
Abstract
Abstract
Somatic cell nuclear transfer (SCNT) success is partially hindered by the low epigenetic reprogramming efficiency of the donor cell. Previous studies suggest cellular heterogeneity among donor nuclei in regard to reprogramming potential, which precludes comparison among different strategies to increase cloning success. In this context, we evaluated the effect of using clonal cell populations (CPs) of bovine adult fibroblasts established by single-cell plating in SCNT. Different CPs were evaluated in regard to proliferation rate, senescence level, and chromosome stability, as well as for POU5F1 (POU class 5 homeobox 1) mRNA expression levels. In total, 9 of 24 CPs (37.5%) were successfully expanded in vitro up to the fourth passage and shown to proliferate following cryopreservation, at which time cell analyses were performed. The use of a CP with low senescence level, normal karyotype, and highest POU5F1 expression levels did not improve embryo development rates or quality following SCNT. As previously suggested, this study supports the notion that levels of POU5F1 expression in the donor nucleus do not impact the SCNT results. Notably, the single-cell seeding approach used herein to isolate CPs may be extended to the evaluation of additional predictor markers of reprogrammability success for SCNT in future experiments.
Introduction
T
However, pregnancy losses increase from the time of placentation onward with high rates of placental and fetal anomalies as well as fetal and neonatal deaths. Reportedly, pregnancy and fetal anomalies related to SCNT stem from faulty epigenetic reprogramming of the donor nucleus (Boyer, 2009; Thuan et al., 2010).
Several changes in methodology have been attempted to improve SCNT success, including pretreatment of donor cells, as well as optimization of enucleation, fusion, activation, and embryo culture protocols (Song et al., 2011; Whitworth et al., 2009). However, pretreatment of donor cells and reconstructed zygotes with chromatin-modifying agents such as oxamflatin (Hou et al., 2014; Park et al., 2012), trichostatin and hydralazine (Sangalli et al., 2012), or valproic acid (Song et al., 2013) did not overcome problems derived from incomplete reprogramming. Nonetheless, early cloning studies did show that donor cell origin, stage of differentiation, and stage of the cell cycle play a role in the success of SCNT (Batchelder et al., 2005).
Moreover, heterogeneity within a primary fibroblast population determined the ability of individual cells to be reprogrammed into induced pluripotent stem (iPS) cells through transgene expression methods (Buganim et al., 2012; Guo et al., 2014). Therefore, recent studies in SCNT have sought to improve reprogramming of somatic cells by identifying cell subgroups within culture with an intrinsic potential to be reprogrammed.
For instance, selection of donor goat fibroblasts expressing the pluripotency marker stage-specific embryonic antigen 3-positive (SSEA3+) enhanced the in vitro development and quality of the resulting SCNT goat embryos (Liu et al., 2013). Similarly, mRNA expression levels of the embryo-specific pluripotency marker POU class 5 homeobox 1 (POU5F1) were evaluated as a prognostic marker of donor cell reprogrammability in SCNT. However, transcript levels of the POU5F1 gene in four bovine adult fibroblast donor cell lines did not correlate with SCNT results (Rodríguez-Alvarez et al., 2013). Conversely, retroviral transfection of the POU5F1 gene into bovine adult fibroblasts induced chromatin-based epigenetic changes that resulted in an increased number of trophectoderm cells in the resulting cloned embryos (Goissis et al., 2013); these authors concluded that even small changes in the epigenetic background of donor cells may affect SCNT success.
Clearly, the effect of identifying better reprogrammable donor cells based on expression of pluripotency-related genes and the evaluation of this approach on SCNT outcome are still matters of debate. Therefore, in this study, we evaluated the effect of adult fibroblast donor cell selection based on the expression of the POU5F1 transcript as assessed by qPCR on preimplantation development of SCNT bovine embryos. Based on evidences that clonal populations (CPs) of fibroblasts with varying potential for in vitro differentiation can be well established in vitro (Bi et al., 2010; Buganim et al., 2012; Chen et al., 2007), we hypothesized that identifying CPs naturally expressing the POU5F1 transcript could represent a method to generate a more homogeneous cell population of donor cells with better reprogramming potential for SCNT.
Materials and Methods
All chemicals and reagents were purchased from Sigma (St. Louis, MO) and cell culture media from Gibco (Grand Island, NY), unless otherwise stated. Culture dishes and flasks were obtained from TPP (Trasadingen, Switzerland). Incubator conditions for oocytes and embryos, as well as for fibroblast cultures, were maintained at 38.58°C in air with 5% CO2 and maximum humidity.
Fibroblast collection and culture
Fibroblasts were isolated from an ear biopsy (∼1 cm2) obtained postmortem from an adult bull at a slaughterhouse and transported to the laboratory in phosphate-buffered saline at 4°C; saline was supplemented with 100 μg/mL gentamicin. Fibroblasts were harvested from tissue by collagenase digestion (1 mg/mL) for 3 hours at 38.5°C. After enzymatic digestion and disaggregation by pipetting, the cell suspension was passed through a 70-μm diameter pore filter and then washed by centrifugation at 300 g for 10 minutes. Cells were resuspended in IMDM supplemented with 10% FBS (IMDM-10%) and 50 μg/mL gentamicin, plated in flasks (25 cm2), and cultured until reaching confluency.
Establishment of fibroblast clonal populations
The primary fibroblast culture (P-0) was trypsinized, and cells were seeded on a 96-well plate at 10 cells/mL (200 μL of IMDM-10% per well), as previously described (Chen et al., 2007). Three hours after cell plating, wells were examined using an inverted microscope; only wells containing one cell were maintained in the experiment. Once established, CPs were numbered according to the well in which they had grown. A sample of the original primary fibroblast cell population (OP) was cultured in parallel with CPs and served as a control for the experiments.
Three days after single-cell plating, cells were examined using an inverted microscope and the cell number was visually counted to estimate CP doubling time. Proliferating CPs were expanded in 96-well plates and only those reaching confluency were successfully passaged through 24-well dishes (P-2), 35-mm Petri dishes (P-3), and 25-cm2 culture flasks (P-4). The OP was also expanded and passaged through 25-cm2 culture flasks up to the P-4 passage. Then, the CPs and OP were frozen in IMDM supplemented with 20% fetal calf serum and 10% dimethyl sulfoxide and stored in liquid nitrogen for subsequent analysis and SCNT.
Cellular senescence and cytogenetic analyses
The CPs and OP were thawed, and 1 × 104 cells (P-5) were seeded on 24-well dishes and cultured for 4 days. Cell senescence was evaluated using the Senescence β-Galactosidase Staining Kit (Cell Signaling Technology, Danvers, MA), according to the manufacturer's instructions. Fibroblast populations were qualitatively classified according to the overall number of senescent cells (blue color) in five different microscopic fields (100 × magnification) analyzed as low (<5 senescent cells), medium (6–15 senescent cells), and high (>15 senescent cells) senescence levels.
Metaphase spreads were obtained using a standard chromosome extraction protocol (Tagliarini et al., 2009). Briefly, cells were thawed on 25-cm2 flasks (P-5) and expanded to 80%–90% confluence. Cells were first treated with 0.5 μg/mL colcemid (Gibco) for 1 hour. Then, cells were harvested and treated with hypotonic solution (0.075 M KCl) for 10 minutes, followed by fixation with methanol:acetic acid (4:1, v:v). Cells were spread onto slides and stained with Giemsa for metaphase analysis. Chromosome content for the CPs and OP was assessed under a microscope (400 × ), and the percentage of euploidy, aneuploidy, and polyploidy was recorded after analysis of 25–30 metaphases.
Somatic cell nuclear transfer
SCNT was performed as described by Miranda et al. (2012), with few modifications. Briefly, the CPs and OP were allowed to grow and were serum-starved (0.5% fetal bovine serum [FBS]) for 24 hours before nuclear transfer to arrest the cell cycle at the G1/G0 stages. For each experiment, fibroblasts from each group at the same cell passage were used as nuclear donors. Nuclear transfer experiments were performed in triplicate.
Bovine ovaries were obtained from a local slaughterhouse and transported to the laboratory in saline solution at 38.5°C. Cumulus oocyte complexes (COCs) were aspirated using a syringe and 18G needle from 3 to 6 mm antral follicles. Immature COCs with compact granulosa cells were cultured in Tissue Culture Medium 199 (TCM199) supplemented with 10% FBS, 0.2 mM pyruvate, 50 μg/mL gentamicin, 0.5 mg/mL FSH, and 150 mg/mL LH. Eighteen hours after the start of maturation, cumulus cells were completely removed from the oocyte by manual pipetting in the presence of 0.2% hyaluronidase, and oocytes with an extruded first polar body were selected for enucleation. Selected oocytes were labeled with 10 μg/mL of the DNA fluorochrome Hoechst 33342 for 10 minutes at room temperature in SOF medium containing 0.2 mM pyruvate and 3 mg/mL bovine serum albumin (BSA) (Wells et al., 1999).
Labeled oocytes were washed and transferred to a micromanipulation drop of SOF medium supplemented with 7.5 μg/mL cytochalasin B. All micromanipulations were performed on an inverted microscope (Leica, Solms, Germany) equipped with Hoffman optics and Narishige micromanipulators (Tokyo, Japan). The first polar body and metaphase II plates were removed by aspiration with a 15-μm inner diameter enucleation pipette (ES TransferTip; Eppendorf). To ensure that the oocyte chromatin was removed, the aspirated cytoplasm was exposed to ultraviolet light and examined for the presence of the removed polar body and metaphase plate.
After enucleation, one single donor cell from a CP or the OP was placed into the perivitelline space of each enucleated oocyte. Then, SCNT-reconstructed embryos were placed into a fusion chamber filled with 0.3 M mannitol, and one pulse of alternating current (5 seconds, 50 V/cm) followed by two continuous current electric pulses (45 μs each, 1.75 kV/cm) were applied to promote fusion in a Multiporator® (Eppendorf, Hamburg, Germany). Around 26 hours after the start of maturation, successfully fused SCNT couplets were activated with 5 μM ionomycin in TCM199-HEPES (supplemented with 1 mg/mL fatty acid-free BSA) for 5 minutes and moved into TCM199-HEPES medium (supplemented with 30 mg/mL BSA) for 5 minutes.
Then, couplets were cultured in SOF medium supplemented with 6-dimethylaminopurine for 3 hours, as previously described (Susko-Parrish et al., 1994). Following activation, reconstructed embryos were washed and placed into 100-μL drops (20–25 embryos) of in vitro culture SOF medium (Vajta et al., 1999). Cleavage and blastocyst rates were evaluated on the second and seventh day, respectively. Nonmanipulated, control metaphase II-arrested oocytes were activated as above and cultured in vitro using the same protocol (parthenogenetic embryos) and used as a control for oocyte quality, activation procedure, and in vitro culture. On day 7, SCNT and control blastocysts were snap-frozen in liquid nitrogen and kept at −80°C for gene expression analysis.
Gene expression analysis
On the fifth passage, both the CPs and control OP were allowed to grow to 80% confluency on 25-cm2 culture flasks. Then, total RNA was extracted using TRIzol (Invitrogen, Molecular Research Center, Cincinnati, OH) following the manufacturer's instructions, including treatment with amplification-grade DNAse I (Thermo Scientific, Rochester, NY). Purified RNA was quantified using a NanoDrop spectrophotometer (Thermo Scientific) and 1 μg of RNA was converted to cDNA using Multiscribe Reverse Transcriptase (Applied Biosystems), according to the manufacturer's instructions.
For gene expression analysis in cloned blastocysts, pools of five embryos were collected and RNA was extracted using TRIzol, including treatment with amplification-grade DNAse I (Thermo Scientific). Purified RNA was converted to cDNA using the High-Capacity cDNA Reverse Transcription Kit (Life Technologies).
Real-time PCR was performed using a StepOnePlus Real-Time PCR system (Applied Biosystems). Primers were designed using Primer Premier software (Premier Biosoft International, Palo Alto, CA) based on cDNA sequences available in GenBank. Primer specificity was checked by BLAST, and sequences were as follows:
POU5F1, sense 5′-CAGGCCCGAAAGAGAAAGC-3′, antisense 5′-CGGGCACTGCAGGAACA-3′; GAPDH, sense 5′-AAGGCCATCACCATCTTCCA-3′, antisense 5′-CCACTACATACTCAGCACCAGCAT-3′; YWHAZ, sense 5′-GCATCCCACAGACTATTTCC-3′, antisense 5′-GCAAAGACAATGACAGACCA-3′; and SDHA, sense 5′-GCAGAACCTGATGCTTTGTG-3′, antisense 5′-CGTAGGAGAGCGTGTGCTT-3′. The PCR total reaction volume was 10 μL, containing 5 μL of SYBR Green Master Mix (Applied Biosystems), 0.5 mM of each primer, and 4 μL of DNA diluted 1:8. The thermocycling conditions consisted of a cycle at 95°C for 10 minutes, followed by 45 cycles at 60°C for 1 minute.
The specificity of PCR products was verified by evaluating the corresponding dissociation curves. The amplification efficiency for each gene was calculated using a relative standard curve of serial dilutions of cDNA (1:4, 1:8, 1:16, 1:32, and 1:64). The relative quantity of the POU5F1 gene was corrected relative to the expression of the housekeeping genes glucose-6-phosphate dehydrogenase (GAPDH) in fibroblasts and tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta polypeptide (YWHAZ) and succinate dehydrogenase flavoprotein subunit (SDHA) in embryos due to their high stability in blastocysts (Goossens et al., 2005). After normalizing to the endogenous gene expression (ΔCt = target gene Ct − endogenous gene Ct) and then to the control group (ΔΔCt = treatment ΔCt−C ΔCt), relative quantification using 2−ΔΔCt was calculated as a fractional change of target mRNA expression in treated versus control.
Statistical analysis
Differences in cleavage rates, blastocyst formation, and gene expression were first submitted to the normality test and evaluated by ANOVA and Fisher's LSD post-test using the SigmaPlot 11.0 software. Values with p < 0.05 were considered significant.
Results
Establishment and characterization of fibroblast clonal populations
Three hours after single-cell plating, 24 of the 96 (25.0%) wells were observed to contain a single cell. Following 3 days of culture, there were marked differences among CPs in terms of cell proliferation (Table 1), with populations ranging from 2 to more than 50 cells. Moreover, morphological differences were also observed, including CPs with binucleated and senescent cells (Fig. 1).
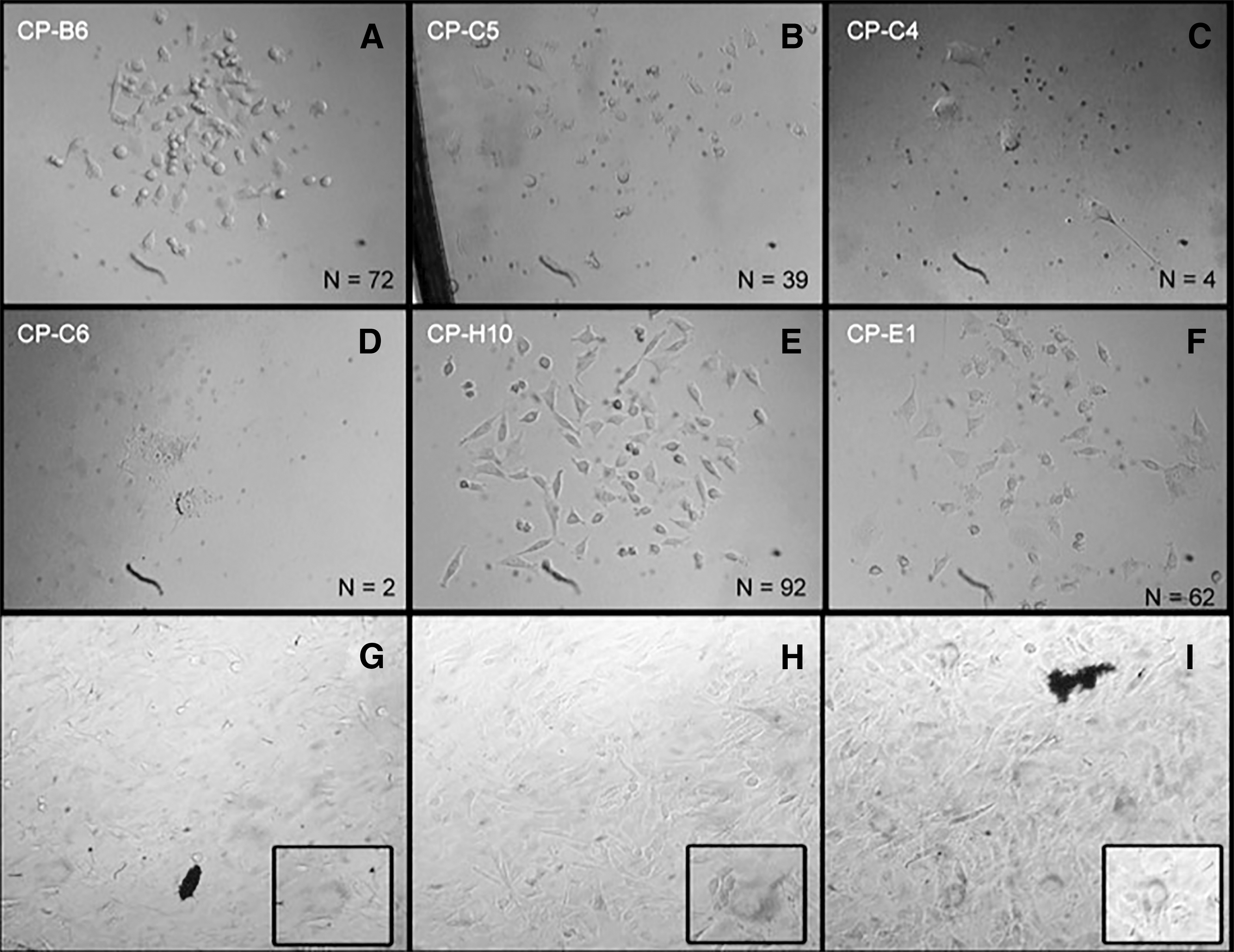
CPs of bovine fibroblasts on the third day following single-cell plating. CPs exhibited a fibroblast-like appearance, with differences related to cell cycle kinetics (cell number) and morphology. Some CPs presented binucleated cells (CP-C6). N = number of cells counted. Magnifications were CP-B6, CP-C5, CP-H10, CP-E1 (100 × ), CP-C4, CP-C6 (400 × ).
Doubling time was calculated 72 hours after single-cell plating. Cytogenetic and senescence level analyses were only performed in CPs that proliferated following thawing.
CP, clonal population; OP, original population.
The calculated doubling time of CPs on the third day of culture is shown in Table 2. From the 24 CPs generated after single-cell plating, 18 (75.0%) reached P-2, 16 reached P-3 and P-4 (66.6%), and only 14 (58.3%) were able to be expanded up to the cryopreservation step at P-4. Six of the 14 frozen CPs showed growth arrest or low proliferation capacity after thawing and were not used in the experiments. Therefore, only the nine CPs that successfully proliferated after thawing were used in experiments. Altogether, these findings supported the existence of cellular heterogeneity in the primary fibroblast culture.
Cellular senescence and cytogenetic analysis
Senescent cells were present in all fibroblast populations evaluated, including the OP (Fig. 1). Senescence level was low for the control group (OP), but varied among CPs (Table 1).
Cytogenetic analysis results of the OP and nine CPs are summarized in Table 1. In general, CPs had fewer euploid metaphases when compared with the OP. Moreover, some CPs showed high levels of aneuploidy (i.e., CP-H10; 53.5%) and polyploidy (i.e., CP-E1; 67.8%). However, some CPs had percentages of ploidy similar to the OP. For instance, the percentage of aneuploidy–polyploidy of the CP-D11 was similar to that in the OP (35.7% vs. 33.3%, respectively). In fact, the CP-G11 had a percentage of euploidy higher than that in the OP (81.2% vs. 66.6%, respectively).
Gene expression analysis
The expression levels of POU5F1 varied among CPs (Fig. 2). In four of the CPs tested, expression levels for this gene were higher than those in the OP, with the highest expression observed in CP-G11. Conversely, for two of the CPs, expression levels of POU5F1 were lower than those in the OP.
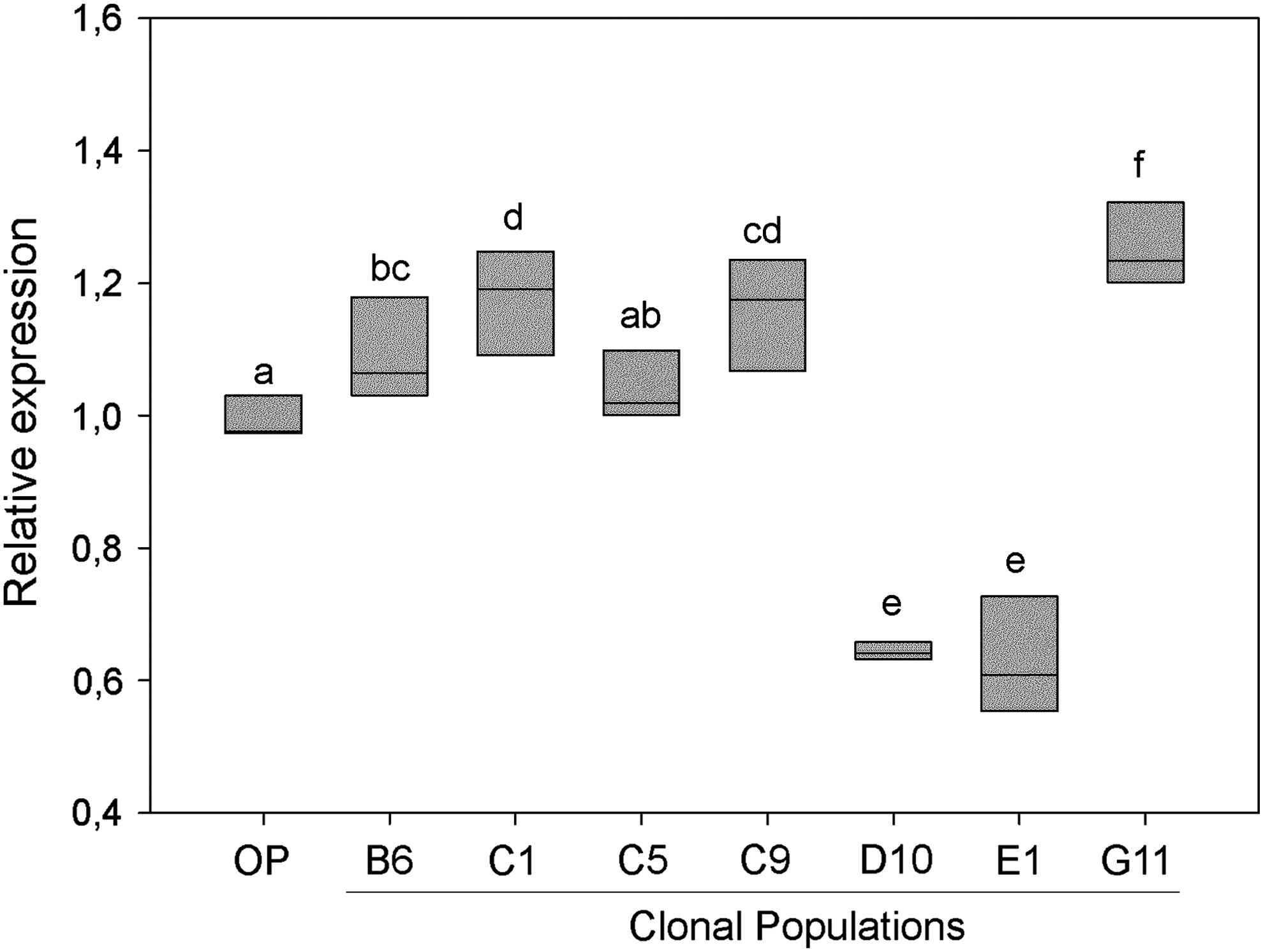
Relative POU5F1 expression in different clonally isolated bovine fibroblast populations. Relative POU5F1 expression was corrected relative to that of the housekeeping gene GAPDH. Relative expression (2−ΔΔCt) was calculated as a fractional change of target mRNA expression in the clonal populations versus that in the OP. Primer efficiencies were above 96% and detected Cq values for POU5F1 varied from 24.6 to 26.6. Different letter superscripts denote significance (p < 0.05). GAPDH, glucose-6-phosphate dehydrogenase; OP, original population; POU5F1, POU class 5 homeobox 1.
Somatic cell nuclear transfer
Based on the CP analysis results, we selected a population that consistently exhibited characteristics of interest for successful SCNT. In this regard, CP-G11 displayed low incidence of senescence, high level of euploidy, and increased expression of the pluripotency marker POU5F1. Conversely, CP-E1 was chosen for comparison given its opposite phenotype. Control embryos were reconstructed with fibroblasts from the OP. Notably, no difference (p > 0.05) was observed regarding cleavage and blastocyst rates between SCNT embryos using donor cells from the OP or either of the two CPs (Table 2). Moreover, SCNT embryos derived from both CPs (CP-E1 and CP-G11) showed decreased POU5F1 gene expression levels (p < 0.05) compared with those of the control group (OP), with no differences between the two CPs (Fig. 3).
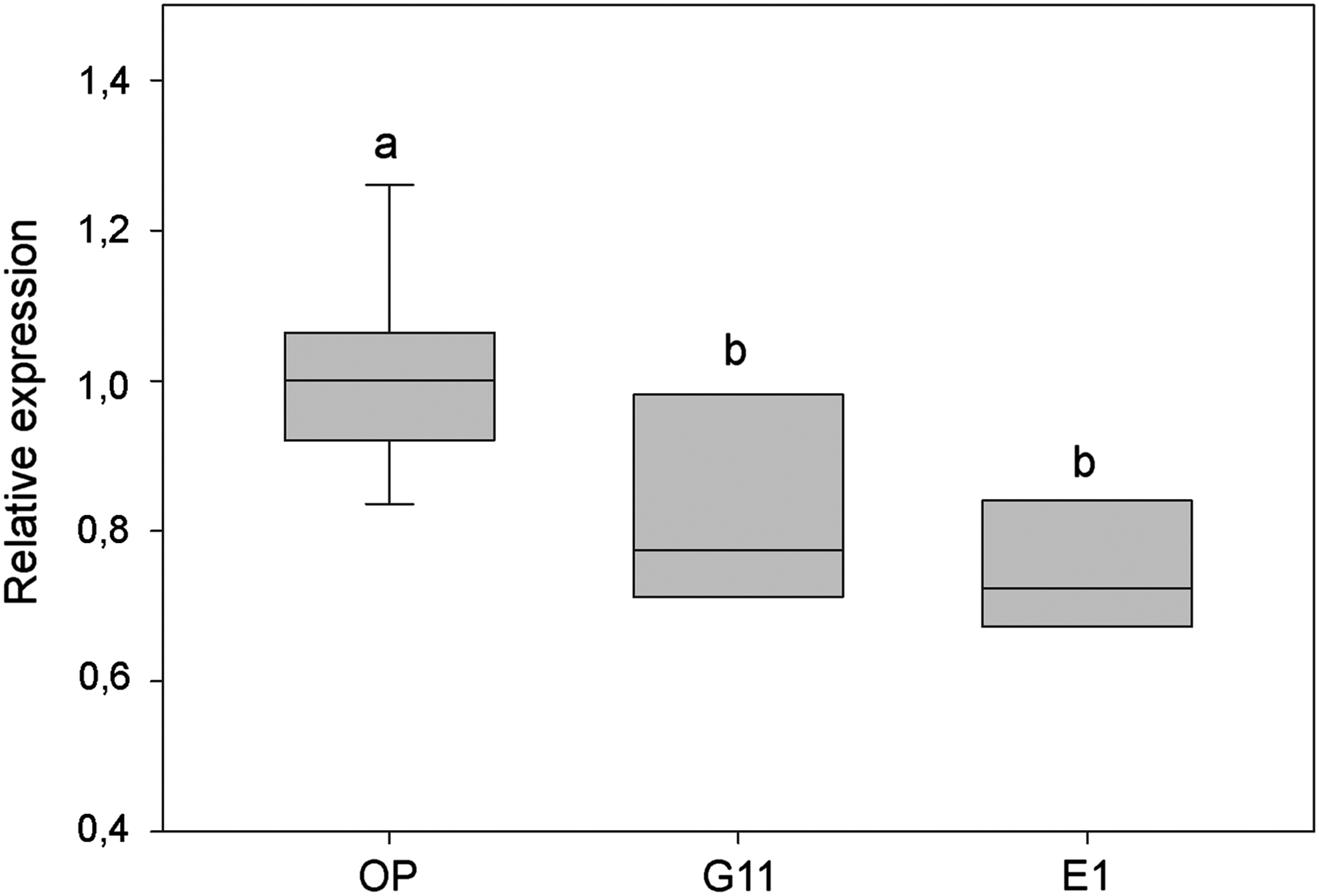
POU5F1 gene expression of somatic cell nuclear transfer embryos reconstructed with fibroblasts from the OP or from two CPs. Relative POU5F1 expression was corrected relative to that of the housekeeping gene GAPDH. Relative quantification (2−ΔΔCt) was calculated as a fractional change of target mRNA expression in embryos from CP cells versus those reconstructed with OP cells. Different letter superscripts denote significance (p < 0.05).
Discussion
In this study, we sought to investigate whether the heterogeneity of a primary population of adult bovine fibroblasts to be used as nuclear donors had an effect on the resulting SCNT-derived embryos. For this purpose, single-cell plating was used as a means to obtain different clonal populations (CPs) representing the heterogeneity of the primary culture in regard to their intrinsic reprogramming potential, a strategy already proven to be effective (Buganim et al., 2012; Chen et al., 2007). Indeed, one of the main issues impacting the efficiency of somatic cell reprogramming by SCNT is the heterogeneity of the donor cell population (Batchelder et al., 2005; Cheong, 2008), which, in turn, yields unpredictable outcomes in regard to live offspring and makes comparison among different protocols/strategies unfeasible.
Moreover, a growing body of research suggests that the successful reprogramming of a given somatic cell may depend more on its innate state (deterministic model) rather than on a random process (stochastic model), as has long been presumed (Chen et al., 2007; Niibe et al., 2011; Pour et al., 2015). Therefore, herein, cells from the different CPs were evaluated in regard to proliferation potential, senescence level, chromosomal complement, and level of expression of the pluripotency marker POU5F1 in hopes to identify any intrinsic pattern that could predict successful reprogramming by SCNT.
Given that senescent phenotypes of transgenic donor fibroblasts negatively affected SCNT efficiency in pigs and that there is a clear negative impact of using defective cells as nuclear donors (Loi et al., 2002; Miranda et al., 2012), both senescence level and chromosomal number were chosen as evaluation parameters for nuclear donor cells in this study. Notably, even following thousands of cell cycles and after five passages, we were able to derive low senescent CPs that displayed chromosomal stability, with levels of euploidy similar to those observed in the original fibroblast population.
Moreover, CPs that retained their proliferating ability following frozen storage were also analyzed for POU5F1 transcript expression levels to test the hypothesis that a higher natural expression of POU5F1 would be positively correlated with cloning results. In this regard, it has been suggested that expression of certain genes in donor cells facilitates their expression in the resulting SCNT embryos. Furthermore, several studies have pursued the selection of markers that might be associated with good reprogramming efficiency, both for SCNT and iPS cell generation experiments (Cheong, 2008; Liu et al., 2013; Pan et al., 2015). In particular, POU5F1 expression has been assessed as a candidate donor cell marker relating to SCNT success, although with conflicting results (Goissis et al., 2013; Kim et al., 2015; Rodríguez-Alvarez et al., 2013).
In our study, a specific clonal population (CP-G11) displayed the highest percent of euploidy (81.82%) combined with a low senescence level and highest expression levels of POU5F1. However, rates of blastocyst development of SCNT bovine embryos reconstructed with cells from this clone were similar (p > 0.05) to those observed in the original population. Unexpectedly, SCNT results for CP-G11 were also similar to those obtained with the CP-E1 clone, which displayed the highest polyploidy combined with a high senescence level and low POU5F1 expression.
Moreover, expression levels for POU5F1 in the resulting blastocysts were not influenced by the relative expression levels, whether low or high, in the corresponding donor fibroblast population. These findings suggest complete epigenetic resetting of POU5F1 expression during the initial stages of embryonic development. Furthermore, given that expression levels of POU5F1 have been positively correlated with embryo quality (Santana et al., 2014), our results imply that presumptive embryo quality does not improve by selecting the donor cell, at least based upon the parameters used herein. In fact, our results agree with those of a previous report showing that expression levels for POU5F1 in five donor cell lines (from five different adult bovines) did not correlate with successful in vitro development of the resulting SCNT-derived embryos (Rodríguez-Alvarez et al., 2013).
However, contributing to the controversy in the field, reports on artificially induced overexpression of POU5F1 in the donor cells for SCNT have shown some beneficial effects related to increased trophectoderm cells in bovine embryos (Goissis et al., 2013) and higher development rates in pig embryos (Kim et al., 2015). Last, but not least, the expression of the POU5F1 gene in somatic tissues itself can be considered controversial once the existence of pseudogenes, which could contribute to false-positive gene expression results (Poursani et al., 2016). In our study, we cannot exclude that possibility; however, in bovines, only one POU5F1 pseudogene has been described so far and its mRNA was not detected either in somatic cells or embryos (Rodríguez-Alvarez et al., 2013; Yadav et al., 2005).
Moreover, expression of POU5F1 in adult tissues has been described previously (Mizuno and Kosaka, 2008; Pochampally et al., 2004; Zangrossi et al., 2007), and recently some evidences have been showing that epiblastic cells may spread out into the body of developing embryo colonizing somatic tissues and being maintained as bona fide pluripotent cells until the adult life (Ratajczak et al., 2012, 2014). Therefore, future studies should address the in vivo potential of embryos derived from donor cells expressing either natural or induced higher levels of POU5F1.
Other potential candidate markers of reprogrammability potential for SCNT donor cells include SSEA3+ (Liu et al., 2013) and SSEA4+ (Pan et al., 2015), with promising results in terms of in vitro development of the resulting cloned embryos. However, these studies used a different approach, in that SSEA3+ and SSEA4+ fibroblasts were first sorted by flow cytometry and then used as donor cells for SCNT. Conversely, herein, we first established CPs and then, for each, we investigated a candidate cell marker together with basic cellular characteristics such as ploidy and senescence to choose our donor cells.
Therefore, considering the simplicity of the single-cell plating approach compared to performing flow cytometry, it would be interesting to analyze other candidate genes (such as SSEA types) in successfully established single-cell lines before using them as nuclear donors for SCNT. Conversely, the single-cell approach could be used in a retrospective study where established cell lines would be first tested for SCNT success and then submitted to DNA array analysis to identify gene expression profiles correlating with high reprogramming efficiency.
With regard to the effects of cell population doubling rates and donor cell size on SCNT success, it was previously shown that clonal populations of porcine fetal fibroblasts with low population doubling times (∼19 hours) and large size (∼20 μm) yielded better results in terms of number and quality of blastocysts produced (Cheong, 2008). Conversely, in a recent study on mice, cell cycle speed was positively correlated with the privileged state of somatic cells in reprogramming experiments, with most iPS cell colonies deriving from somatic cells with a cell cycle of ∼8 hours (Guo et al., 2014). In our study, the estimated cell cycle length 3 days after single-cell plating, before the first passage, varied largely among clonal populations, ranging from ∼11.0 to ∼34 hours. However, the effect of this phenotypical heterogeneity on SCNT results was not investigated and should be addressed in future experiments.
In summary, our results show no relationship between expression levels of POU5F1 in clonal populations of bovine fibroblasts and blastocyst development rates following SCNT, supporting the notion that the level of POU5F1 expression in somatic cells is not the best indicator of SCNT success. The fact that we were able to obtain clonal populations with chromosomal stability and low senescence levels indicates that the single-cell seeding approach used herein can be extended to the evaluation of additional predictors of reprogrammability success for SCNT and iPS experiments.
Footnotes
Acknowledgments
This work received financial support from the CAPES Foundation, Ministry of Education, Brazil, Pará State Foundation for Research, FAPESPA, Brazil, and from São Paulo State Foundation for Research, FAPESP, Brazil. The authors thank Dr. Sylvia J. Bedford-Guaus for critical reading of the manuscript.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.