
Abstract
Abstract
We evaluated the suitability of 10 candidate internal control genes (ICGs), belonging to different functional classes, namely ACTB, EEF1A1, GAPDH, HPRT1, HMBS, RPS15, RPS18, RPS23, SDHA, and UBC for normalizing the real-time quantitative polymerase chain reaction (qPCR) data of blastocyst-stage buffalo embryos produced by hand-made cloning and in vitro fertilization (IVF). Total RNA was isolated from three pools, each of cloned and IVF blastocysts (n = 50/pool) for cDNA synthesis. Two different statistical algorithms geNorm and NormFinder were used for evaluating the stability of these genes. Based on gene stability measure (M value) and pairwise variation (V value), calculated by geNorm analysis, the most stable ICGs were RPS15, HPRT1, and ACTB for cloned blastocysts, HMBS, UBC, and HPRT1 for IVF blastocysts and RPS15, GAPDH, and HPRT1 for both the embryo types analyzed together. RPS18 was the least stable gene for both cloned and IVF blastocysts. Following NormFinder analysis, the order of stability was RPS15 = HPRT1>GAPDH for cloned blastocysts, HMBS = UBC>RPS23 for IVF blastocysts, and HPRT1>GAPDH>RPS15 for cloned and IVF blastocysts together. These results suggest that despite overlapping of the three most stable ICGs between cloned and IVF blastocysts, the panel of ICGs selected for normalization of qPCR data of cloned and IVF blastocyst-stage embryos should be different.
Introduction
S
Also, fetal loss, neonatal health problems, and death are more prevalent than in conventional methods of reproduction. Not only do a majority of cloned embryos die during pre- and postimplantation development, but also those which develop to term suffer from abnormalities, such as large offspring syndrome, short lifespan, severe placental deficiency, respiratory problems, prolonged gestation, dystocia, etc. (Niemann, 2016; Panarace et al., 2007). It is believed that the primary reason behind these problems are abnormal epigenetic modifications such as DNA and histone hypermethylation, abnormal histone modifications, and incomplete nuclear reprogramming of donor nuclei by the oocyte cytoplasm (Dean et al., 2001). Nuclear reprogramming is a complex process, which involves abolishing the expression profile of a differentiated cell and establishment of a new embryo-specific expression profile, involving 10,000 to 12,000 genes, which drives embryonic and fetal development (Niemann, 2016).
Many studies have now shown similar gene expression in bovine blastocysts produced in vivo or by in vitro or SCNT procedures (Betsha et al., 2013; Min et al., 2015; Nagatomo et al., 2015; Zhou et al., 2008). Unusual epigenetic modifications result in abnormal expression of several genes, which are reflected in alterations in gene expression in preimplantation embryos (Lazzari et al., 2002). Therefore, understanding gene expression patterns is critical to provide insights into complex regulatory networks and for identification of genes relevant to different types of biological processes involved in the events of embryonic development. Among the array of techniques available, real-time quantitative polymerase chain reaction (qPCR) is a well-established and widely used technique to quantify precise mRNA levels of target gene of interest in different biological samples (Bustin, 2000). Due to high reliability of qPCR results, this technique has also become the preferred method to validate the results obtained from high throughput analyses like microarray and RNA-seq.
However, the expression data from qPCR studies needs normalization as this technique is prone to analytical variations (Dheda et al., 2005). To achieve accurate quantification, it is quite essential to take into account the variations that might occur due to differing amount of starting material, pipetting errors, efficiencies of RNA extraction, and reverse transcription (Hugget et al., 2005; Radonic et al., 2004). Use of reference genes or internal control genes (ICGs) with constant expression level between samples, in response to experimental treatment or physiological state, is considered an effective method for normalization of transcriptional data to account for the experimental variations (Bustin et al., 2009).
However, these reference genes should be identified for every experimental design or cell/tissue types, because the use of inappropriate ICGs may lead to erroneous results (Dheda et al., 2005; Glare et al., 2002; Schmittgen and Zakrajsek, 2000; Tricarico et al., 2002). The conventionally used reference genes like β-actin, GAPDH, and rRNAs as normalizers have often been found to show variable expression in many experimental conditions and are not ideally suited for all study types (Bustin and Nolan, 2004; Glare et al., 2002; Hugget et al., 2005; Toshihide et al., 2000). Although there are several studies in which a single reference gene has been used as an internal control, using a single reference gene for normalization has been shown to be inappropriate (Tricarico et al., 2002), and an optimal number and choice of the internal controls must be determined experimentally (Andersen et al., 2004; Pfaffl et al., 2004; Vandesompele et al., 2002).
In view of the importance of cloning and IVF technologies, it is necessary to evaluate transcriptional profile of key genes in blastocyst-stage embryos, produced through these technologies. Although some studies have investigated reference genes in bovine SCNT and IVF embryos (Luchsinger et al., 2013; Ross et al., 2010), to our knowledge, not much information is available on set of reference genes that can be used in expression studies involving hand-made cloning (HMC) and IVF-produced blastocyst-stage embryos of buffaloes. In the present study, therefore, we evaluated 10 commonly used reference genes for selecting the panel of stable reference genes to avoid variation and errors in expression data of HMC and IVF-produced buffalo blastocyst-stage embryos.
Materials and Methods
All the chemicals and media used in the present study were obtained from Sigma Chemical Co. (St. Louis, MO), and all the plasticware were purchased from Nunc (Rosklide, Denmark), unless indicated otherwise. Fetal bovine serum (FBS) was purchased from Hyclone (Logan, UT).
Production of blastocysts by HMC
Usable quality cumulus–oocyte complexes (COCs) obtained from abattoir buffalo ovaries were washed three times with Ca2+ and Mg2+-free Dulbecco's phosphate-buffered saline (DPBS) containing 0.4% bovine serum albumin (BSA) and 50 μg/mL gentamicin sulfate. For selection of developmentally competent oocytes, the COCs were incubated with 26 μM Brilliant Cresyl Blue (BCB) for 90 minutes at 38.5°C in a CO2 incubator as described previously (Mohapatra et al., 2015). The COCs were then washed three times with DPBS and those with blue color (BCB+) were used for in vitro maturation (IVM). The maturation medium consisted of TCM-199 containing 10% FBS, 5 μg/mL porcine follicle-stimulating hormone, l μg/mL estradiol-17β, 0.81 mM sodium pyruvate, 10% buffalo follicular fluid, and 50 μg/mL gentamicin sulfate. Ear skin fibroblast cells from a buffalo bull (Mu-5926), which had already been established, characterized and cryopreserved in our laboratory (Selokar et al., 2014), were used as donor cells for HMC.
The cells were synchronized in G1 stage of cell cycle by growing them in culture to full confluence for contact inhibition as described previously (Selokar et al., 2012a). HMC, which included IVM, cumulus/zona removal, manual enucleation, fusion, activation, and in vitro culture (IVC), was performed as described earlier (Selokar et al., 2012b).
Production of IVF blastocysts
Immature buffalo oocytes obtained from slaughterhouse ovaries were subjected to BCB staining as described above. BCB+ oocytes were subjected to IVM and IVF as described previously (Sharma et al., 2011). To produce genetically half-identical IVF blastocysts, semen was procured from the same buffalo bull (Mu-5926) from which somatic nuclear donor cells were obtained for HMC. IVC of presumptive zygotes was carried out in Research Vitro Cleave Medium (K-RVCL-50; Cook, Queensland, Australia) supplemented with 1% fatty acid-free BSA for 8 days at 38.5°C in a CO2 incubator on original bed of granulosa cells, which were obtained from COCs during IVM. Day 8 blastocysts produced by HMC and IVF were stored at −80°C until RNA isolation.
RNA isolation and cDNA synthesis
Total RNA was isolated from three pools of blastocysts produced by HMC and IVF (n = 50 per pool) using the Norgen's Single Cell RNA Isolation Kit (Thorold, ON, Canada) as per the manufacturer`s protocol. Genomic DNA was removed by on-column DNA removal protocol using the Norgen's RNase-Free DNase Kit. Total RNA concentration and purity was measured by NanoQuant (Tecan, Mannedorf, Switzerland). The total RNA isolated from each pool of blastocysts exhibited high purity as determined by A260/280 ratio, which was between 1.8 and 2.1. cDNA was prepared using the SuperScript III Kit (Invitrogen, Carlsbad, CA) with 100 ng RNA, 1 μL dT12−18, 1 μL 10 mM dNTP mix, 1 μL random primers, and 10 μL DNase–RNase-free water.
The mixture was incubated at 65°C for 5 minutes and then kept on ice for 3 minutes. A total of 6 μL of Master Mix, composed of 4.5 μL 5 × First-Strand Buffer, 1 μL 0.1 M DTT, 0.25 μL (50 U) of SuperScript III RT, and 0.25 μL of RNase inhibitor (10 U; Promega, Fitchburg, WI), was added. The reaction was performed using the program: 25°C for 5 minutes, 50°C for 60 minutes, and 70°C for 15 minutes. cDNA was then diluted 1:4 (v:v) with DNase–RNase-free water.
Selection of reference genes
Ten reference genes, belonging to different functional classes, were selected for evaluation: β actin (ACTB), eukaryotic translation elongation factor 1 alpha 1 (EEF1A1), glyceraldehyde 3-phosphate dehydrogenase (GAPDH), hypoxanthine-guanine phosphoribosyltransferase 1 (HPRT1), hydroxymethylbilane synthase (HMBS), ribosomal protein S15 (RPS15), ribosomal protein S18 (RPS18), ribosomal protein S23 (RPS23), succinate dehydrogenase complex flavoprotein subunit A (SDHA), and ubiquitin C (UBC) (Table 1). The primer sequences were taken from the literature (Denicol et al., 2013; Kapila et al., 2013) (Table 2).
Real-time quantitative polymerase chain reaction
qPCR was performed using StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster City, CA) in a 96-well transparent plate (Thermo Fisher Scientific, Waltham, MA). Each reaction was comprised of 4 μL diluted cDNA combined with 6 μL of a mixture composed of 5 μL (2 × ) Maxima SYBR Green/ROX qPCR Master Mix (Fermentas Thermo Fisher Scientific), 0.4 μL each of 10 μM forward and reverse primers, and 0.2 μL DNase–RNase-free water. For each gene, samples were analyzed in duplicate (technical replicates) along with five-point relative standard curve and the nontemplate control. The amplification conditions used were: 10 minutes at 95°C, 40 cycles of 15 seconds at 95°C (denaturation), and 1 minute at 60°C (annealing + extension).
A dissociation protocol, with incremental temperatures of 95°C for 15 seconds plus 65°C for 15 seconds was used for investigating the specificity of qPCR reaction and presence of primer dimers. Data acquired were computed by the StepOne™ Real-Time PCR Software v2.0 (Applied Biosystems) and subjected to subsequent analysis. The specificity and integrity of PCR products were confirmed by single melt curve peak. The efficiency of PCR reaction was determined by relative standard curve derived from fivefold serial dilutions of pooled cDNA mixture from both cloned and IVF blastocysts.
Evaluation of the stability of expression of reference genes
The stability of expression of each of the 10 candidate genes under study was evaluated using two independent statistical applications: geNorm (Vandesompele et al., 2002) and NormFinder (Andersen et al., 2004). geNorm is a Microsoft excel-based application, which measures the expression stability (M value) based on overall pairwise comparisons among the reference genes. Based upon the M value, which is inversely correlated to gene expression stability, the reference genes are ranked accordingly.
The input data for geNorm were created using the relative quantities based on comparative cycle threshold (Ct) method as described by Livak and Schmittgen (2001) and Vandesompele et al. (2002). Pairwise variation (V) was also calculated using geNorm between the normalization factor of n (best reference) gene (NF n ) and normalization factor of n + 1 (next less stable reference) gene (NFn+1). NormFinder, a model-based approach, was also used for evaluation of the reference genes for which Ct values were transformed to linear scale expression quantities. It determined the best reference genes and gave the corresponding stability value taking into account the variations across different sample groups.
Results
Quality of blastocysts produced by HMC and IVF
A total of 164 blastocysts were produced from 1090 oocytes by IVF in eight trials, the average blastocyst rate being 15.12% ± 2.27%; whereas, in HMC, 171 blastocysts were produced from 431 reconstructs in nine trials, with the average blastocyst rate of 39.97% ± 1.49%. The blastocyst rate obtained was similar to that reported in previous studies in our laboratory (Mohapatra et al., 2015; Selokar et al., 2014). For both embryo types, only those blastocyst-stage embryos were used that were morphologically healthy, that is, which had well-defined blastocoels and a pronounced differentiation of the outer trophoblast layer, and the darker, more compact inner cell mass.
Transcription profiling of reference genes
A single melt-curve peak was obtained for each primer pair, confirming gene-specific amplification. The average Ct value for the candidate reference genes varied between 15.32 (EEF1A1) and 27.66 (RPS15) across both cloned and IVF blastocysts. The Ct values for the 10 genes evaluated are represented as box-and-whisker plot (Fig. 1). Genes were grouped into three different classes on the basis of their average Ct values. Those in class I, which included two genes EEF1A1 and RPS18, with average Ct values of 15.32 and 16.76, respectively, were the most abundant in both cloned and IVF blastocysts.
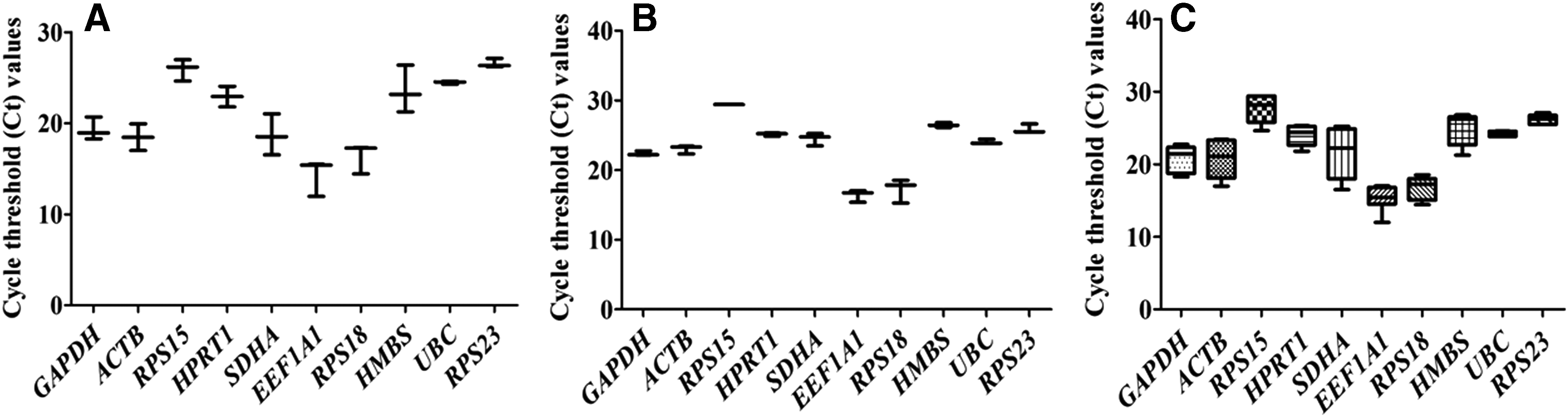
Overall expression pattern of 10 genes evaluated in
Class II included three genes, SDHA, GAPDH, and ACTB, with average Ct values of 21.58, 20.82, and 20.73, respectively, indicating moderate expression, whereas, class III included five genes RPS15, RPS23, HMBS, UBC, and HPRT1 with relatively lower expression and average Ct values of 27.66, 26.20, 25.02, 24.24, and 24.02, respectively. The qPCR performance of each gene in terms of coefficient of determination (R2) and amplification efficiency (E = 10−1/slope), based upon the slope of five-point standard curve, are given in Table 2. The qPCR efficiency ranged between 92.19 (SDHA) and 110.71 (HPRT1), and all the genes gave qPCR efficiency >90% and R2 > 0.97.
Analysis of gene expression stability by geNorm
The gene stability measure (M value) calculated by geNorm analysis ranks the genes from most stable (lowest M value) to least stable (highest M value). The 10 reference genes were ranked for both cloned and IVF blastocysts, together as well as separately, for individual blastocyst type. The reference genes found to be most stable were RPS15 and HPRT1 for cloned blastocysts (M value 0.22), HMBS and UBC for IVF blastocysts (M value 0.13), and GAPDH and RPS15 (M value 0.45) for both cloned and IVF blastocysts together (Fig. 2). RPS18 was found to be the least stable gene for both cloned (M value 1.63) and IVF blastocysts (M value 0.82). To find out the optimal number of genes to be used for determining the normalization factor, pairwise variation, termed as “V value,” was calculated. The combination of two most stable genes was found to be RPS15 and HPRT1 for cloned blastocysts, HMBS and UBC for IVF blastocysts, and GAPDH and RPS15 for the two blastocyst types together.
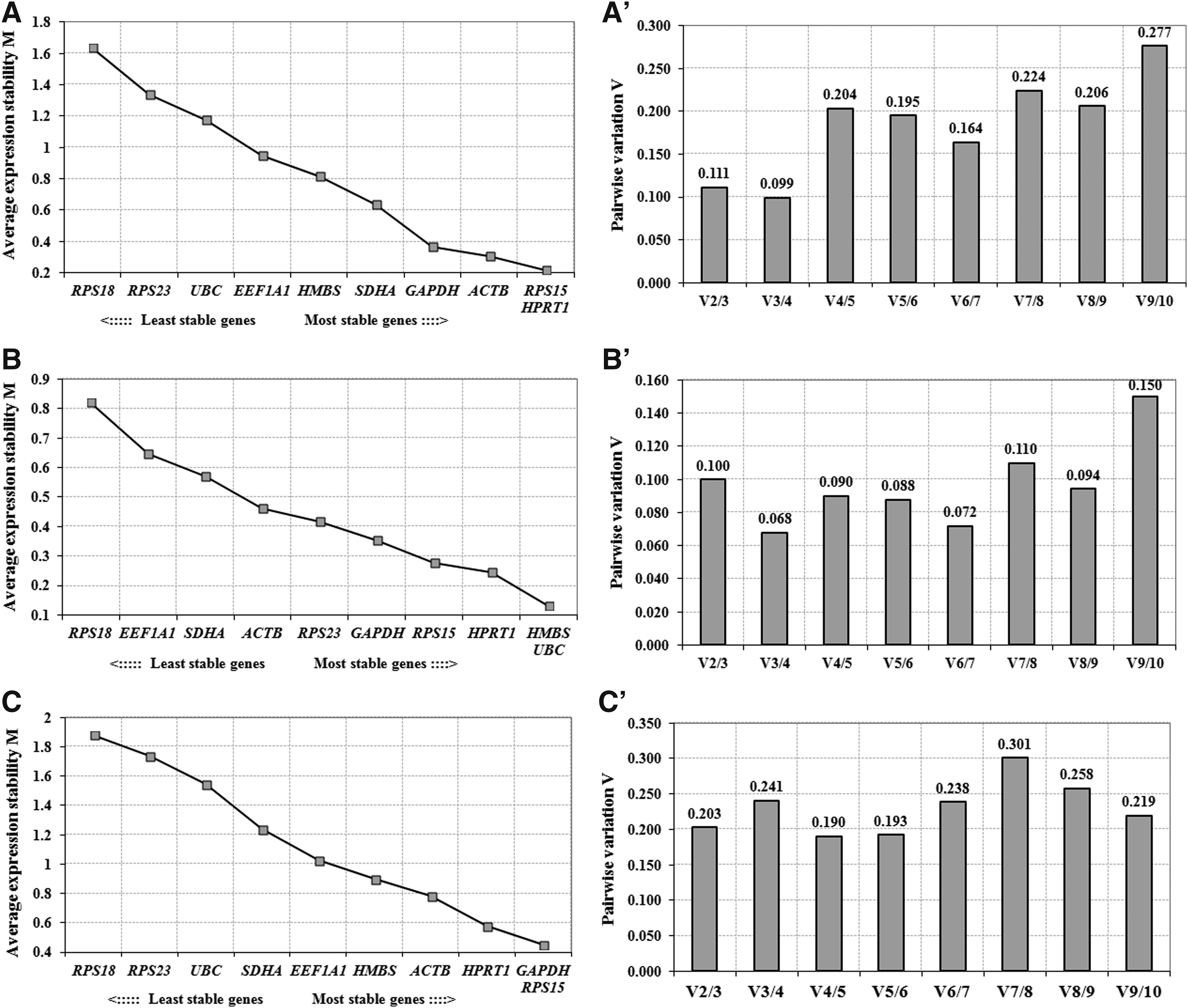
Average expression stability measure (M) for reference genes in
Then, less stable genes were sequentially added to these combinations of two most stably expressed genes, that is, combination of V2/3, V3/4, V4/5, V5/6, and so on. In case of cloned blastocysts, addition of third gene to the best two gave the V2/3 value of 0.111, therefore, the combination of three most stably expressed genes—RPS15, HPRT1, and ACTB was found to be the most suitable panel. For IVF blastocysts, the most suitable panel of three genes was HMBS, UBC, and HPRT1 with V2/3 value of 0.100.
Analysis of gene expression stability by NormFinder
The stability values of the genes determined by NormFinder are presented in Table 3. In cloned blastocysts, the best suitable reference genes were found to be in the following order based on their stability values RPS15 = HPRT1 > GAPDH > ACTB > UBC > EEF1A1 > SDHA > RPS23 > HMBS > RPS18. For IVF blastocysts, the genes were in the order HMBS = UBC > RPS23 > RPS15 > HPRT1 > SDHA > EEF1A1 > GAPDH > ACTB > RPS18; whereas, across both cloned and IVF blastocysts, it was HPRT1 > GAPDH > RPS15 > EEF1A1 > HMBS > ACTB > UBC > RPS18 > RPS23 > SDHA. In addition, the intra- and intergroup variations were also estimated by NormFinder. In the intragroup variation, the most stable genes were HPRT1, RPS15, and GAPDH for cloned, and HMBS, UBC, and RPS15 for IVF blastocysts (Fig. 3).

NormFinder-based intragroup variation analysis of candidate reference genes in cloned and IVF blastocysts. *Most stable genes.
IVF, in vitro fertilization.
In the intergroup variation between cloned and IVF blastocysts, the expression of HPRT1, EEF1A1, and HMBS was found to be least variable followed by that of GAPDH and RPS15 (Fig. 4). A comparison of geNorm and NormFinder analyses for cloned blastocysts, IVF blastocysts, and both the blastocyst types combined identified the same set of three genes to be the most stable with slight change in the ranking order (Table 4).
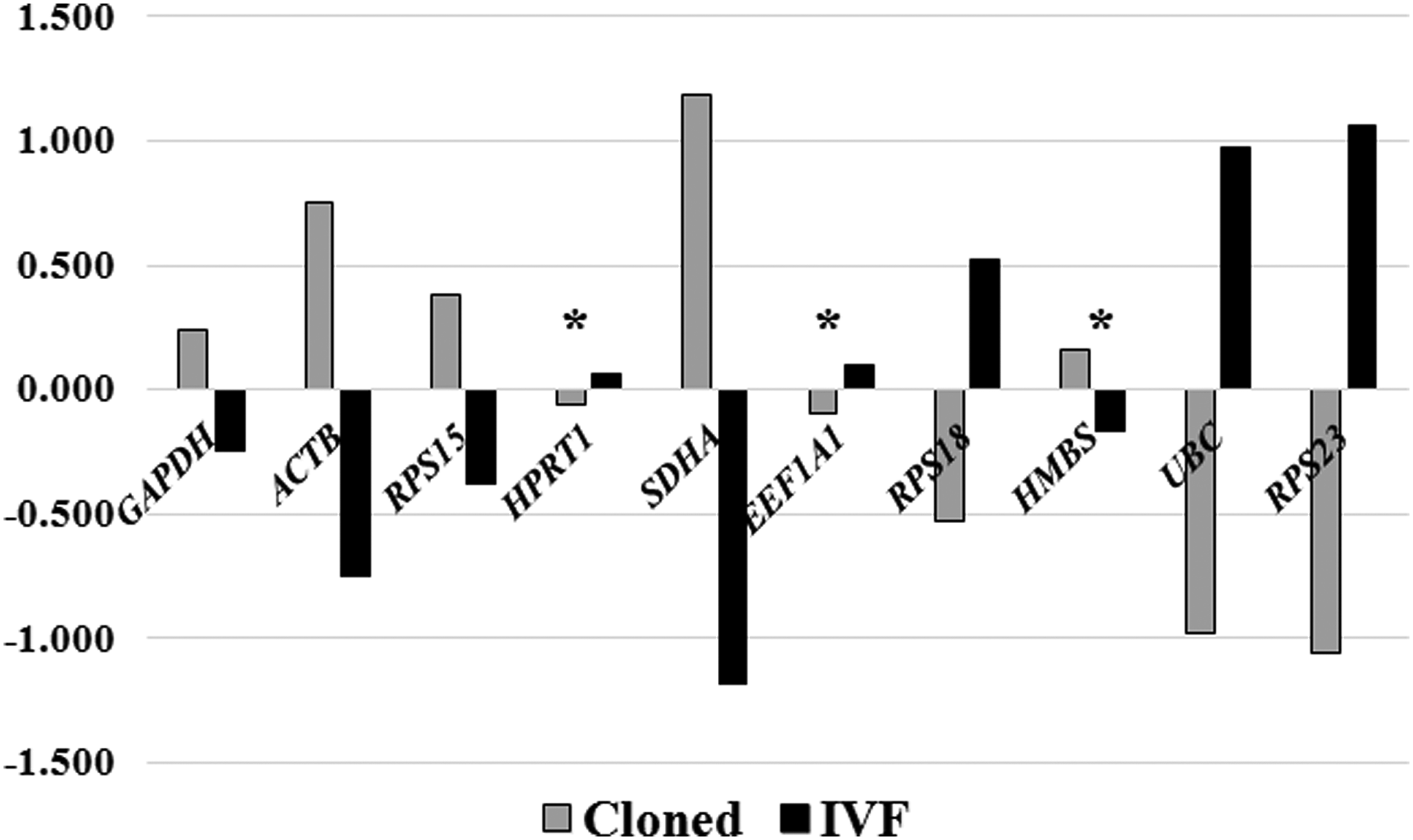
NormFinder-based intergroup variation analysis of candidate reference genes in cloned and IVF blastocysts. *Most stable genes.
Discussion
Using an ICG is considered to be the best normalization method for analysis of target gene expression data. Ideally, a perfect ICG should have a stable expression among various cells/tissues/developmental stages or experimental conditions (Thellin et al., 1999). If this condition is not met, the reference gene expression can be influenced by the experimental procedure, and detection of minute differences in the expression of genes under study becomes unfeasible (Bustin et al., 2009). To our information, there is only one report on the identification of suitable reference genes in buffalo embryos in which Kumar et al. (2012) reported RPS15, RPS18, and GAPDH to be the best reference genes, out of the six analyzed by geNorm, across oocytes and different embryonic stages. But identification of a highly stable gene, which is valid for oocytes and embryos of all preimplantation development stages, is confounded by the fact that the gene expression levels exhibit significant changes during embryonic development.
Expression of various reference genes has been found to remain constant or decrease slightly from mature oocytes to 6- to 8-cell or morula-stage bovine embryos, but is greatly increased in blastocyst-stage embryos (Bilodeau-Goeseels and Schultz, 1997). In another study in bovine cloned and IVF embryos, the RNA content was found to be similar in oocytes to 16-cell stage embryos, but there was a significant increase at morula and blastocyst stages (Ross et al., 2010). Also, none of the reference genes analyzed (GAPDH, PPIA, ACTB, RPL15, GUSB, and H2A.2) maintained stable expression levels across all the preimplantation stages of development. Therefore, it is essential to analyze individual embryonic stages for selection of best reference genes.
To identify the most stable reference gene and to find out the optimum number of reference genes, which should be used for mRNA expression studies, we examined the expression level of 10 commonly used ICGs, belonging to different functional classes, in blastocyst-stage cloned and IVF buffalo embryos by real-time qPCR and evaluated their stability using two different bioinformatics tools namely, geNorm and NormFinder. geNorm ranks reference genes according to the similarity of their expression profile by a pairwise comparison and uses their geometric means as a normalization factor, assuming that gene pairs showing stable expression patterns relative to each other are appropriate control genes (Vandesompele et al., 2002). NormFinder is a model-based approach that takes into consideration not just the overall expression variation, but also systematic variation across sample subgroups and top ranks the candidates with minimal estimated intra- and intergroup variations (Andersen et al., 2004).
Based on the lowest M value, determined by geNorm analysis, RPS15 and HPRT1 were found to be the most stable genes for cloned blastocysts, whereas HMBS and UBC for IVF blastocysts. In a similar study in cattle, among the 10 candidate genes examined, ACTB and SF3A1 were found to be the most stable genes in cloned blastocysts, and HMBS and SF3A1 for IVF blastocysts (Luchsinger et al., 2013). In another study, UBC, ACTB, RPL32, and GAPDH were found to be the most stable genes in equine IVF blastocysts (Smits et al., 2009). Contrastingly, we found RPS18 to be the least stable gene for both cloned and IVF blastocysts; whereas, it has been reported to be one among the panel of three best suitable ICGs, that is, RPS15, RPS18, and GAPDH for buffalo oocytes and IVF embryos in a previous study (Kumar et al., 2012).
This discrepancy could be due to the use of several developmental stages of oocytes and embryos in the previous study; whereas, our study was confined to the blastocyst stage. Since the use of a single reference gene for normalization of gene expression data can lead to biased and erroneous results, Vandesompele et al. (2002) recommended that for appropriate normalization, at least three most stable reference genes should be used for calculation of an RT-PCR normalization factor (NF n , n = 3).
On this basis, the panel of three most suitable genes was found to be RPS15, HPRT1, and ACTB for cloned blastocysts, and HMBS, UBC, and HPRT1 for IVF blastocysts. These authors further recommended that after identification of the three most stable genes, stepwise inclusion of more genes should be done until the (n + 1)th gene has no significant contribution to the newly calculated normalization factor (NFn+1). A large variation (V value) indicates that the next added gene has a significant effect and should preferably be included for calculation of a reliable normalization factor. The pairwise variation (V) analysis was carried out by stepwise inclusion of more genes until the next added gene made no significant contribution to the normalization factor. Since addition of the fourth gene, that is, GAPDH for cloned and HPRT1 for IVF blastocysts resulted in very low changes in V3/4 values of 0.099 and 0.068, respectively, inclusion of these genes in the panel does not offer any advantage.
Based on the lowest M value, RPS15 and GAPDH were found to be the most stable genes when data for both the blastocyst types were analyzed together in comparison to RPS15 and HPRT1, which were found to be the most stable genes for cloned blastocysts, and HMBS and UBC for IVF blastocysts. Luchsinger et al. (2013) reported GAPDH and HMBS to be the most stable reference genes in bovine blastocysts produced by cloning, IVF, and intracytoplasmic sperm injection.
In our study, the panel of three most suitable genes was found to be RPS15, GAPDH, and HPRT1 for both the blastocyst types analyzed together. The V3/4 value increased substantially to 0.241 when data for both the blastocyst types were analyzed together compared with that of 0.099 and 0.068 for cloned and IVF blastocysts, respectively. This is probably because the set of three most suitable genes was different for IVF blastocysts (HMBS, UBC, and HPRT1) compared with cloned (RPS15, HPRT1, and ACTB), and both blastocyst types analyzed together (RPS15, GAPDH, and HPRT1). These results suggest that although there is an overlapping of the most suitable gene between cloned and IVF blastocyst-stage embryos in the form of HPRT1, ideally, the panel of reference genes selected for both types of embryos should be specific to the embryo type used.
Similar results were obtained following NormFinder analysis, although with minor differences in the ranking of genes (boldface in Table 4). The order of stability was RPS15>HPRT1>GAPDH for cloned blastocysts, HMBS>UBC>RPS23 for IVF blastocysts, and HPRT1>GAPDH>RPS15 for cloned and IVF blastocysts analyzed together. Following intragroup variation analysis by NormFinder, RPS18 was found to be the most variable gene in both cloned and IVF blastocysts, whereas intergroup variation analysis revealed HPRT1, EEF1A1, and HMBS to be the least variable genes.
There could be several possible reasons behind the differences in the panel of most stable genes identified for cloned and IVF embryos. The epigenetic reprogramming, which involves 10,000 to 12,000 genes that drive embryonic and fetal development has been established to be incomplete or abnormal in cloned embryos leading to aberrant expression of several genes compared with their IVF counterparts (Niemann, 2016).
In addition to the effect of IVC conditions (Niemann and Wrenzycki, 2000) and procedures used for production of cloned and IVF embryos (Ross et al., 2010), which may have affected gene expression, the gender of the embryos may also have contributed to these differences since all the cloned embryos were of the same sex; whereas, the IVF embryos were a mixed population of male and female embryos. The differences in the stability of expression level of candidate genes between cloned and IVF embryos contributed to the large variation in M value range, which was observed when the two embryo types were analyzed together.
However, almost all the candidate genes examined in the present study showed high expression stability in each embryo type, as indicated by their low M values, which was <1.5 for most of the genes, which led to their selection as reference genes in previous studies in cattle and buffalo embryos (Goossens et al., 2005; Kumar et al., 2012; Luchsinger et al., 2013). Our results show that differences in gene expression stability were still present among these genes and between different embryo production methods, confirming that a careful selection of the best candidate genes is strongly recommended for each experimental condition.
In conclusion, the results of this study suggest that despite overlapping in the panel of three most stable genes between cloned and IVF blastocyst-stage embryos, the panel of ICGs selected for both types of embryos should be different. Our results show that the panel of the most suitable ICGs for qPCR data normalization of buffalo blastocyst-stage embryos was RPS15, HPRT1, and ACTB for cloned blastocysts, HMBS, UBC, and HPRT1 for IVF blastocysts, and GAPDH, RPS15, and HPRT1 for both the embryo types analyzed together. RPS18 is not a stable gene for use in qPCR data normalization for buffalo cloned and IVF blastocysts.
Footnotes
Acknowledgments
T.J.S. and S.V.L. were supported by fellowships from the Indian Council of Medical Research and University Grants Commission (India), respectively.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.