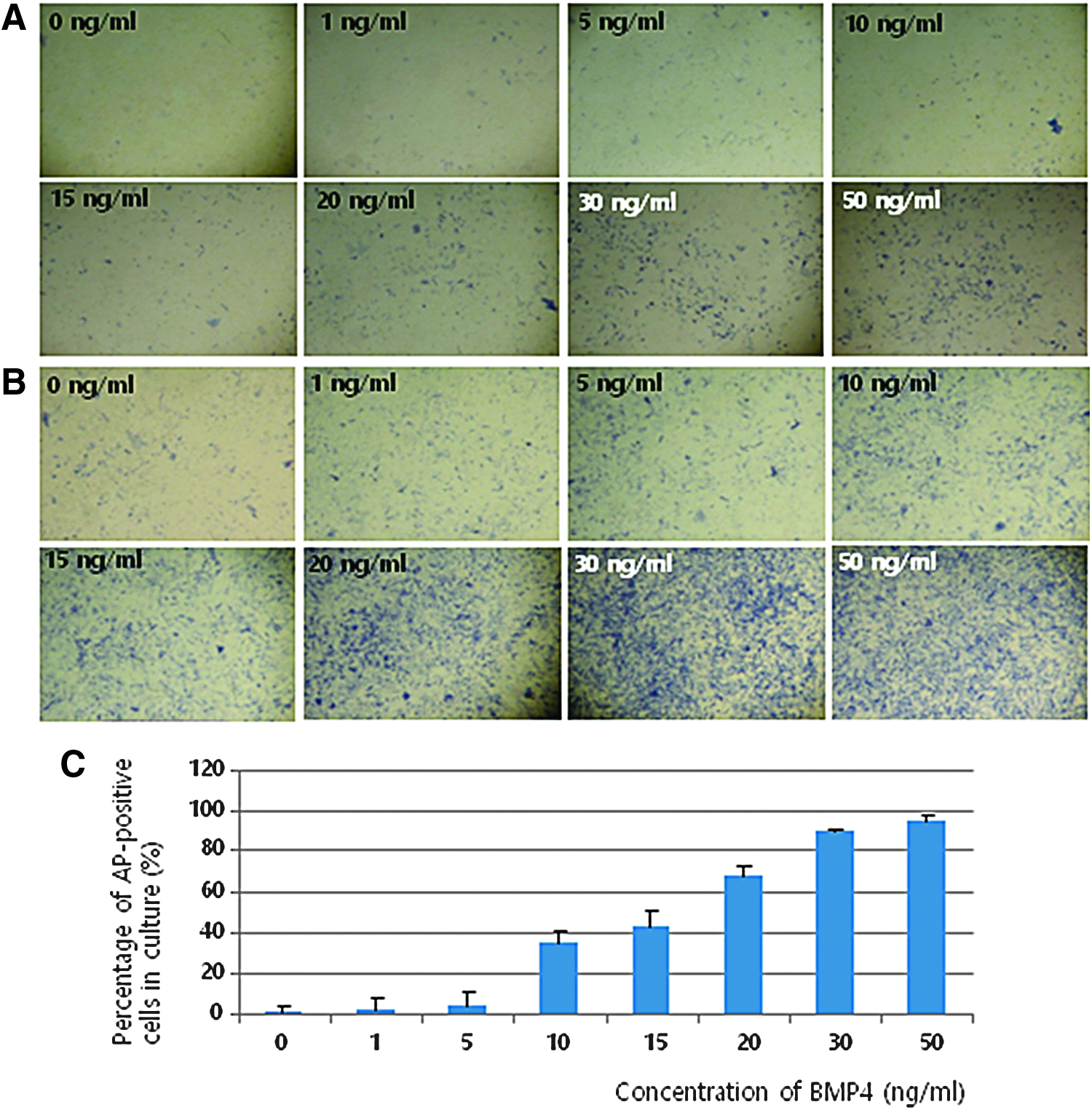
Measurement of AP activity following treatment of cells derived from adult mouse epidermal tissue with various concentrations of BMP4. The percentage of AP-positive cells was gradually increased by BMP4 treatment in a dose-dependent manner, both in low-density
Although the percentage of AP-positive cells was highest for samples treated with 50 ng/mL BMP4, treatment with 30 ng/mL BMP4 was used in subsequent experiments because there was not a large difference in the percentage of AP-positive cells between samples treated with 30 and 50 ng/mL BMP4 (89.2% and 95.2%, respectively), in contrast to samples treated with 20 and 30 ng/mL BMP4 (67.2% and 89.2%, respectively). Grinnell and Bickenbach (2007) reported that BMP4 is present at a concentration of 24.6 ng/mL in mouse ESC-CM, as determined by Western blotting. Using this CM, they induced expression of embryonic genes in cultured keratinocytes. This also influenced our decision regarding the optimal BMP4 concentration for treatment.
Generation of sphere-like OBs
Cultured fibroblast-like epidermal cells were dissociated by enzymatic digestion. The percentage of AP-positive cells was dramatically increased by treatment with BMP4 at concentrations higher than 10 ng/mL (p < 0.05). Using 0.25% trypsin-EDTA, dissociated cells were grown in a serum-free basal culture medium (DMEM/F12 containing 20% KSR) containing 30 ng/mL BMP4 for 24 hours to generate free-floating sphere-like OBs. This system differs from the conventional procedure reported previously, in which cells are cultured in DMEM/F12 containing B-27, 20 ng/mL epidermal growth factor (EGF), and 40 ng/mL basic fibroblast growth factor (bFGF) for 3–7 days (Toma et al., 2001).
After 24 hours of culture, a number of free-floating sphere-like OBs were observed with only a few cells in the medium. Various sizes (smaller than 10 μm to larger than 100 μm) of free-floating sphere-like OBs were distinctly generated and observed at this time point (Fig. 3A, B). However, some small OBs attached to the surface of the dish and budded. Only OBs smaller than 100 μm were positively stained for AP activity (Fig. 3C).
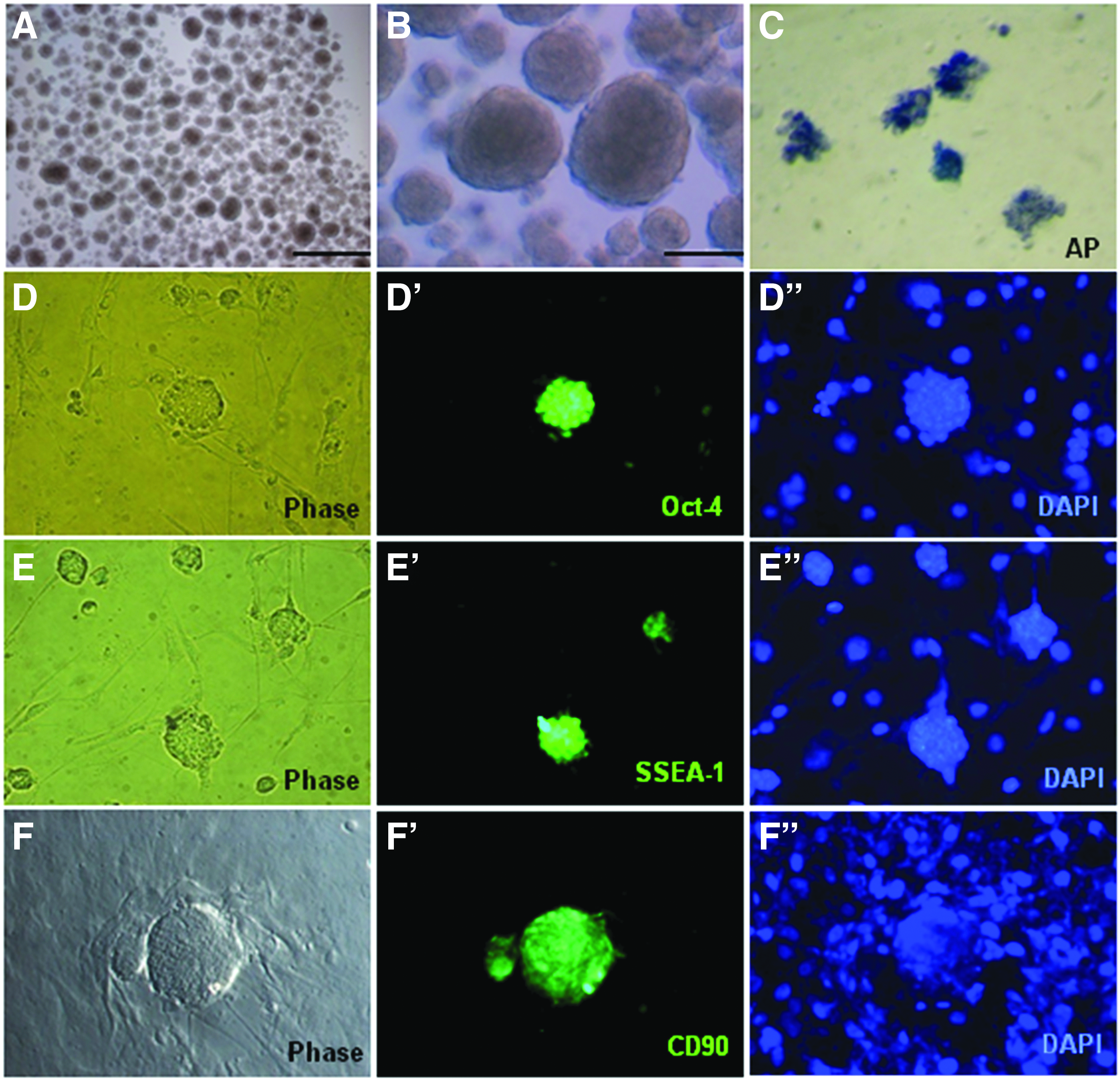
Formation of sphere-like OBs from single, putative epidermal stem cells isolated from adult mouse epidermis.
Characteristics of sphere-like OBs
To determine the characteristics of free-floating sphere-like OBs, immunocytochemistry was performed first. OBs cultured for 24–36 hours were collected and fixed, and their expression of several markers was examined. OBs of 50–100 μm in diameter expressed Oct-4 (ESC marker), SSEA-1, and CD90 (mesenchymal marker) (Fig. 3D–F"), but those larger than 100 μm did not express all these markers. This means that OBs expressed mesenchymal markers and BMP4 treatment stimulated and increased embryonic gene expression in OBs smaller than 100 μm. It is thought that it is difficult for BMP4 stimulation to be sufficiently relayed to the core region in OBs larger than 100 μm, meaning increased embryonic gene expression is rapidly reduced in such OBs.
To confirm the immunocytochemical results, mRNA expression of genes associated with pluripotent and multipotent stem cells was examined in cells and OBs. BMP4-treated cells and OBs expressed stem cell transcription factors such as Oct-4, Nanog, AP, Nestin, and Sox2 (Fig. 4A). All examined transcription factors were expressed in both cells and OBs; however, expression was much higher in OBs than in cells. These results imply that continued treatment of mouse epidermal cells with BMP4 results in transcriptional changes, such as inhibition or downregulation of genes involved in the lineage-committed differentiation of epidermal cells, and stimulation of multipotent and even pluripotent gene transcription.

Effects of recombinant BMP4 treatment on putative epidermal stem cell gene expression.
To address whether BMP signaling components were affected and analyze the intracellular signaling pathway, Western blot analysis of both cells and OBs was performed. First, BMP receptors were examined to determine whether BMP4-treated epidermal cells and OBs have such receptors, and then Smad, a downstream signaling effector protein, was examined. TGF-β proteins cannot enter living cells, and instead attach to receptor proteins on the cell's outer surface, and the signal generated by this interaction is transferred across the cell membrane, through the cytoplasm, and into the nucleus by Smad proteins. BMPR1, BMPR2, and Smad proteins were detected in both cells and OBs (Fig. 4B).
Second, Oct-4 was examined to determine whether this protein was synthesized in BMP4-treated epidermal cells and OBs. This was performed because BMP4 signaling is mediated through Smad proteins, which may translocate to the nucleus and activate target gene transcription, meaning Oct-4 could be a target gene of BMP4 signaling in somatic cells. Oct-4 was detected in both cells and OBs by Western blot analysis (Fig. 4B). This finding suggests that the potency of somatic cells can be changed by continued exogenous BMP4 treatment in ESC culture conditions.
Cardiac cell differentiation of BMP4-treated cells
For cardiac cell differentiation, BMP4-treated cell-derived EBs were formed in suspension culture for 8 days and then plated onto gelatin-coated dishes after 1 or 2 days. Cells were then differentiated for an additional 15 days. BMP4-treated cells exhibited EB growth. During differentiation from day 8 to 15, contracting areas were observed in EBs of BMP4-treated cells (Fig. 5A). On day 10 of differentiation, gene expression in contracting EBs derived from BMP4-treated cells was analyzed (Fig. 5). Cardiac mesoderm (Nkx2.5 and the atrial marker Cx40) and cardiomyocyte-specific (the cardiac muscle marker TnT) markers were expressed in cells surrounding beating masses and beating masses in a pattern similar to that in control mouse heart cells (Fig. 5B). Expression of Nkx2.5, Cx40, and TnT was upregulated in beating masses in comparison with that in cells surrounding beating masses.
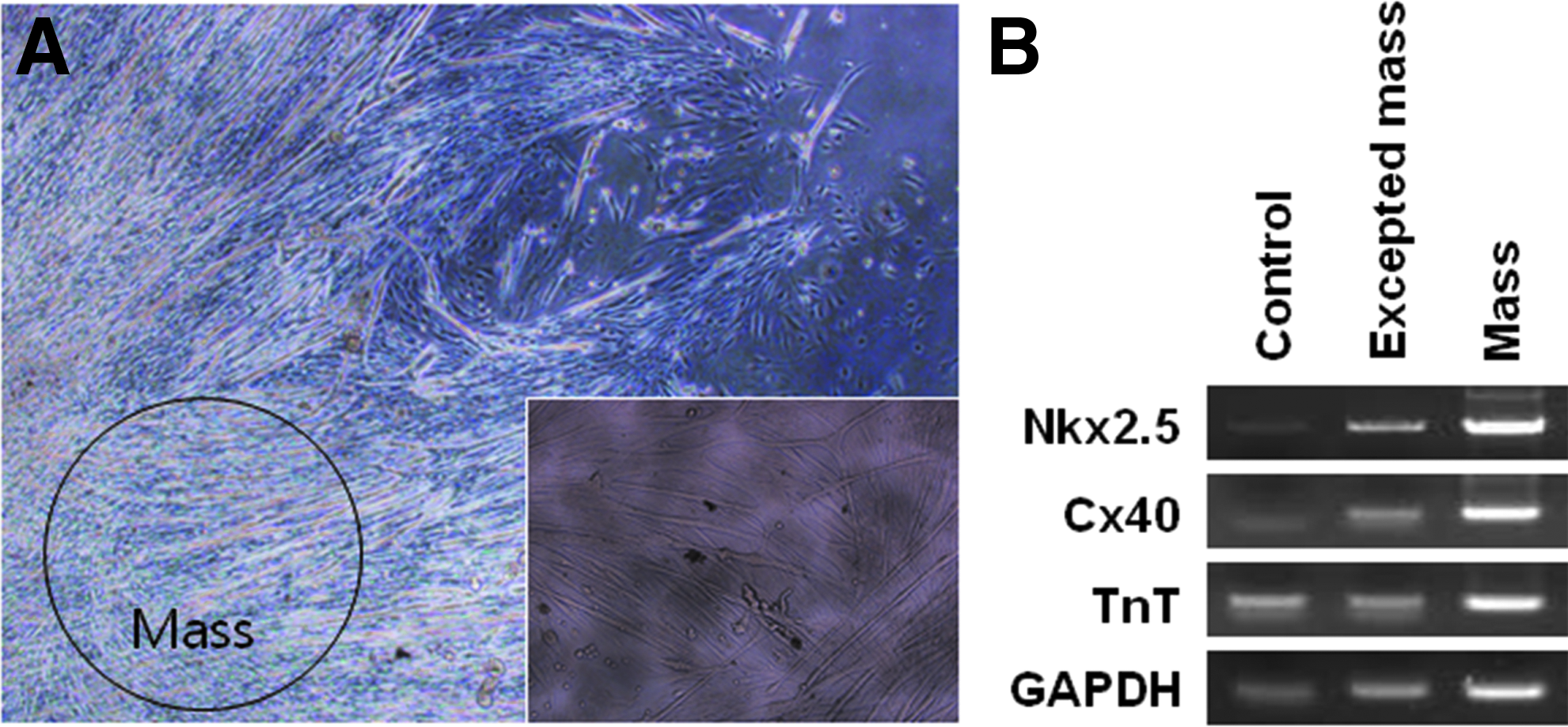
Differentiation of cells into cardiomyocytes on day 10. Differentiated cells exhibited contracting masses
Discussion
Through a series of experiments, there are three major conclusions of this study. First, putative adult stem cells (epidermal stem cells) exist in adult skin, especially the epidermis. During the last decade, many types of adult tissues have been investigated, such as the scalp (Hoogduijn et al., 2006), hair follicles (Gurdon et al., 2003; Tiede et al., 2007; Yu et al., 2006), skin dermis (Fuchs, 2007; Kawase et al., 2004; Nishimura et al., 2002; Shi and Cheng, 2004; Toma et al., 2001; Young et al., 2001), pancreas (Kruse et al., 2006), foreskin (Toma et al., 2005), term placenta (Yen et al., 2005), ear skin (Yang et al., 2007), adipose tissue, and periodontal ligament (Zuk et al., 2001), to identify adult stem cell reservoirs in mammals.
Among them, skin is the most accessible tissue because it is the largest organ of the body and differs from other tissues. According to various studies, skin-derived precursors or stem cells have various features of multipotent stem cells. Moreover, skin has the obvious advantage of being associated with fewer ethical problems than other tissues. Thus, skin was used in this study.
Second, treatment with an optimal concentration of BMP4 combined with murine ESC culture conditions markedly improved the proliferation of skin-derived stem-like cells in vitro. At the same time, in these conditions, the potency of skin-derived stem cells, which are programmed to differentiate in vivo, was altered such that they became multipotent or even pluripotent. In addition, using the simple method of adding BMP4 to the culture medium, which is considerably different from the conventional method, sphere-like OBs were generated within a short amount of time. These OBs had the same characteristics and differentiation potential as BMP4-treated cells.
For this, KSR was used instead of FBS. KSR does not contain serum; therefore, it is postulated that KSR may not be appropriate for anchorage-dependent culture and the exact action of BMP4 can be elucidated using KSR, which lacks the BMP-like factor that is present in FBS (Kodaira et al., 2006). Based on this assumption, KSR was successfully used for OB formation. Although KSR is not a chemically defined product, it has been successfully used for human ESC maintenance. A very recent study suggests that albumin-associated lipids, rather than albumin apoprotein in KSR, stimulate human ESC self-renewal (Garcia-Gonzalo and Izpisua Belmonte, 2008). Therefore, it is thought that BMP4 and albumin-associated lipids of KSR alter and maintain cell potency, respectively, in culture.
Third, it seems that the Smad pathway participates in transcriptional and differentiation potential changes in adult somatic cells induced by BMP4 treatment, to some extent. A similar case was reported for Smad7 in hair follicles (Han et al., 2006). BMP4 treatment facilitates and sustains the self-renewal of murine ESCs, during which p38 MAPK pathways are inhibited and Smad pathways are activated, leading to the inhibition of differentiation (Id) genes (Qi et al., 2004; Ying et al., 2003).
Similar to the results of this study, Grinnell and Bickenbach (2007) reported that CM of mouse ESCs or BMP4-pretreated skin keratinocytes alters the lineage-specific potential and BMP4 activates embryonic gene expression through the MAPK and Smad pathways. However, the authors insisted that BMP4 is not the only factor responsible for reprogramming of keratinocytes and several ESC-secreted factors are involved in expression of Oct-4 (Grinnell and Bickenbach, 2007).
This differs from the results of this study. Because FBS was used for primary culture and part of the subculture of epidermal cells, the transcription of Oct-4 and other embryonic genes was sustained in BMP4-treated cells and OBs cultured using KSR instead of FBS. Based on these results, BMP4 is suggested to be a reprogramming factor of mouse putative epidermal stem cells.
In this study, we confirmed the appearance of beating EBs derived from BMP4-treated cells on day 8, and the majority of contracting EBs was produced from day 8 to 15. The process of cardiomyocyte differentiation includes the emergence of a specific pattern of gene expression that regulates myofibrillogenesis, ion channel development, and cellular function (Zhou et al., 2009). In BMP4-treated cell-derived EBs, expression of Nkx2.5, Cx40, and TnT was higher in beating cell masses than in control heart cells. Nkx2.5 is expressed in both the first and second heart field (Buckingham et al., 2005). Nkx2.5-null mice exhibit an abnormal cardiac looping phenotype (Imajoh-Ohmi et al., 1995), indicating that Nkx2.5 is essential for cardiac development.
Therefore, Nkx2.5 is a cardiac-specific transcription factor that regulates cardiac development and heart diseases (Reamon-Buettner and Borlak, 2010). Gap junctions in mammalian atrial cardiomyocytes are mainly composed of Cx40 and Cx43. Heart failure increases susceptibility to atrial fibrillation and is associated with altered cardiomyocyte Cxs. Changes in atrial gap junctions and Cxs in cardiac disorders were reported previously (Kostin et al., 2002; Polontchouk et al., 2001; van der Velden et al., 1998, 2000; Wetzel et al., 2005; Yeh et al., 2001). The high level of Cx40 expression indicates the formation of robust gap junctions between BMP4-treated cell-derived cardiomyocytes. The troponin complex is composed of three elements: troponin I, troponin C, and TnT. Troponin I expression predominates throughout embryonic, fetal, and cardiac development. TnT expression was detected in cells differentiated from BMP4-treated cells. This study shows that treatment with BMP4 significantly influences the cardiac differentiation of mouse skin fibroblasts (MSFs).
In summary, it was demonstrated that BMP4 treatment enables reprogramming of somatic cells and these somatic cells have an altered differentiation potential, with the ability to differentiate into other cell lineages. These results demonstrate that cardiomyocytes can be generated from MSFs and suggest that these cells are useful for the treatment of cardiovascular disease. This finding may contribute to understanding of somatic cell reprogramming and cell therapies in the future.
Footnotes
Acknowledgment
Cooperative Research Program for Agriculture Science and Technology Development, RDA, Republic of Korea (Grant No.: PJ009103).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.