
Abstract
Abstract
The application of cloning technology on a large scale is limited by very low offspring rate primarily due to aberrant or incomplete epigenetic reprogramming. Trichostatin A (TSA), a histone deacetylase inhibitor, and 5-aza-2′-deoxycytidine (5-aza-dC), an inhibitor of DNA methyltransferases, are widely used for altering the epigenetic status of cloned embryos. We optimized the doses of these epigenetic modifiers for production of buffalo embryos by handmade cloning and examined whether combined treatment with these epigenetic modifiers offered any advantage over treatment with the individual epigenetic modifier. Irrespective of whether donor cells or reconstructed embryos or both were treated with 50 nM TSA +7.5 nM 5-aza-dC, (1) the blastocyst rate was significantly higher (71.6 ± 3.5, 68.3 ± 2.6, and 71.8 ± 2.4, respectively, vs. 43.1 ± 3.4 for controls, p < 0.05); (2) the apoptotic index was lower (5.4 ± 1.1, 9.5 ± 1.0, and 7.4 ± 1.3, respectively, vs. 19.5 ± 2.1 for controls, p < 0.05) and was similar to that of in vitro fertilization blastocysts (6.0 ± 0.8); (3) the global level of H3K18ac was higher (p < 0.01) and that of H3K27me3 lower (p < 0.05) than in controls and was similar among all treatment groups; and (4) the expression level of epigenetic-(HDAC1, DNMT1, and DNMT3a), pluripotency-(OCT4 and NANOG), and development-related (FGF4) genes, but not that of SOX2 and CDX2, was similar among all treatment groups. These results demonstrate that similar levels of beneficial effects can be obtained following treatment of either donor cells or reconstructed embryos or both with the combination of TSA +5-aza-dC. Therefore, there is no advantage in treating both donor cells and reconstructed embryos when the combination of TSA and 5-aza-dC is used.
Introduction
C
Specifically, in case of bovines, only 9% of transferred embryos resulted in calves (Panarace et al., 2007), whereas across different species, it is only 0.9% to 6% (Loi et al., 2011). Majority of cloned embryos die during pre- and postimplantation development, and those which develop to term have been reported to show a high incidence of abnormalities such as large offspring syndrome, short life span, severe placental deficiency, respiratory problems, prolonged gestation, and dystocia. (Farin et al., 2006; Yang et al., 2007). The reasons behind all these factors, which contribute to lowering the cloning efficiency, are not fully understood. However, aberrant or incomplete nuclear reprogramming of the donor somatic cell by the oocyte is believed to be the primary reason responsible for these abnormalities, resulting in low cloning efficiency.
Several epigenetic modifiers have been used in many species for correcting epigenetic remodeling during reprogramming with the aim of improving the developmental competence and altering the epigenetic status of cloned embryos. Among these, trichostatin A (TSA), which is a potent histone deacetylase inhibitor, increases the pool of acetylated histones, induces DNA demethylation (Hattori et al., 2004; Yoshida et al., 1990), and can reduce abnormal DNA hypermethylation (Kishigami et al., 2006). TSA has been shown to increase the blastocyst rate and improve their quality by inducing hyperacetylation in NT bovine embryos (Cui et al., 2011; Ding et al., 2008).
Another strategy has been to inhibit the activity of DNA methyltransferases (DNMTs) through the use of inhibitors such as 5-aza-2′-deoxycytidine (5-aza-dC), which has been shown to improve the epigenetic status of cloned bovine embryos by decreasing their DNA methylation (Ding et al., 2008; Enright et al., 2005; Tsuji et al., 2009). In some studies, the donor cells (Enright et al., 2003) or the reconstructed embryos (Cui et al., 2011; Iager et al., 2008) were exposed to TSA, whereas in others, the donor cells were treated with 5-aza-dC (Enright et al., 2003, 2005). We (Saini et al., 2016) and others (Luo et al., 2013) have earlier shown that treatment of donor cells with TSA and 5-aza-dC in combination improved the developmental competence and quality of cloned buffalo embryos.
Although there are a few studies in cattle, in which both donor cells and reconstructed embryos were exposed to both of these epigenetic modifiers (Ding et al., 2008; Wang et al., 2011), no such study has been conducted in buffalo, an important source of milk, meat, and draught power in developing nations of Asia. The aim of this study was to find out if treatment of both donor cells and reconstructed embryos with TSA and 5-aza-dC in combination offered any advantage over treatment of either donor cells or reconstructed embryos with either of the two epigenetic modifiers individually. We examine the effect of treatment with these epigenetic modifiers on the developmental competence, quality, epigenetic status, and gene expression in buffalo embryos produced by handmade cloning (HMC).
Materials and Methods
All the chemicals were obtained from Sigma Chemical Co. (St. Louis, MO); the media were from GIBCO (Grand Island, NY), and the disposable plastic ware was purchased from Nunc (Roskilde, Denmark) unless otherwise stated. In vitro culture of somatic cells, oocytes, and embryos was carried out in a CO2 incubator (5% CO2 in air) at 38.5°C.
Production of cloned and IVF embryos
Somatic cells obtained from ear skin biopsy taken from an adult 8-year-old female buffalo (Mu-5517) and established in culture were used as donor cells for HMC for producing blastocyst-stage embryos. HMC, which included in vitro maturation (IVM) of cumulus–oocyte complexes (COCs) obtained from slaughterhouse buffalo ovaries, zona removal, manual enucleation, fusion, activation, and in vitro culture (IVC) of cloned embryos, was performed as described previously (Selokar et al., 2012). For production of IVF embryos, COCs were aspirated from abattoir buffalo ovaries following which IVM and IVF were carried out as described previously (Sharma et al., 2011). Day 8 blastocysts produced by HMC or IVF were stored at −80°C until further use for gene expression analysis.
Assessment of embryo quality
For assessment of the quality of embryos, total cell number (TCN) and the level of apoptosis in day 8 blastocysts were determined by terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining using the In Situ Cell Death Detection Kit, Fluorescein (11684795910; Roche), as described previously (Selokar et al., 2014).
TSA and 5-aza-dC treatment
For the treatment of donor cells with TSA and/or 5-aza-dC, these were seeded at a concentration of 1 × 104 cells/well in six-well culture plates and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 0.68 mM
Immunofluorescence staining for epigenetic markers in cloned embryos
The global level of H3K18ac and H3K27me3 was examined by immunofluorescence staining in cloned blastocysts as described previously (Saini et al., 2016). NIS-element basic research image processing software (Nikon, Tokyo, Japan) equipped with a microscope was used for image acquisition and quantitative measurements of the mean pixel intensity emitted by each individual nucleus. A minimum of 15 blastocysts (50 nuclei/blastocyst) were analyzed for each epigenetic marker.
Gene expression analysis in embryos
Gene expression analysis was carried out by quantitative real-time polymerase chain reaction (qPCR) as described previously (Saini et al., 2016). Briefly, RNA was isolated from blastocysts (n = 3–4) using the RNAqueous micro kit (Ambion, Austin, TX) according to the manufacturer's instructions. The genomic DNA contamination was removed by DNase treatment at 37°C for 20 minutes. The reverse transcriptase (RT) reaction was achieved using the Moloney murine leukemia virus (M-MLV) RT provided in the SuperScript reverse transcriptase III kit (Invitrogen). qPCR was performed using the optimized primer sets shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/cell) on CFX96 real-time system (Bio-Rad) with maxima@SYBR Green master mix (Fermentas, St. Leon-Rot, Germany) at the following thermal cycling conditions: 95°C for 5 minutes, followed by 40 PCR cycles at 95°C for 15 seconds, 58°C for 30 seconds, and 72°C for 30 seconds.
Melting peaks were determined using melting curve analysis to ensure specific amplification. Agarose (2%) gel electrophoresis analysis was carried out to determine the length of the amplified PCR products. β-Actin mRNA was employed as an internal standard for the analysis of relative transcript levels of each gene. cDNA was replaced by H2O in the negative controls. For comparison, the average expression level of each gene from IVF embryos was set as 1. Three separate experiments were performed with three replicates for each gene.
Experimental design and statistical analysis
First of all, we examined the effect of treating HMC embryos with epigenetic modifiers on developmental competence and embryo quality. Reconstructed embryos were cultured with TSA (0, 25, 50, 75, or 100 nM) or 5-aza-dC (0, 5, 7.5, or 10 nM) for 10 hours. Developmental competence was assessed by the cleavage and blastocyst rate at day 8 of culture, and the apoptotic level in produced blastocysts was examined by determining the TCN and the apoptotic index by TUNEL assay. Then, based on the results of this experiment, we treated both the donor cells and the reconstructed embryos with either TSA (0, 50, or 75 nM) or 5-aza-dC (0, 7.5, or 10 nM) and examined the effects on the developmental competence and quality of cloned embryos.
Following this, based on the results of previous experiments and our previous study (Saini et al., 2016), we treated the donor cells, the reconstructed embryos, and both with 50 nM TSA +7.5 nM 5-aza-dC and examined the effects on the developmental competence and quality of cloned embryos. The apoptotic index was compared with that of IVF embryos. We also investigated the effects of these treatments on the global level of H3K18ac and H3K27me3 in cloned blastocysts. Finally, we examined the effects of treatment of donor cells, reconstructed embryos, or both with 50 nM TSA +7.5 nM 5-aza-dC on the expression level of important pluripotency-(OCT4, NANOG, and SOX2), trophectoderm-(CDX2), epigenetic-(DNMT1, DNMT3a, and HDAC1), and development-related (FGF4) genes.
The percentage data were analyzed using SYSTAT 12.0 (SPSS, Inc., Chicago, IL) after arcsine transformation. Differences between means were analyzed by one-way analysis of variance, followed by Fisher's least significant difference (LSD) test. Differences were considered to be significant at p < 0.05.
Results
Effect of treatment of cloned embryos with TSA or 5-aza-dC
Following treatment of reconstructed embryos with TSA (0, 25, 50, 75, or 100 nM), the cleavage and blastocyst rate and TCN were not significantly different among different groups (Table 1). However, the apoptotic index of blastocysts was significantly lower (p < 0.05) for the 25, 50, and 75 nM TSA groups than that for the 100 nM TSA group or for that of controls. Following treatment of reconstructed embryos with 5-aza-dC (0, 5, 7.5, or 10 nM), the blastocyst rate of 10 nM 5-aza-dC group was higher (p < 0.05) than that of controls, whereas that of 5 and 7.5 nM 5-aza-dC groups was not significantly different from these two groups. The apoptotic index of 7.5 and 10 nM 5-aza-dC groups was lower (p < 0.05) than that of other groups, whereas the cleavage rate and TCN were similar among all the groups.
Data from 10 trials; values with different superscripts within the same column for TSA or 5-aza-dC differ significantly (p < 0.05).
5-aza-dC, 5-aza-2′-deoxycytidine; TSA, trichostatin A.
Effect of treatment of donor cells and cloned embryos with TSA or 5-aza-dC
Based on the results of previous experiments, we then treated both the donor cells and the reconstructed embryos with either TSA (0, 50, or 75 nM) or 5-aza-dC (0, 7.5, or 10 nM). Following treatment with TSA, the cleavage and blastocyst rate and TCN were not significantly different among different groups; however, the apoptotic index was lower (p < 0.05) in all the TSA treatment groups than in the controls (Table 2). The blastocyst rate was higher (p < 0.05) and the apoptotic index was lower (p < 0.05) for all the 5-aza-dC treatment groups than in the controls, although the cleavage rate and TCN were similar.
Data from 8 trials; values with different superscripts within the same column for TSA or 5-aza-dC differ significantly (p < 0.05).
Effect of treatment of donor cells and cloned embryos with TSA +5-aza-dC
In this experiment, we treated either the donor cells or the reconstructed embryos or both with 50 nM TSA +7.5 nM 5-aza-dC. The blastocyst rate was higher (p < 0.05) and the apoptotic index was lower (p < 0.05) than that of the controls, irrespective of whether the donor cells or the reconstructed embryos or both were treated with TSA +5-aza-dC (Table 3), although the cleavage rate and TCN were similar among different groups. The apoptotic index was similar to that of IVF embryos when either the donor cells or both donor cells and reconstructed embryos had been treated with TSA +5-aza-dC, but was higher (p < 0.05) when only reconstructed embryos had been treated or when no treatment was given (controls).
Data from 12 trials; values with different superscripts within the same column for TSA or 5-aza-dC differ significantly (p < 0.05).
HMC, handmade cloning.
Effect of treatment of both donor cells and cloned embryos with TSA +5-aza-dC on the epigenetic status
Based on the results of the previous experiment, we then treated either the donor cells or the reconstructed embryos or both with the combination of the two epigenetic modifiers, that is, 50 nM TSA +7.5 nM 5-aza-dC. The global level of H3K18ac was found to be significantly higher (p < 0.01) and that of H3K27me3 lower (p < 0.05) compared with that in the controls, irrespective of whether the donor cells or reconstructed embryos or both were treated with 50 nM TSA +7.5 nM 5-aza-dC (Fig. 1). The global level of each of these epigenetic markers was similar among the three treatment groups.
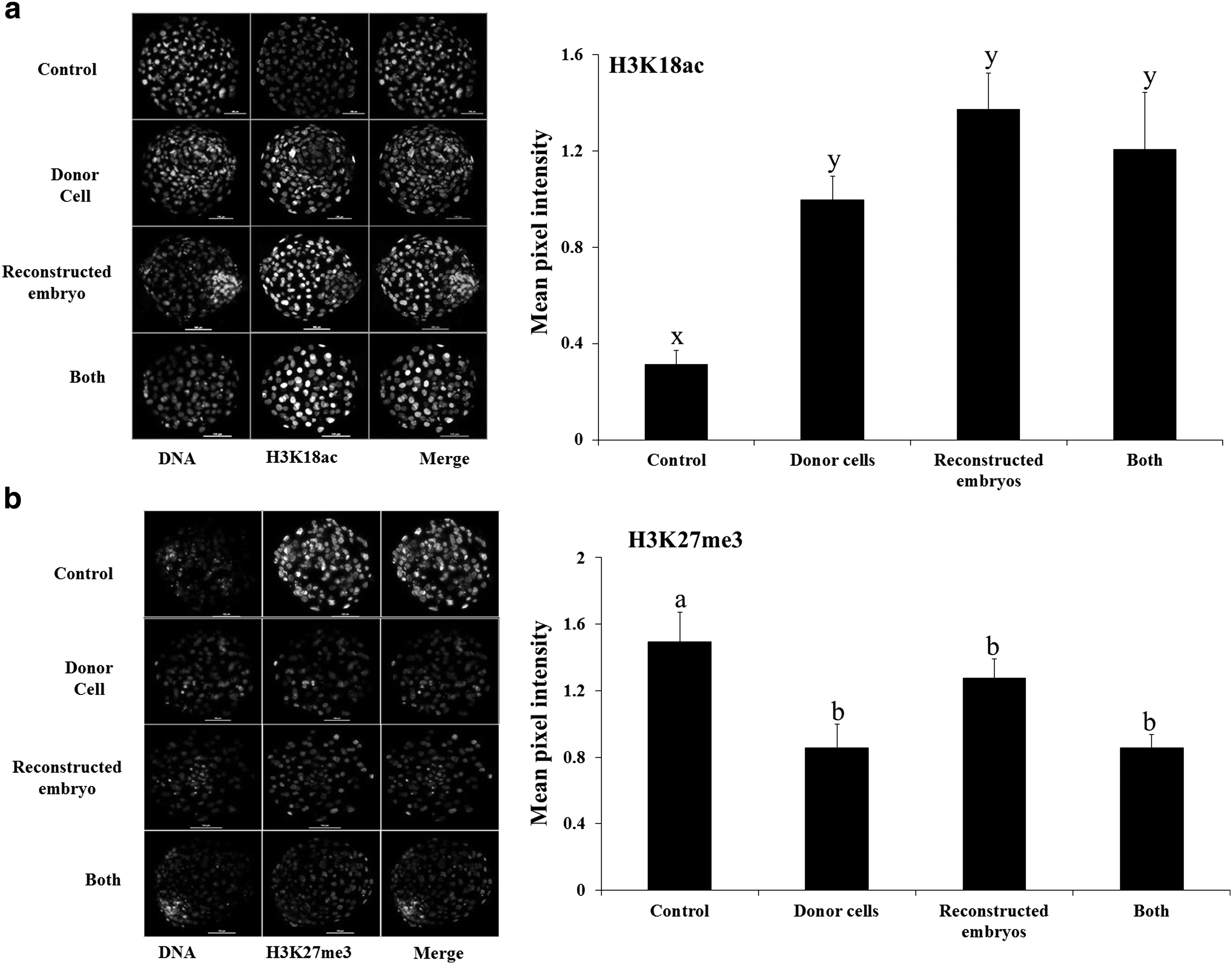
Global level of
Effect of treatment of donor cells or cloned embryos or both with TSA +5-aza-dC on the expression level of some important genes
Treatment of either donor cells or reconstructed embryos or both with TSA +5-aza-dC decreased (p < 0.05) the expression level of epigenetic-related genes DNMT1 and DNMT3a in cloned blastocysts compared with that in control blastocysts, in which the expression level was lower (p < 0.05) than in IVF blastocysts (Fig. 2). The expression level of HDAC1 was similar among the cloned blastocysts of the control and all the treatment groups and IVF counterparts. Among pluripotency-related genes, the relative expression level of OCT4 and NANOG was similar among cloned blastocysts of all the treatment groups and controls, but was significantly lower (p < 0.05) than in their IVF counterparts.
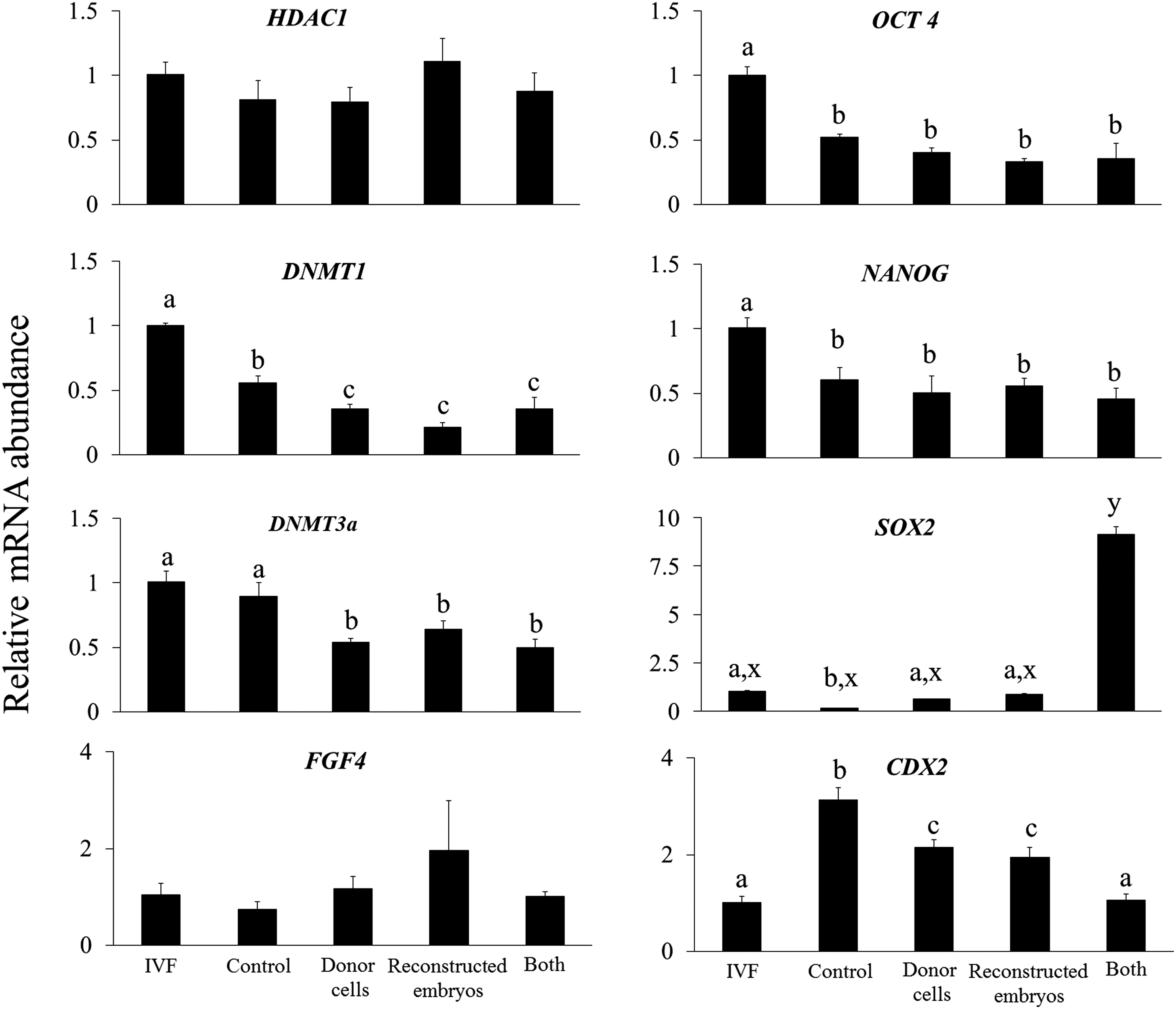
Relative mRNA abundance of HDAC1, DNMT1, DNMT3a, FGF, OCT4, NANOG, SOX 2, and CDX 2 genes in cloned blastocysts produced following treatment of donor cells or reconstructed embryos or both with 50 nM TSA +7.5 nM 5-aza-dC or those produced by IVF. Bars with different superscripts differ significantly (a, b, c: p < 0.05; x, y, z: p < 0.001). IVF, in vitro fertilization.
The expression level of SOX2 was higher (p < 0.01) in cloned blastocysts produced after treatment of both donor cells and reconstructed embryos with TSA +5-aza-dC than in cloned blastocysts of all the other groups and IVF blastocysts. Treatment of either donor cells or reconstructed embryos with TSA +5-aza-dC decreased (p < 0.05) the expression level of trophectoderm-related gene CDX2 compared with untreated controls. However, when both donor cells and reconstructed embryos were treated, the relative expression of CDX2 was decreased (p < 0.05) further to the level in IVF blastocysts. The expression level of development-related gene FGF4 was similar among all the groups (Fig. 2).
Discussion
We investigated whether treatment of both donor cells and reconstructed embryos with TSA and 5-aza-dC in combination offered any advantage over treatment of either donor cells or reconstructed embryos with either of these two epigenetic modifiers individually. However, since the effects of these epigenetic modifiers are not only species specific but also dependent upon the concentrations employed (Cervera et al., 2009), we first optimized the doses of both these epigenetic modifiers. In the first experiment, we examined the effects of treatment of reconstructed embryos with different concentrations of TSA and 5-aza-dC individually on the developmental competence and the level of apoptosis in cloned embryos. We found that compared with the controls, treatment with 5-aza-dC (7.5 or 10 nM), but not with TSA (25, 50, 75, or 100 nM), improved the blastocyst rate and that treatment with either of them decreased the apoptotic index.
Then, we investigated the effects of treatment of both donor cells and reconstructed embryos with either TSA or 5-aza-dC used individually at different concentrations. We found that 5-aza-dC (7.5 or 10 nM) increased the blastocyst rate and decreased the apoptotic index, whereas TSA (50 or 75 nM) decreased the apoptotic index without affecting the blastocyst rate compared with the controls. Based on these results, we chose the lowest effective dose of the two epigenetic modifiers, that is, 50 nM for TSA and 7.5 nM for 5-aza-dC. In the next experiment, we treated both donor cells and reconstructed embryos with 50 nM TSA and 7.5 nM 5-aza-dC in combination. We found that the blastocyst rate was increased over 1.5-fold (p < 0.05) over that in controls and was not significantly different, irrespective of whether donor cells or reconstructed embryos or both were treated with TSA +5-aza-dC.
In addition, the apoptotic index was significantly lower in all three treatment groups than that in the controls and was similar to that in the IVF blastocysts. There are several reports in which beneficial effects of treatment of donor cells (Enright et al., 2003) or reconstructed embryos (Cui et al., 2011; Iager et al., 2008) with TSA have been observed on the developmental competence of bovine embryos. Similarly, treatment of donor cells with 5-aza-dC has also been reported to improve the blastocyst rate or quality in cattle (Enright et al., 2003). To our information, there is only one study that examined the effects of treatment of both donor cells and cloned embryos with the combination of TSA and 5-aza-dC on in vitro developmental competence. Ding et al. (2008) reported that treatment of donor cells, cloned embryos, and continuous treatment of both donor cells and cloned embryos with a combination of 10 nM 5-aza-dC and 50 nM TSA significantly increased the blastocyst rate and TCN in cattle.
Our results extend the findings of this study by demonstrating that because the use of the combination of TSA and 5-aza-dC results in similar levels of increase in blastocyst rate and decrease in the apoptotic index, irrespective of whether donor cells or reconstructed embryos or both are treated, there is no advantage in treating both donor cells and reconstructed embryos when TSA and 5-aza-dC are used in combination. Treating either donor cells or reconstructed embryos with the combination of TSA and 5-aza-dC offers a similar level of beneficial effects.
In the next experiment, we examined the effects of treatment of donor cells or reconstructed embryos or both with the combination of TSA and 5-aza-dC on the global level of two important histone modifications, that is, acetylation of histone 3 at lysine 18 (H3K18ac) and trimethylation of histone 3 at lysine 27 (H3K27me3). Acetylation and methylation of specific lysine residues on the histone molecule play an important role in transcriptional activation and repression of genes, and in general, acetylation is associated with gene activation and methylation with inhibition of gene expression (Valls et al., 2005). TSA treatment of donor cells or NT bovine embryos has been reported to increase acetylation of several important histone lysines such as H3K9 (Jafarpour et al., 2011), H4K5 (Iager et al., 2008; Lee et al., 2011), and H4K12 (Akagi et al., 2011) in cloned embryos. We found that treatment of donor cells or reconstructed embryos or both with the combination of TSA and 5-aza-dC increased the global level of H3K18ac to a similar extent compared with that in the controls.
H3K27me3 plays an important role in embryonic development. Demethylation of H3K27me3 before and after embryonic genome activation has been found to be necessary to ensure normal embryonic development of bovine IVF embryos (Canovas et al., 2012). We found that treatment of donor cells or reconstructed embryos or both with the combination of TSA and 5-aza-dC decreased the global level of H3K27me3 to a similar extent. These results further substantiate those of the previous experiment by demonstrating that treatment of either donor cells or reconstructed embryos with the combination of TSA and 5-aza-dC offers similar level of effects on histone modifications at least in terms of the two important epigenetic markers examined in our study.
Finally, we examined the effects of treatment of donor cells or reconstructed embryos or both with the combination of TSA and 5-aza-dC on the expression level of some important genes. We found that treatment with the combination of TSA and 5-aza-dC decreased the expression level of epigenetic-related genes DNMT1 and DNMT3a, which agrees with the results of a previous study (Wang et al., 2011). We also observed that the magnitude of decrease was similar whether donor cells or reconstructed embryos or both were treated.
In addition, the expression level of another epigenetic-related gene HDAC1 was similar among cloned blastocysts of all the treatment groups. The expression level of pluripotency-related genes OCT4, NANOG, and SOX2 was lower and that of trophectoderm-related gene CDX2 was higher in cloned blastocysts compared with their IVF counterparts, as also reported previously (Mohapatra et al., 2015). We found that the expression level of OCT4, NANOG, and CDX2 and that of development-related gene FGF4 was similar among cloned blastocysts of all the treatment groups.
However, the expression level of pluripotency-related gene SOX2 was higher and that of trophectoderm-related gene CDX2 was lower when both donor cells and reconstructed embryos were treated with the combination of TSA and 5-aza-dC than when either of them was treated. Nevertheless, the expression level of a majority of genes examined in the present study was similar following treatment of donor cells or reconstructed embryos or both with the combination of TSA and 5-aza-dC.
In conclusion, the results of this study demonstrate that treatment of either donor cells or reconstructed embryos or both, with the combination of TSA and 5-aza-dC, offers similar level of beneficial effects in terms of improved blastocyst rate and quality. The epigenetic status, in terms of global level of two important epigenetic markers H3K18ac and H3K27me3, and the expression level of several important genes related to epigenetics, pluripotency, and development are also not affected significantly, irrespective of whether donor cells or reconstructed embryos or both are treated with the combination of TSA and 5-aza-dC. Therefore, there seems to be no advantage in treating both donor cells and reconstructed embryos when the combination of TSA and 5-aza-dC is used.
Footnotes
Acknowledgments
The present work was funded by the National Agriculture Innovative Project (NAIP) grant to S.K.S. (C 2-1-(5)/2007) and M.S.C. (C-2067 and 075). N.L.S. and M.S. are recipients of CSIR-SRF fellowship. M.S., N.L.S., and H.A. carried out the HMC experiments. P.P. and S.K.S. designed the experiments. M.S.C., R.S.M., and S.K.S. contributed funds, reagents, and infrastructure. P.P. prepared and revised the manuscript.
Author Disclosure Statement
The authors declare that no conflicting personal or financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.