
Abstract
Abstract
In mammals, the imprinted genes play vital roles in development and are generally controlled by DNA methylation at imprinting control regions (ICRs). Recently, it was discovered that 5-hydroxymethylcytosine (5-hmC) is a stable epigenetic modification; however, its functions in cloned animal genomes have not yet been fully elucidated. In this study, we interrogated and quantified the 5-hmC levels in the brain of cloned goats and discovered upregulation of Uhrf1 (p < 0.001), Dnmt1 (p < 0.05), Dnmt3a (p < 0.05), Igf2 (p < 0.01), and H19 (p < 0.05) and downregulation of Dnmt3b (p < 0.001), Tet1 (p < 0.001), Tet2 (p < 0.05), Tet3 (p < 0.001), Mecp2 (p < 0.05), and Igf2r (p < 0.05) in deceased cloned goat tissues compared with the normal controls. We demonstrated that DNA methylation was increased at H19 ICR (51.33% ± 2.03% vs. 93.07% ± 3.06%; p < 0.01) and that DNA was hypomethylated at Igf2 ICR (4.57% ± 1.48% vs. 7.63% ± 1.83%; p > 0.05) in the brain of deceased cloned goats. Finally, we showed that within the cloned goat brain genome, the amount of genome-wide 5-hmC was significantly decreased (0.083% ± 0.026% vs. 0.024% ± 0.007%; p < 0.05), whereas the 5-hmC levels within H19 and Igf2 CCGG sites were not significantly altered (0.17% ± 0.09% vs. 0.03% ± 0.01%; p > 0.05) in the brain of deceased cloned goats. Our data bring further experimental evidence regarding the abnormalities in 5-hmC and advance our current understanding of the role of 5-hmC in cloned animals.
Introduction
G
Recently, it was shown that DNA methylation can be iteratively oxidized by the ten-eleven translocation (TET) enzymes to yield 5-hydroxymethylcytosine (5-hmC), 5-formylcytosine (5-fC), and 5-carboxycytosine (5-caC) (He, 2011; Ito, 2011; Tahiliani et al., 2009). The 5-hmC modification has been shown to be a stable epigenetic modification in its own right. It is associated with gene expression, aging, and postnatal neural development (Li and O'Neill, 2013; Pfeifer et al., 2013; Stroud et al., 2011; Szulwach et al., 2011).
Somatic cell nuclear transfer (SCNT) is an exciting experimental approach for elucidating the basic epigenetic mechanisms involved in disease and embryonic development (Jaenisch and Young, 2008). However, several pathologies associated with aberrant DNA methylation and genomic imprinting, including abnormal organ size, respiratory issues, and perinatal death, have been described in cloned conceptuses and neonates (Cibelli et al., 2002; Farin et al., 2006; Ogura et al., 2013; Yang et al., 2007). The loss of 5-hmC signal in bovine (Zhang et al., 2016) and decreased 5-hmC signal in swine (Cao et al., 2014) have been observed during SCNT embryo development. These studies investigated the abnormal dynamics of 5-hmC in SCNT-cloned embryos. As we hypothesized that the amount of 5-hmC was also abnormal in tissues of cloned animals and could be different across animals, it would be important to determine the 5-hmC levels in cloned goat tissues.
In this study, we interrogate and quantify 5-hmC levels in the brains of cloned goats, both globally and locally. Our data show that the amount of genome-wide 5-hmC was significantly decreased, whereas the 5-hmC levels at H19 and Igf2 CCGG sites were not significantly altered. Furthermore, we observed hypermethylation at H19 ICRs and hypomethylation at Igf2 ICRs. These findings demonstrate that the 5-hmC level was abnormal in the brain of cloned goats.
Materials and Methods
Cloned goat production and sample collection
The generation of SCNT-cloned goats was accomplished as previously described (Meng et al., 2013; Wan et al., 2012). Briefly, donor cells at passage 5 were thawed and subcultured in a 12-well culture plate at 37°C with 5% CO2 and saturated humidity. When the cells were 80% confluent, they were serum starved for 3 days by culturing in Dulbecco's modified Eagle's medium (Gibco) containing 0.5% fetal bovine serum (Gibco). Oocytes in MII stage were obtained by flushing a donor goat following superovulation. Oocytes were treated with 1.0 μg/mL demecolcine solution (Sigma) for 30 min, after which the first polar body, nucleus, and a small amount of cytoplasm were removed by aspiration.
Subsequently, the donor cell was inserted between the zona and cytoplast membrane followed by fusion with two 1.2 kV/cm DC pulses lasting 20 μs (BTX, Inc., San Diego, CA) in fusion solution. The reconstructed embryos were activated in 5 mM ionomycin for 5 min, followed by incubation in 2 mM 6-DMAP for 4 h, and culturing in M16 (Sigma) for 8 h. Finally, these one-cell stage embryos were surgically implanted (approximately eight embryos per goat) into the oviducts of local recipient goats at 12 to 24 h after estrus.
Due to extremely low efficiency of somatic reprogramming, our efforts only resulted in 12 cloned transgenic goats, and 3 female goats died at around 3 days after birth. Fetal weight, age at death, and cause of death were determined. Based on a prior report that 5-hmC is age dependent in the mammalian brain (Szulwach et al., 2011), three normal age-matched goats from conventional reproduction were slaughtered as controls. Samples from the cloned and control goats were collected immediately after euthanasia and tissues were excised and stored at −80°C in RNA/DNA protector (Takara, Dalian, China). The transgenic goats were verified by using polymerase chain reaction (PCR) screening and Southern blotting (data not shown).
All animal handling and experimentation were in accordance with the National Research Council's publication “Guide for the Care and Use of Laboratory Animals” and approved by Institutional Animal Care and Use Committees at Nanjing Agricultural University.
Gene expression analysis
Total RNA was extracted from tissues using an RNAprep Pure Tissue Kit (Tiangen, Beijing, China) following the manufacturer's instruction. The cDNA was synchronized using the PrimeScript™ RT reagent kit with gDNA Eraser (TaKaRa, Dalian, China) according to the manufacturer's protocol. Quantitative real-time PCR (qPCR) was performed as previously described (Wan et al., 2016). A complete list of primers used in this study is described in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/cell).
DNA methylation status of H19/Igf2 ICR
Genomic DNA was extracted from goat brain by using a TIANamp Genomic DNA Kit (Tiangen, Beijing, China) and eluted in 50 μL TE Buffer and stored at −20°C for further PCR and epigenetic analyses.
The DNA methylation status of the H19 and Igf2 ICRs was analyzed by bisulfite sequencing. Briefly, genomic DNA was treated with sodium bisulfate, using the EZ DNA Methylation-Direct Kit (ZYMO RESEARCH) and PCR amplified. The PCR procedures were performed as follows: one cycle of 95°C for 5 min; two cycles of 95°C for 30 s, 60°C (annealing Tm) for 1 min, and 72°C for 1 min; two cycles of 95°C for 30 s, 58°C for 1 min, and 72°C for 1 min and so forth; and 20 cycles of 95°C for 30 s, 50°C (annealing Tm) for 1 min, 72°C for 1 min, and a final cycle of 72°C for 7 min. The annealing Tm for H19 and Igf2 ranged from 60°C to 50°C and from 62°C to 54°C, respectively. The PCR products were then cloned into the pMD19-T vector (Takara, Dalian, China) and the clones were sequenced. The DNA methylation patterns derived from more than 10 clones with 99% cytosine conversions were analyzed using the BIQ Analyzer software. A complete list of primers used in bisulfite sequencing is described in Supplementary Table S2.
Quantification of 5-hmC at specific CpG dinucleotides
The quantification of 5-hmC at H19 and Igf2 CCGG sites was performed using the EpiMark 5-hmC and 5-mC Analysis Kit (E3317S; EpiMark, New England Biolabs) according to the manufacturer's protocol. Briefly, DNA was treated with T4 β-glucosyltransferase (T4-BGT), which can specifically glucosylate 5-hydroxymethylcytosine to yield β-glucosyl-5hmC (5ghmC). MspI is a restriction enzyme, capable of cleaving its recognition sequences containing either 5mC (C5mCGG) or 5hmC (C5hmCGG), but cleavage is blocked by 5ghmC (C5ghmCGG). The extent of DNA template cleavage by MspI before and after treatment with T4-BGT was determined by qPCR. This assay allows us to estimate the absolute 5-hmC levels at the MspI sites. A complete list of primers used in this experiment is shown in Supplementary Table S3.
Quantification of genome-wide 5-hmC
Quantification of genome-wide 5-hmC was accomplished by using a colorimetric Methyl Flash Hydroxymethylated DNA Quantification Kit (Epigentek, Farmingdale, NY) according to the manufacturer's protocol. Briefly, 200 ng DNA was added to the assay well and incubated with anti-5-hmC antibody (50 μL, 1000 μg/mL) at room temperature (RT) for 60 min and then incubated with a second antibody (50 μL, 400 μg/mL) at RT for 30 min. Enhancer and developer solution were added and the signal was detected using a microplate reader at 450 nm. Negative and positive controls were also performed in triplicate.
Statistical analysis
All qPCRs were performed in triplicate. Data are presented as mean ± standard error of the mean (SEM). DNA methylation status, amount of 5-hmC, and gene expression between the deceased cloned goats and controls were tested by Student's t-test using SPSS 18.0 software (SPSS, Inc., Chicago, IL). A difference with p-value of <0.05 was considered statistically significant.
Results
Morphology of transgenic cloned goats
Fetal weight at necropsy was greater in the deceased cloned goat group (3.43 ± 0.87 kg vs. 1.47 ± 0.15 kg; p < 0.05), which was consistent with large offspring syndrome (LOS). All three deceased transgenic cloned goats were female and derived from the same donor cell line, and one of the deceased transgenic clones exhibited intestinal disease (Table 1).
Column data marked with different superscripts denotes significant differences of p < 0.01 (Student's t-test). Values are presented as the mean ± SD.
—, parameterless property; ND, not determined; S, slaughtered; P, after birth.
Gene expression characteristics in cloned goat tissues
Before we analyzed the expression of key DNA methylation genes in cloned goat tissues, we investigated the integrity of the RNAs. Our data revealed that total RNA from the brain of the dead cloned goats and slaughtered control goats are integrated and not degraded (Supplementary Fig. S1). As shown in Figure 1A, the expression of Uhrf1 (p < 0.001), Dnmt1 (p < 0.05), and Dnmt3a (p < 0.05) was increased, whereas the expression of Mecp2 (p < 0.05) and three DNA demethylases Tet1 (p < 0.001), Tet2 (p < 0.05), and Tet3 (p < 0.001) was decreased in the deceased cloned goat brain, compared to the controls. In the liver tissue form deceased cloned goats, the expression of Dnmt3b was significantly decreased (p < 0.001), whereas the expression of Uhrf1 (p < 0.01) was significantly increased (Fig. 1B). The expression of Dnmt3b (p < 0.001) and Tet3 (p < 0.05) was decreased, whereas the expression of Dnmt3a (p < 0.05) was increased in the lungs of deceased cloned goats (Fig. 1C).
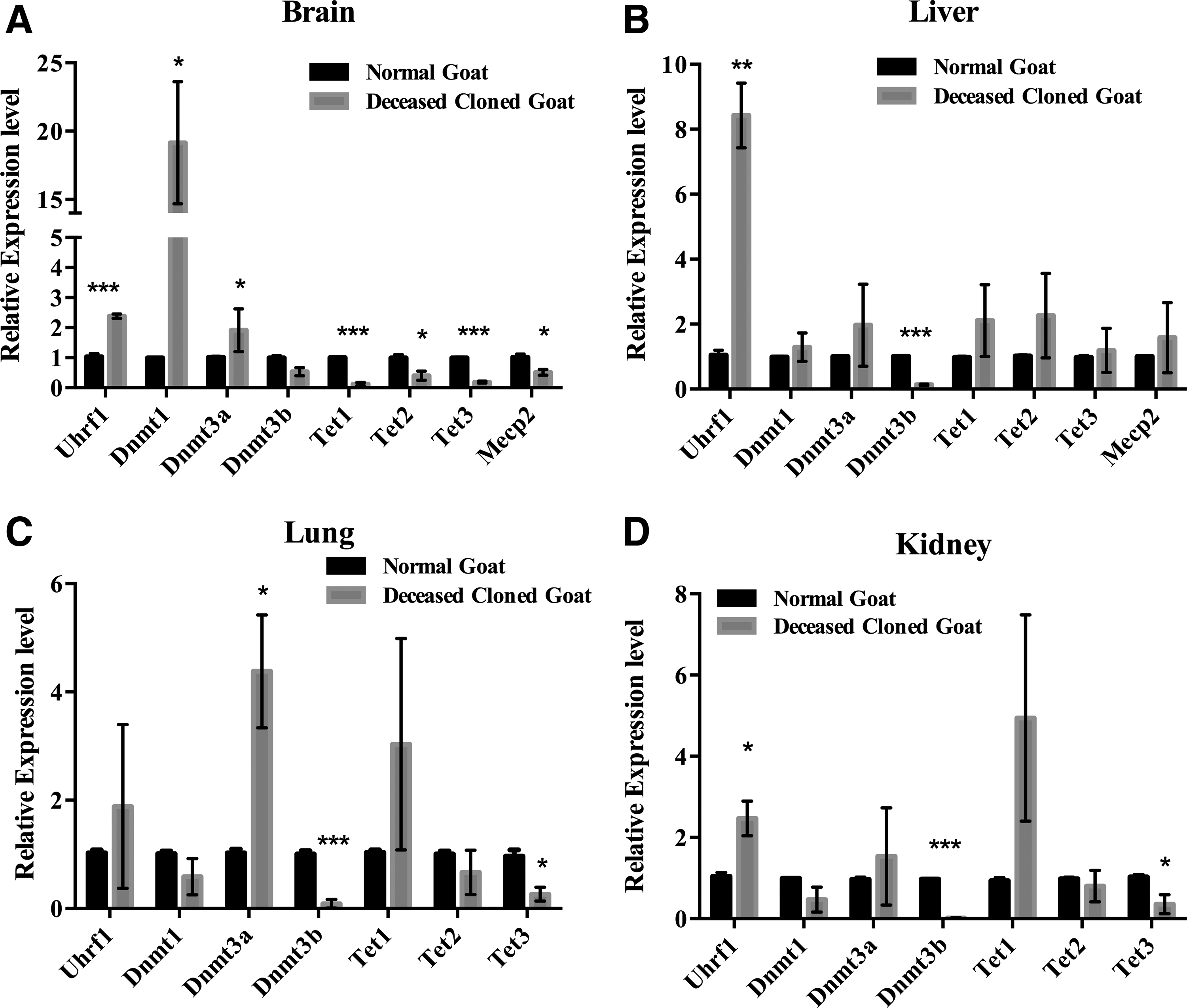
Quantitative analysis of DNA methyltransferase (Dnmts), ten-eleven translocation (Tets), Uhrf1, and Mecp2 expression in
Furthermore, the expression of Uhrf1 (p < 0.05), Dnmt3b (p < 0.001), and Tet3 (p < 0.05) was reduced in the kidneys of deceased cloned goats compared with the control group (Fig. 1D). Notably, the expression of Uhrf1 in the brain, liver, and lung tissue, the expression of Dnmt1 in the brain, and the expression of Tet1 in the lung and kidney were highly variable between the individual clones and even in organs from the same clone. These results indicate that DNA methylation might be irregular in cloned goat tissues, particularly in brain tissue.
We next studied the expression of four imprinted genes in the brain and liver tissues from the deceased cloned goats. As shown in Figure 2A, the expression level of Igf2r (p < 0.05) and Xist (p < 0.01) was downregulated, whereas the expression of H19 (p < 0.05) and Igf2 (p < 0.01) was significantly upregulated in deceased cloned goat brain tissue compared with the expression in brain tissue from the control group. In the livers of deceased cloned goats, the expression of Igf2r (p < 0.05) was decreased, whereas the expression of Xist (p < 0.05), H19 (p < 0.01), and Igf2 (p < 0.01) was markedly increased (Fig. 2B).

Quantitative analysis of imprinted gene Igf2r, Xist, H19, and Igf2 expression in deceased cloned goat
Taken together, these results suggest that the DNA methylation of these three imprinted genes at their ICRs might be modulated.
Methylation status of H19/Igf2 ICR
We selected the H19 and Igf2 loci as a representation of imprinted genes and investigated the methylation status of the putative H19 and Igf2 ICRs by using bisulfite sequencing. The H19 putative ICR was located upstream of the promoter, from locus 1362 to 1683. It contained 362 nucleotides and 1 CTCF motif, and covered 25 CpG sites (Fig. 3A). This ICR was highly methylated in brains from deceased cloned goats (93.07% ± 3.06%; p < 0.01; Fig. 3C, D), but was only partially methylated in the control group (51.33% ± 2.03%; Fig. 3B).
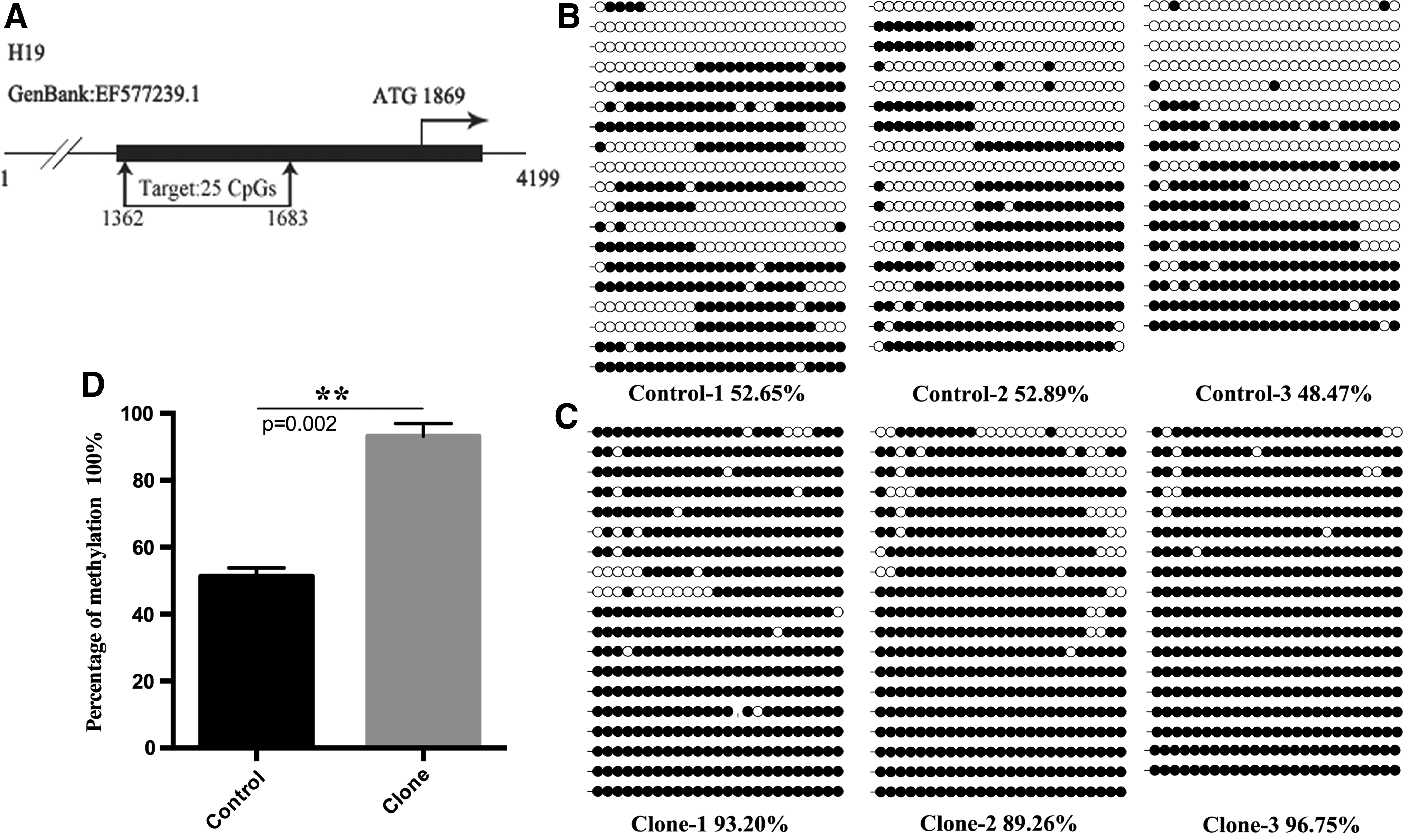
Methylation profiles of H19 ICR in deceased cloned goat brain. Methylation profile of H19 ICR was determined using bisulfite sequencing PCR. Horizontal line of circles represents one single clone that was sequenced. Each circle represents one of the CpG dinucleotides within the H19 ICR. Unfilled (white) and filled (black) circles represent unmethylated and methylated CpGs, respectively. Lollipop diagrams were generated by BIQ Analyzer software. Data were analyzed by Student's t-test and are shown as mean ± SEM. The symbol (**) denotes very significant differences of p < 0.01 between naturally reproduced goats and deceased cloned goats. ICR, imprinting control regions.
Next, we addressed the DNA methylation profile of the Igf2 locus. The Igf2 ICR was located in the promoter region. It had a higher CpG density, spanned one CpG island, and covered 32 CpG sites within 329 nucleotides (Fig. 4A). The percentage of DNA methylation at this ICR was 7.63% ± 1.83% in deceased cloned goat brains and 4.57% ± 1.48% in the control brains (Fig. 4B, C). However, no statistically significant difference in methylation was detected between these two groups (p > 0.05; Fig. 4D).
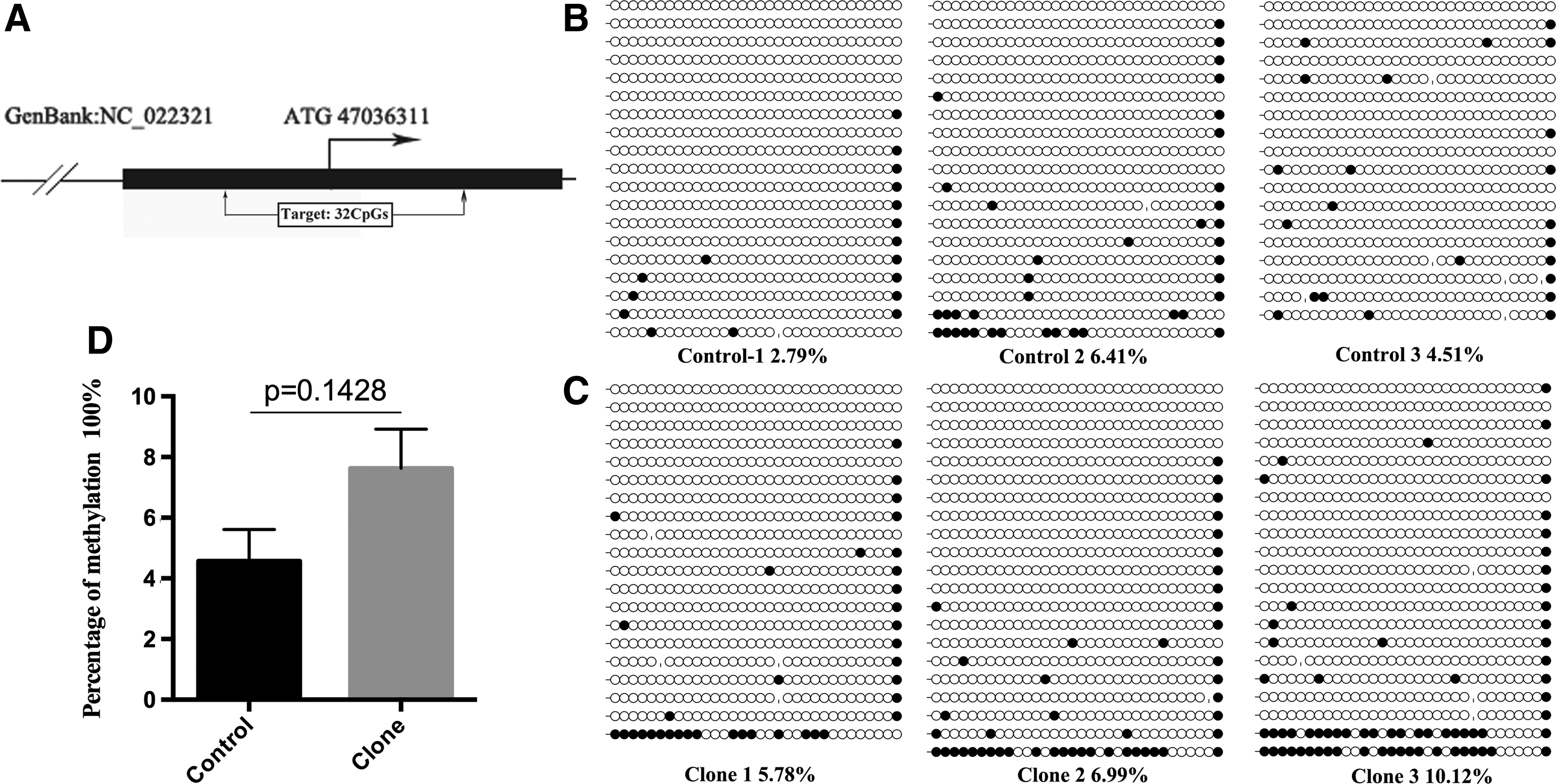
Methylation profiles of Igf2 ICR in deceased cloned goat brain. Methylation profile of Igf2 ICR was determined using bisulfite sequencing PCR. Horizontal line of circles represents one single clone that was sequenced. Each circle represents one of the CpG dinucleotides within the Igf2 ICR. Unfilled (white) and filled (black) circles represent unmethylated and methylated CpGs, respectively. Lollipop diagrams were generated by BIQ Analyzer software. Data were analyzed by Student's t-test and are shown as mean ± SEM. There was no significant difference between naturally reproduced goats and deceased cloned goats.
HmC in H19 and Igf2 CCGG sites
Because 5-hmC cannot be distinguished from 5-mC by bisulfite sequencing, we used an enzymatic assay to ensure that we correctly interpreted the bisulfite signal as 5-mC. Two H19 CCGG sites, located at the 19th and 24th CpG sites, were targeted within the putative ICR. The amount of 5-hmC presented at these two sites was not statistically significantly different in the cloned group compared with the control group (0.17% ± 0.09% vs. 0.03% ± 0.01%; p > 0.05; Fig. 5A, left panel). Taken the DNA methylation status at the H19 ICR into consideration, this result indicates that the entire bisulfite signal represented 5-mC.
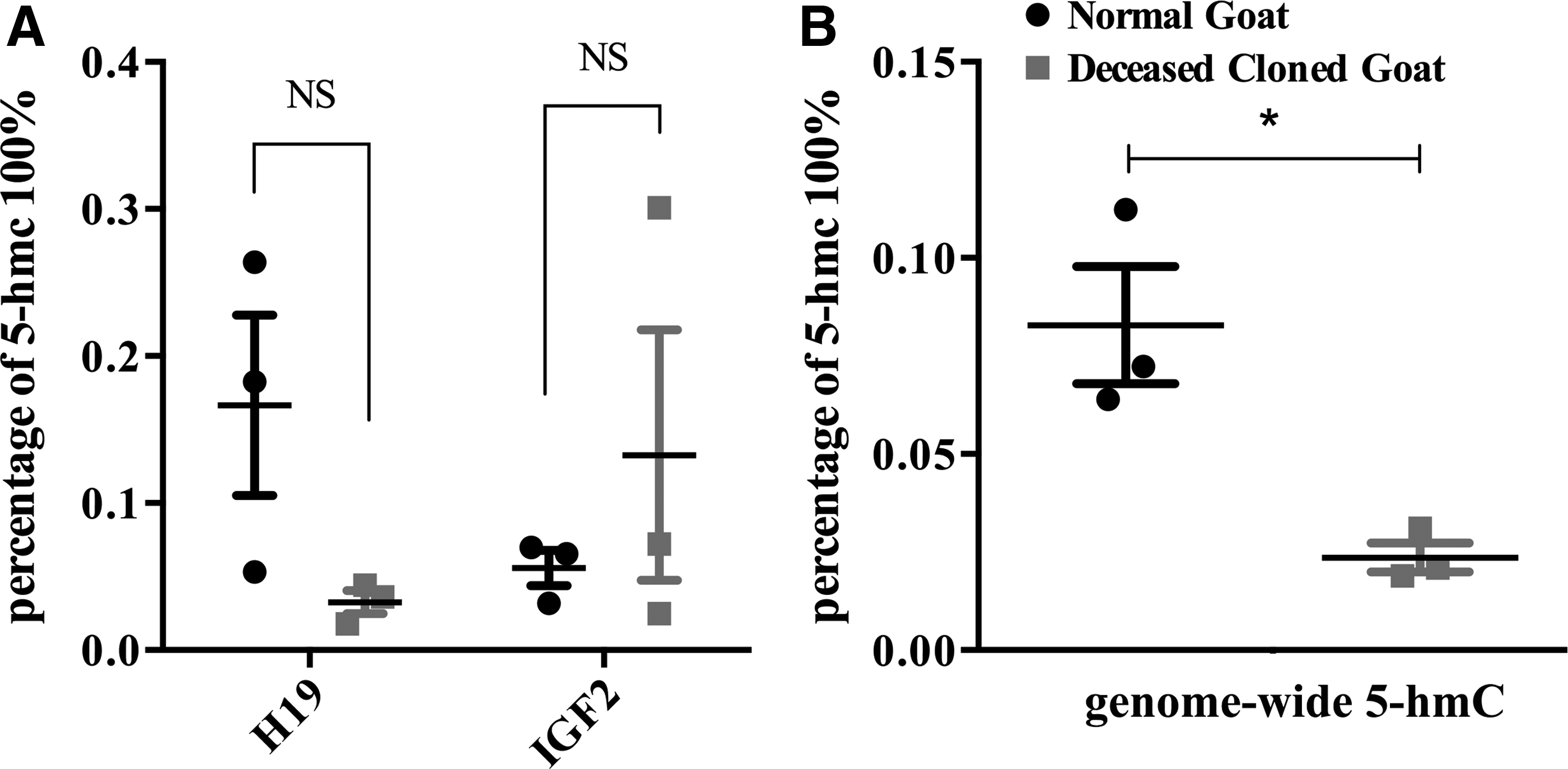
Loci-specific and genome-wide hydroxymethylcytosine profiles in the brain of deceased cloned goats. Genome-wide and locus-specific 5-hmC profile was determined by using ELISA and enzymatic reaction utilizing T4 β-glucosyltransferase.
For Igf2, we designed primers to target three CCGG sites in its promoter. In deceased cloned goat brains, the 5-hmC levels were slightly higher, but not significantly different, compared with the control group (0.06% ± 0.02% vs. 0.13% ± 0.12%; p > 0.05; Fig. 5A, right panel), which indicates that the 5-hmC levels were extremely low at the Igf2 promoter.
Quantification of HmC in goat brain
Our analyses of 5-hmC at H19 and Igf2 CCGG sites revealed no significant difference when we compared the deceased cloned goat group to the natural reproduction goat group. However, the characteristics of the global 5-hmC remained unclear. We quantified global expression of 5-hmC in goat brains and found that the amount of genome-wide 5-hmC was distinctly lower in brain tissues of deceased cloned goats compared with expression in that of control goats (0.083% ± 0.026% vs. 0.024% ± 0.007%; p < 0.05; Fig. 5B).
Discussion
The work presented in this study reveals that the expression of key DNA methylation genes was highly variable between individual cloned goats, even in organs from the same cloned goat, which was consistent with previous reports (Jiang et al., 2007; Meng et al., 2014; Yang et al., 2005). Specifically, our data revealed the upregulation of Uhrf1, Dnmt1, and Dnmt3a and the downregulation of Tet1, Tet2, Tet3, and Mecp2 in the brains of deceased cloned goats. These genes are associated with the DNA methylation process. DNA methylation is established and maintained by DNMTs and catalyzed by TETs to demethylation (Smith and Meissner, 2013). Ubiquitin-like, containing PHD and RING finger domains, 1, encoded by UHRF1, also plays a role in maintaining DNA methylation in mammalian cells by stimulating the activity and specificity of DNMT1 (Bashtrykov et al., 2014; Pacaud et al., 2014).
Methyl CpG binding protein 2, encoded by MeCP2, reads epigenetic information contained in the methylated and hydroxymethylated DNA and represses transcription by binding DNA (Della Ragione et al., 2016). Upregulation of these Key DNA methylation genes and downregulation of these three Tet genes could result in hypermethylation at repetitive sequences, specific nonimprinted loci, or imprinted ICR. To this end, we detected hypermethylation at the H19 ICR in brain tissues from deceased cloned goats.
H19 and Igf2 are two frequently studied imprinted genes that regulate early fetal growth. The expression of H19 is relatively low, whereas Igf2 is overexpressed in wild-type mice (Barlow, 2011), and a similar expression pattern was found in SCNT-cloned mice (Humpherys et al., 2001). In contrast, Ogawa et al. showed that the reduced expression of H19 was not associated with an increased Igf2 expression when hypermethylation of the H19 ICR was detected in cloned mouse (Ogawa et al., 2003). In this study, we found an increased expression of both H19 and Igf2, which is consistent with previous reports in cloned bovine embryos (Han et al., 2003) and placentas (Su et al., 2011). The increased expression of Igf2 could be related to LOS, because Igf2 is a growth-promoting hormone.
However, in our previous study, the expression level of H19 was significantly reduced in the lung of these deceased cloned goats (Meng et al., 2014). These apparent discrepancies could reflect the fact that the expression of H19 is highly variable between individual clones and even in organs from the same clone (Su et al., 2011; Wei et al., 2010).
Igf2 ICR in the brains of deceased cloned goats and those resulting from natural reproduction goats was highly hypomethylated, consistent with Ye et al. (2014). Thus, the increased expression of Igf2 in the brains of deceased cloned goats cannot be simply explained by the hypomethylation of Igf2 promoter. It was shown that Igf2 is clustered with H19, with both genes regulated by the ICR located between the two genes in mammals (Bartolomei and Ferguson-Smith, 2011; Thorvaldsen et al., 1998). This ICR contains the binding sites for the CTCF, which binds to the unmethylated allele and assembles a chromatin insulator that blocks the interaction between the Igf2 promoter and a downstream enhancer (Phillips and Corces, 2009).
Thus, we tested whether, in the brain of deceased cloned goats, the methylation level of H19/Igf2 ICR around one of the CTCF binding sites is different compared with the controls. We observed hypermethylation of H19 ICR in the brain of deceased cloned goats. These highly methylated ICRs could prevent CTCF binding, which facilitates Igf2 expression. An increased Igf2 expression might therefore result from hypermethylation of the CpGs around the CTCF binding sites.
The H19 ICR is only partially methylated in animals resulting from natural reproduction (Su et al., 2011; Zhao et al., 2013), which is in consist with our results. In the deceased cloned goats, the H19 ICR was highly methylated, which has been observed previously (Meng et al., 2013; Su et al., 2011; Wei et al., 2010). Thus, these prior studies and our results support this hypothesis that DNA demethylation was not completed during the reprogramming and could lead to a hypermethylation in deceased cloned offspring. Generally, the gain of DNA methylation at transcriptionally regulatory regions leads to a closed chromatin that is blocking accessibility for transcription factors, which results in the downregulation of gene expression. However, in our study, we discovered that the expression of H19 was increased and the H19 ICR was highly methylated. This result suggests that gene expression was epigenetically controlled at multiple levels (Zhou et al., 2015).
Our study demonstrated that the levels of genome-wide 5-hmC varied in the genomes of brain from deceased cloned goats and those resulting from natural reproduction. Reducing the levels of 5-hmC inhibits neuronal differentiation by regulating the expression of genes that are critical for this important developmental transition (Vasanthakumar and Godley, 2015). It is tempting to hypothesize that the global loss of 5-hmC and aberrant expression of Tet in the deceased cloned goats might lead to brain disorders and, ultimately, death, as reduced levels of 5-hmC might alter the ability to recruit Mecp2 (Mellen et al., 2012; Wang et al., 2013).
Although the 5-hmC exhibits tissue-specific distribution (Kinney et al., 2011; Nestor et al., 2011), we found reduced 5-hmC levels and decreased Tets expression in the brain of deceased cloned goats, which might have resulted from iterative oxidization of 5-hmC by TETs to yield 5fC and 5caC (Ito, 2011). It seems plausible that the activity of the TET enzymes could be another contributing factor, because the presence of vitamin C can enhance the TETs activity, even without change in the expression level (Minor et al., 2013). Furthermore, the amount of 5-hmC observed in the goat brain was ∼0.10%, which is lower than levels observed in the human brain. It is conceivable that this discrepancy resulted from the brains obtained from animals of different ages, as the expression of 5-hmC is age dependent and proliferating cells lose 5-hmC (Jin et al., 2011; Laird et al., 2013; Szulwach et al., 2011).
Finally, fibroblast cell line steadily expressing human lactoferrin (hLF) was used as donor cells for SCNT-cloned goat production in our study, because hLF plays key roles in antibacterial activity, antiviral activity, antifungal activity, and anticarcinogenic activity. We intended to produce large quantities of milk containing hLF. The green fluorescent protein (GFP) expression vectors or SV2-neomycin resistance genes were also presented in our hLF gene mammary expression vector. Published studies have demonstrated that transfection of bovine fetal fibroblasts with GFP expression vectors or SV2-neomycin resistance genes did not decrease their potency as nuclear donors (Zakhartchenko et al., 2001).
As for the extended culture associated with transfection and selection procedures, they might induce changes of donor cells, which markedly decrease the efficiency of nuclear transfer. Our previous data revealed that hLF transgenic cells showed poor proliferation (Wan et al., 2016). However, the methylation status of H19 ICR in the hLF transgenic fibroblast cells was similar with that in normal goat fibroblast cells (64.4% and 68.8%, respectively; unpublished data). Thus, this hLF-gene mammary expression vector does not influence the methylation status of H19 ICR.
In conclusion, our data demonstrate that the global, but not the locus specific, 5-hmC profile was abnormal in the brain of deceased cloned goats. We also observed modulation in the expression of key DNA methylation genes and abnormal DNA methylation at H19 ICRs in the brain of deceased cloned goat. Our results suggest that the methylation status of imprinted genes and the process of DNA demethylation in deceased cloned goats differ from the corresponding methylation status in brain tissue from goats obtained by natural reproduction. We concluded that these alterations could result in developmental insufficiencies and ultimately fetal or perinatal death in cloned goats. Our results also provide further experimental evidence for the abnormal expression of the 5-hmC and expand our current knowledge of the role of 5-hmC in cloned animals.
Footnotes
Acknowledgments
We are grateful to Ruoxin Jia for producing the SCNT-cloned embryos, and to Wenjun Zhou, Kaiping Deng, Zhen Wang, and Jingang Wang for performing the embryo transfer and raising the goats. This study was financially supported by the National Natural Science Foundation of China (No. 31301973; No.31272443) and by the National Major Special Projects on the Fundamental Research Funds for the Central Universities (No. KJQN201401).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.