
Abstract
Abstract
MicroRNAs (miRNAs) are posttranscriptional regulators of gene expression. They play an important role in various cellular processes such as apoptosis, differentiation, secretion, and proliferation. Embryonic stem cells (ESCs) are derived from the inner cell mass of the blastocyst stage of the embryo. miRNAs are critical factors for the self-renewal and differentiation of ESCs. In this review, we will focus on the role of miRNAs in the self-renewal and directional differentiation of ESCs. We will present the current knowledge on key points related to miRNA biogenesis and their function in ESCs.
Introduction
E
MicroRNAs (miRNAs) are endogenous noncoding small RNAs that contain 19–25 nucleic acids; they are present in most animals and plants and highly conserved during evolution. Their expression is not only space and time specific but also tissue and cell specific. miRNAs are extensively involved in the regulation of cell proliferation, differentiation, secretion, and apoptosis as well as other life activities. They exert their effects by acting on the 3′-untranslated region of target miRNAs and degrading it or inhibiting its transcription (Kim et al., 2009).
Research on the key enzymes Dicer and DGCR8, which are involved in producing miRNAs, has indicated that miRNAs play an important role in ESC differentiation. The role of miRNAs in ESC differentiation, including the mechanism and the specific effects of miRNAs, has also been studied. These effects involve changes in the cell morphology and phenotype of human mesenchymal stem cells before and after induction, and these studies have provided an experimental model for the detection of miRNAs and their methylation before and after induction, as well as gene chip technology for analyzing the associated genetics (Zhao et al., 2013). In this study, we summarize the research on miRNAs regarding their role in ESC differentiation to clarify the mechanisms involved in ESC cell renewal and directional differentiation.
Key Biosynthetic miRNA Factors Participate in the Self-Renewal and Differentiation of ESCs
After an RNA product is transcribed in the cell nucleus, RNA polymerase II can process it (including 5′-capping and poly-adenylation at the 3′-end) to form pri-miRNAs (primary transcripts of miRNA genes). Then, the product will undergo shearing twice. The first shearing occurs in the cell nucleus and then it is sheared into pre-miRNAs (precursor miRNAs) of 60–70 nucleotides by a microprocessor consisting of the RNase III enzymes Drosha and DGCR8. Drosha catalyzes enzymatic digestion, and DGCR8 is involved in localization. For the second shearing, pre-miRNAs are transported out of the cell nucleus with the help of the Exportin-5 and Ran-GTP proteins and then sheared into a double helix structure of 22 nucleotides by the RNase III enzyme Dicer (dcr) in the cytoplasm. The double helix is then uncoiled by RNA double-strand helicase. One of double strands combines with the RNA-induced silencing complex to perform its gene silencing function, and the other one is degraded (Sartipy et al., 2009).
Dicer
Under normal conditions, ESCs first form a cell mass, which is called the embryoid, and can differentiate into ectoderm, mesoderm, and endoderm under the regulation of a series of molecular factors. The role of Dicer in the self-renewal and differentiation of ESCs was demonstrated in mouse ESCs in which Dicer was knocked out. The results showed that DCR1−/− ESCs have severe defects in differentiation both in vitro and in vivo, indicating that Dicer participates in biological processes and differentiation of mammalian stem cells (Kanellopoulou et al., 2005).
However, an in vivo experiment indicated that ESCs without Dicer can form embryoids. The homozygous DCRΔ/Δ mutant stopped growing after culturing for 8–10 days and did not form the three germ layers or express marker genes; however, the heterozygous DCRneo/− and wild-type DCRneo/neo continued to grow and then differentiate into the three germ layers and express marker genes. An in vivo study produced similar results, in which an embryoid analogue formed by DCRΔ/Δ ESCs did not exhibit any indication of differentiation, and this defect was repaired after Dicer was added. These experiments demonstrated that Dicer does not affect the survival of ESCs, but plays an important role in differentiation. As Dicer plays an important role in producing miRNAs, the miRNAs processed by it may participate in differentiation and survival. In addition, Dicer is also involved in the maturation of miRNAs and siRNAs (short interfering RNAs), but the influence of miRNAs on ESCs was not directly demonstrated (Lee et al., 2004).
DGCR8
DCGR8 cooperates with the RNase III enzyme Drosha for biogenesis of pre-miRNA. DCGR8 is transcribed from the noncoding regions of the genome in ESCs. Cells with DCGR8 knocked out remain in the G1 phase of the cell cycle. However, these DGCR8-knockout ESCs still survive (Sladitschek and Neveu, 2016). Furthermore, the DGCR8 gene plays an important role in the suppression of ESC self-renewal, which normally occurs with the induction of differentiation. The noncanonical miRNAs miR-320 and miR-702 promote Dgcr8-deficient ESC proliferation (Kim and Choi, 2012). The miR-290 family and the methyltransferase Ash1 also reprogram polycomb-mediated gene silencing in ESCs (Kanellopoulou et al., 2015). Wang et al. (2007) performed Dgcr8 gene knockout experiments in ESCs and reported that various miRNAs in ESCs were downregulated after knockout of Dgcr8, indicating that Dgcr8 is essential for the biosynthesis of miRNAs.
In contrast to the phenotype of ESCs without Dicer, ESCs without DGCR8 do not show full downregulation of totipotency markers during induced differentiation; in addition, a fraction of the ESCs retains the ability to form a cell colony, but also express some markers related to differentiation (Wang et al., 2007). Although DGCR8 has specificity for the synthesis of miRNAs, it has been concluded that Dicer may affect the self-renewal and differentiation of ESCs independent of miRNAs (Wang et al., 2007).
Specific miRNAs Regulate the Self-Renewal and Differentiation of ESCs
Computational and high-throughput studies indicate that a single miRNA can interact with hundreds of target genes. miRNAs can induce widespread changes in protein synthesis (Selbach et al., 2008). Moreover, most major human mRNAs are regulated by miRNAs, and some are conserved targets of miRNAs (Friedman et al., 2009). Studies have increasingly found that ESC-specific miRNAs are responsible for the initiation of ESC differentiation and normal development. Many miRNAs are specifically expressed in ESCs, and some of them show increased or decreased expression following differentiation (Chen et al., 2007; Houbavuy et al., 2003; Ren et al., 2009). The differential expression of miRNAs can regulate specific target proteins at the posttranscriptional level and then regulate the differentiation of ESCs, thus further determining the destiny of cells.
The miR-290-295 and miR-302 cluster
The miR-290-295 cluster is homologous with the miR-371-373 cluster in human ESCs, and its encoding genes are distributed in 1050 ALK gene sequences on chromosome 19. The ESC-specific miR-290-295 cluster is highly and specifically expressed in ESCs and accounts for ∼50% of the miRNA population in mouse ESCs (Leung et al., 2011; Zheng et al., 2011). Three miRNAs, namely, miR-291a-3p, miR-294, and miR-295, are coexpressed with the miR-290-295 cluster, and they are sufficient to force a G1→S transition (Lee et al., 2016).
Expression of these miRNAs decreases rapidly in stem cells during differentiation and almost cannot be detected in mature cells (Kim et al., 2009). This cluster functions to maintain the self-renewal of ESCs and inhibit their differentiation. To determine the participation of this cluster in maintaining self-renewal and inhibiting the differentiation of ESCs, Zovoilis et al. (2009) measured the expression of the marker Brachyury in mesoderm and Dppa-3 in primordial germ cells (PGCs). They found that this the miR-290-295 cluster can inhibit the differentiation of ESCs into mesoderm and germ lines and it may act on the inhibitor Dkk-1, whose target site is the Wnt signaling pathway. Some reports indicated that this cluster inhibits Dnmt3a and Dnmt3b by acting on Rbl2 and others concluded that the inhibitory effect is related to the methylation of DNA (Sinkkonen et al., 2008). In addition, binding sites for Oct4, Sox2, Nanog, and Tcf3 are found in the miRNA sequences of the miR-290-295 cluster; therefore, this cluster may be controlled by these transcription factors (Kim et al., 2009).
Similar to the miR-290-295 cluster, miR-302 has been used to induce fibroblasts into pluripotent stem cells (Anokye-Danso et al., 2011). The miR-302 cluster is homologous with the miR-302 cluster in human ESCs and its encoding genes are distributed in 700 ALK gene sequences on chromosome 4. Members of the miR-302 cluster are highly conserved and have high homology (although miR-367 is distinctly different). The miR-302 cluster is highly expressed in ESCs and its expression decreases after ESC differentiation (Rosa et al., 2009) or increases first and then decreases (Gangaraju and Lin, 2009). The miR-302 cluster may maintain the omnipotency of ESCs by regulating Oct4 and cyclinD2 (Kim et al., 2009), thus regulating the determination of early cell fate.
Because the miR-302 cluster has the same seed sequence as the miR-290-295 cluster, it has been suggested that this cluster can also inhibit the differentiation of ESCs into mesoderm and germ lines (Zovoilis et al., 2009). However, Rosa et al. (2009) showed that because this cluster can increase the activity of Nodal by inhibiting Lefty, it can maintain the normal differentiation of the mesoderm and endoderm, while also inhibiting the differentiation of the neuroderm. The miR-302 cluster has also been reported to be involved in determining sex differentiation (Ciaudo et al., 2009). The expression of this cluster may be regulated by Oct4, Sox2, Nanog, Tcf3, and Rex1 (Kim et al., 2009).
miR-1 and miR-133
In mice, the miR-1 and miR-133 clusters are initially transcribed into a single continuous RNA molecule, which is then cleaved into two separate miRNA precursors. miR-1 and miR-133 are muscle-specific miRNAs and are highly expressed in skeletal muscle and cardiac muscle cells, and they function in regulating the differentiation of ESCs into cardiac muscle cells (Ivey et al., 2008; Tao et al., 2015). Expression of miR-133 has also been detected in several nonmuscle tissues, such as complex neural network. pri-miR-1/miR-133 can be amplified from both embryonic and adult muscle tissues.
In addition, it has been suggested that, although the primary transcription of pri-miR-1/miR-133 occurs predominantly in muscle tissues, mature miR-1 and miR-133 are also differentially expressed throughout the body (Glass and Singla, 2011). High expression levels of miR-1/miR-133 have been found in cells positive for cardiac-specific markers such as the beta-myosin heavy chain in embryos; overexpression of miR-1 and miR-133 in ESCs can inhibit the expression of molecules involved in nonmuscle expression to promote differentiation into mesoderm, which gives rise to cardiac muscle, thus realizing directional regulation for the gradation of the germ layer.
However, as the cardiac muscle stem cells continue to differentiate, the miR-1 and miR-133 have opposite effects. miR-1 promotes further differentiation, but miR-133 inhibits the terminal differentiation of cardiac muscle. Consistent with this mechanism, transfection and overexpression of miR-1 were shown to promote the differentiation of ESCs into cardiac muscle and promote the repair of myocardial infarction-damaged tissue through regulation of the PTEN/AKT pathway (Glass and Singla, 2011). Muscle-specific miR-1 and miR-133 regulate ESC differentiation at the germ layer level and thus play an important role in further terminal differentiation of cardiac muscle.
miR-134
miR-134 is highly conserved and widely expressed in mice and other quadrupeds. Expression of miR-134 was shown to induce ESC differentiation in retinoic acid- and N2B27-induced cells (Tay et al., 2008). However, this function was not obtained when the experiment was performed using embryoids. In addition, as the expression of miR-134 increases, the expression of Sox1, Nestin, and Neurogenin-2, which are related to the differentiation of the ectoderm, also increases and this increase can be inhibited by an miR-134 antagonist.
In addition, miR-134 expression decreases the levels of positive regulatory factors Nanog, LRH1, and Oct4 in ESCs (Tay et al., 2008; Wu et al., 2007). Thus, miR-134 can promote the differentiation of ESCs into ectoderm by acting on Nanog and LRH1. Also, the miR-134 antagonist could inhibit endogenous miR-134 function directly and increase the number of ESC clones. Inhibition of miR-134 normally promotes ESC differentiation into the three germ layers, thus blocking full development in chimeric mice. Overexpression of miR-134 significantly represses the reprogramming progress. It was also confirmed that miR-134 can induce pre-ESC maturation and inhibition of miR-134 promotes pluripotency in pre-ESCs (Zhang et al., 2016).
miR-145
miR-145 has a very low expression in self-renewing human ESCs, but its expression increases rapidly during differentiation. It downregulates the totipotency factors Oct4, Sox2, and KLF4 to inhibit the self-renewal of human ESCs, and it induces the lineage-restricted differentiation of mesoderm and entoderm. Moreover, Oct4 inhibits the transcription of miR-145 by binding to its promoter, and it has been proposed that the reprogramming core factors and miR-145 regulate the self-renewal and differentiation of ESCs in a two-way negative feedback loop (Xu et al., 2009).
A recent study showed that p53 inhibits the expression of OCT4, SOX2, and KLF4 by activating miR-34a and miR-145, which promote the differentiation of ESCs (Jain et al., 2012). In addition, miR-145 participates in the differentiation of ESCs into endothelium and modulates ESC differentiation through the Oct4/Sox2 pathway (Xu et al., 2009; Yamaguchi et al., 2011). The unique identification of miR-145 in different organs and species indicates its critical function and evolutionary importance in cell physiology (Cruz-Santos et al., 2016; Wang et al., 2016). It has been reported that mir145 inhibits pluripotency and interacts with core pluripotency factors to control ESC differentiation (Adegani et al., 2013). In addition, miR-145 overexpression significantly reduces the expression of the self-renewal marker SSEA4 and upregulates multiple differentiation markers (Adegani et al., 2013; Latorre et al., 2016). However, downregulation of miRNAs has also been found to result in the upregulation of pluripotency-related genes (Adegani et al., 2011).
The let-7 cluster
The let-7 cluster is not expressed in ESCs, but its expression becomes apparent and increases gradually during ESC differentiations. The let-7 cluster has been found to regulate the differentiation of ESCs into neurons. In ESCs, Lin28 binds to pri-let-7 to inhibit its transformation into pre-let-7 and also inhibits the formation of the mature let-7 molecule, enabling the self-renewal of ESCs. After an induced differentiation, Lin28 is downregulated to relieve the inhibition of let-7 formation, which promotes the differentiation of ESCs into neurons (Viswanathan et al., 2008). Other studies on PGCs found that let-7 promotes differentiation by targeting Blimp1 (West et al., 2009).
Let-7 has been shown to downregulate Cdk4 and CyclinD to repress the G1-S transition (Hertel et al., 2012). Interestingly, let-7 also directly targets several ESC-enriched genes, such as Lin28, Sal4, C-myc, and N-myc. Mature let-7 miRNAs are potently induced during ESC differentiation to accelerate the exit from pluripotency (Choi et al., 2015). Surprisingly, even though the levels of mature let-7 miRNA are very low in ESCs, pri-let-7 miRNAs are expressed at an invariable level in self-renewing and differentiating ESCs because let-7 maturation is blocked in self-renewing ESCs. In addition, let-7 miRNAs repress important cell proliferation and pluripotency genes (Melton et al., 2010). Furthermore, Lin28 is a negative regulator of let-7, making it a potent reprogramming factor. In addition, let-7 miRNAs constitute a major barrier for maintaining a differentiated cell fate.
Core pluripotency factors
The molecular mechanisms underlying early embryonic development, especially the genes that control the differentiation of ESCs, have been receiving increasing attention. Core pluripotency factors, including Oct4, Sox2, and Nanog, are important genes in ESC differentiation and pluripotency. OCT4 prevents ESC differentiation and maintains pluripotency by forming repressor complex 9. SOX2 also controls downstream gene expression to maintain pluripotency and inhibit differentiation in ESC development. NANOG maintains ESC self-renewal by controlling the expression of gata4 and gata6 (Li et al., 2016). Some pluripotency-related miRNAs can manipulate the expression of core pluripotency factors. For example, miR-290 and miR-302 clusters have been shown to activate TCF3, NANOG, SOX2, and OCT4 promoters (Lee et al., 2013).
In addition, the miR-302 can bind to the promoters of OCT4/SOX2 and induce transcription by inhibiting NR2F2 translation (Hu et al., 2013). MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human ESCs (Xu et al., 2009). Both OCT4 and SOX2 bind to the promoter region of the miR-200 cluster and activate its transcription (Wang et al., 2013). miR-29b transcription is also induced early during reprogramming in ESCs by SOX2. Oct4 and Sox2 have been reported to regulate the expression of miR-302 in mice (Guo et al., 2013). These data suggest that the core pluripotency factors and miRNAs cooperate to preserve pluripotency and self-renewal during mouse embryonic development. However, the exact relationship between the core pluripotency factors and miRNAs is unclear, and more research is required in the future.
Other miRNAs
In addition to the above-described well-studied miRNAs, there are also reports on the influence of other miRNAs on ESC differentiation. For example, changes in the expression of miR-17-5p, miR-93, miR-20a, and miR-106a, which are members of the miR-17 cluster, during embryonic development, affect ESC differentiation. miR-93 induces the formation of primeval endoderm and trophectoderm; miR-17-5p and miR-93 are highly expressed in the mesoderm of gastrulae. In an ESC differentiation model, it was shown that the regulation of specific miRNAs in the miR-17 cluster delays or promotes differentiation. These miRNAs regulate the expression of STAT3, which is an important regulator of the differentiation and proliferation of ESCs (Foshay and Gallicano, 2009).
miR-25 and miR-10a are highly expressed during endodermal differentiation. The miR-122 and miR-192 levels are in line with the gene expression levels in the mesoderm; however, the stable and high expression of miR-122 in ESCs does not promote differentiation. Rather, it has an inhibitory effect (Tzur et al., 2008). In addition, miR-124 has been shown to promote the differentiation of ESCs into neurons (Kashyap et al., 2009); miR-203 promotes the differentiation of ESCs into epithelium (Lena et al., 2008); the miRNA complex consisting of miR-99b, miR-181a, and miR-181b promotes the differentiation of ESCs into vascular endothelial cells (Kane et al., 2012); the miR-200 cluster is involved in the differentiation of ESCs into mesoderm (Rosati et al., 2011); and miRs-126/126* regulates the differentiation of ESCs into hematopoietic cells (Huang et al., 2011). Importantly, some other miRNAs have been implicated in the differentiation of ESCs into somatic stem cells and their special functions (Table 1).
mESC, mouse embryonic stem cells; mHSC, human embryonic stem cells; miRNA, microRNA; PcG, polycomb group; TCR, T cell receptor.
Discussion
Although the importance of miRNAs in regulating the differentiation and self-renewal of ESCs was demonstrated many years ago, it has been challenging to elucidate the underlying mechanisms. miRNAs are considered to be involved in a powerful mechanism to regulate functional gene products. However, when using antagomirs or mimics to downregulate or upregulate miRNAs, the expressed protein of the target gene may differ among individual cells and experiments, thus complicating the explanation of the observed phenotypes.
In addition, obtaining a clear interpretation of the direct miRNA-regulated targets and pathways has been challenging. The miRNA-mediated differentiation and self-renewal of ESCs are inherently complex. The length of miRNAs necessary for partial target recognition is only 6–8 bases, and the target site that a miRNA can recognize varies among different cell types. The expression of miRNA-mediated genes appears to be controlled at the level of each individual target; however, many other factors in a single pathway can be influenced by a single miRNA at the same time, which results in a feedback effect on the functional response of the pathway itself. Therefore, elucidating the molecular mechanisms of miRNA enhancers or repressors important for ESC differentiation and self-renewal remains a major challenge.
Emerging evidence suggests that noncoding RNAs also play an important role in regulating ESC differentiation and renewal. A genome-wide process was used to identify hundreds of large intervening noncoding RNAs that were expressed in ESCs (Carey et al., 2011). Thus, these data could shed light on new novel mechanisms and research directions for ESC renewal and differentiation, and noncoding RNAs could be an exciting field for research on ESCs in the future.
Since miRNAs were discovered in the 1990s, they have been increasingly confirmed to play a critical role in stem cell self-renewal and differentiation (Fig. 1). Further studies are necessary to obtain a detailed understanding of the function of miRNAs in the biology of stem cell self-renewal and differentiation. It remains unclear whether different miRNAs interact with each other in regulating stem cell processes and if there is a molecular regulatory network that controls self-renewal and differentiation. Discovering novel pluripotency genes and pathways controlled by miRNAs is a critical future direction of research to overcome hurdles impeding the application of ESCs, such as the difficulty in differentiating ESCs to the desired cell types in large amounts. In addition, extensive basic research and preclinical investigations are required to achieve the clinical application of miRNAs. The resulting insights into the molecular mechanisms of stem cells will provide novel strategies for clinical research diagnostics and targeted therapeutics, laying a basis for translational research and clinical application.
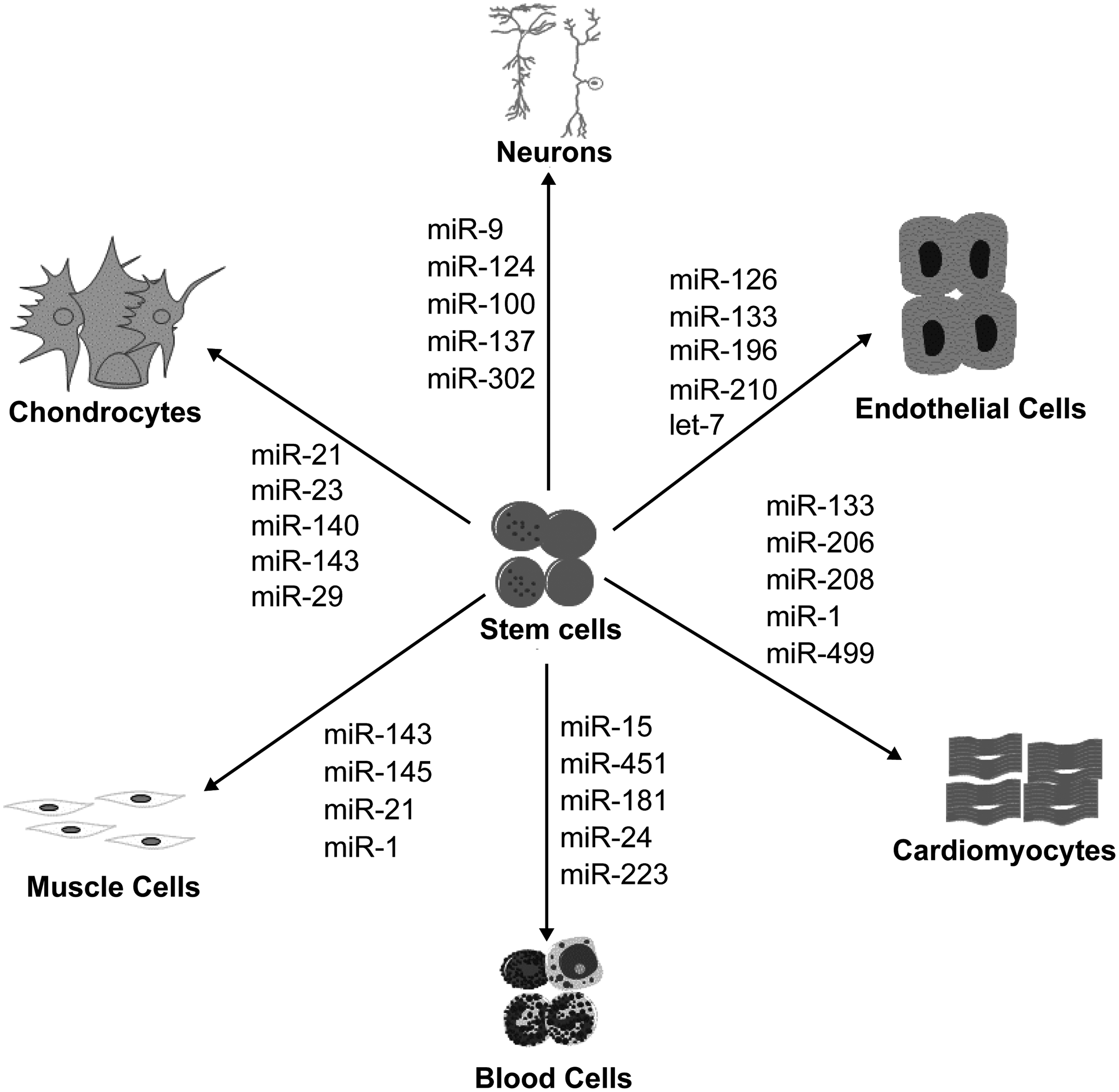
miRNAs regulate differentiation in various somatic stem cells. Shown are several lineage-specific cells that are regulated by special miRNAs. miRNAs, microRNAs.
Footnotes
Acknowledgment
This work was funded by the Special Fund for National Key Project “973” for Development of Basic Research (Grant No. 2012CB518103).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.