
Abstract
Abstract
This study was aimed to investigate the role of Wnt/β-catenin signaling pathway in the differentiation from periodontal ligament stem cells (PDLSCs) to Schwann cells (SCs) and the possible mechanisms. FzB was applied to inhibit the Wnt/β-catenin signaling pathway of differentiated PDLSCs (dPDLSCs), and then immunofluorescence, polymerase chain reaction (PCR), and Western blotting analysis were performed to detect SC marker genes and proteins such as S100, glial fibrillary acidic protein (GFAP), and P75NTR. Results showed that when the Wnt/β-catenin signaling pathway was inhibited, the expression of S100, GFAP, and P75NTR protein significantly decreased in dPDLSCs, p < 0.05, whereas PCR results showed that expression of SC myelinogenesis-related genes krox-20, Oct-6, P0, and PMP-22 was significantly downregulated at the same time, p < 0.05. These results showed that Wnt/β-catenin signaling pathway participated in the differentiation from PDLSCs to SCs, and inhibiting it could inhibit the differentiation process.
Introduction
S
However, some different results show that the Wnt pathway inhibits the differentiation from embryonic stem cells to nerve stem cells (Sato et al., 2004), whereas the high expression of the Wnt1 pathway depresses myelogenesis; meanwhile depression of the Wnt pathway with Sfrp2 precipitates nervous system phylogeny (Schulte et al., 2005). Recent researches also prove the vital role of the Wnt pathway in the orientation differentiation from periodontal ligament stem cells (PDLSCs) to osteoblast (Nagatomo et al., 2006).
Our previous researches have proven that PDLSCs could differentiate into SCs like cells, but the mechanism was still unclear (Li et al., 2010; Xiaojie et al., 2009). Owing to the important role of Wnt/β-catenin in neural development, we aimed to investigate whether Wnt/β-catenin could regulate the differentiation from PDLSCs to SCs. After applying special inhibitor of Wnt/β-catenin during the differentiation, we detected the protein and gene expression of SC markers, S100, GFAP, and P75, and SC myelinogenesis-related genes, krox-20, Oct-6, P0, and PMP-22, to investigate whether Wnt/β-catenin played an important role in this differentiation.
To our best knowledge, up to now, no study focused on Wnt signaling pathway in process of transdifferentiation from PDLSCs to
Materials and Methods
Culture and identification of PDLSCs
Twenty-four teeth from 8-month-old beagle dogs (total 4) were collected under aseptic conditions and placed in PBS solution with 1% (v/v) penicillin/streptomycin for 5 minutes. Then the periodontal ligament was gently scraped from the middle of teeth root and dissociated with 0.25% trypsin (Sigma, United Kingdom) and 0.1% collagenase (Sigma) for 90 minutes at 37°C. The suspension was passed through a 75 μm filter to remove undissociated tissue, and then neutralized with α-MEM plus 20% (v/v) FBS (Hyclone), and centrifuged at 1000 g for 8 minutes. Then the cell pellet was resuspended in the mentioned medium including 1% (v/v) penicillin/streptomycin solution, and cultured in an incubator at 37°C with 5% carbon dioxide. The medium was first changed 7 days later and then changed regularly at 3-day intervals. The cells were passaged with trypsin/EDTA (Sigma) when needed. Single-colony selection was used to isolate PDLSCs (Li et al., 2010; Seo et al., 2004). Then PDLSCs were proliferated and identified as previously described (Li et al., 2010).
This study was approved by the Ethic Committee of Department of Dentistry, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, China.
Induction of PDLSCs to differentiate into SCs
Three groups were divided in our experiment: the first was the differentiated PDLSCs (dPDLSCs), the second was the dPDLSCs with FzB as a special inhibitor of Wnt/β-catenin (dPDLSCs+FzB), and the third group was undifferentiated PDLSCs (uPDLSCs). The protocol to induce PDLSCs to differentiate into SCs was as formerly described (Li et al., 2010): the growth medium was removed from subconfluent cultures and replaced with medium supplemented with 1 mM β-mercaptoethanol (Sigma-Aldrich, United Kingdom) for 1 day. Then, the cells were cultured in α-MEM supplemented with 2% DMSO for 5 hours. Subsequently the medium was replaced with a differentiation medium, including 10 ng/mL PDGF, bFGF, and brain-derived neurotrophic factor (BDNF) (PeproTech Ltd., United Kingdom) with 14 μM forskolin (Sigma). Fresh medium was added every 3 days and the cells were induced for 2 weeks. In group 2, 30 μM FzB was added in the differentiation medium.
Fluorescence microscope observation
Cells of three groups were innoculated on glass cover slips with a density of 2 × 105 cells/well for 1 day. Then antibodies of S100, GFAP, and P75 (Abcam) were applied to perform immunofluorescence detection as previously described (Li et al., 2010).
Western blotting analysis
Total protein was extracted with a commercially available protein assay kit (Bio-Rad, United Kingdom). After detection of protein concentrations, 20 μg of the protein was mixed with an equal amount of 2 × sodium dodecyl sulfate loading buffer for Western blotting analysis as previously described (Li et al., 2010).
RNA extraction and RT polymerase chain reaction
Total RNA was extracted with the TRIZOL reagent according to manufacturer's suggestions (Invitrogen, Carlsbad, CA). The freshly isolated RNA was measured by a spectrophotometer at 260/280 nm, and then detected by electrophoresis on a 3% (w/v) agarose gel. Total RNA (500 ng) was reverse transcribed into cDNA with a PrimeScriptTM RT Reagent Kit (Perfect Real Time, Takara, Japan) according to the manufacturer's advices. Primers were designed with Primer 5.0 analysis software. Polymerase chain reaction (PCR) solution was amplified by the ABI 7300 Real-Time PCR System (Applied Biosystems). The program adopted was 94°C for 5 minutes, 38 cycles of 94°C for 30 seconds, and 72°C for 30 seconds. The PCR products were analyzed using agarose gel electrophoresis (BIO-RAD). Relative quantification of RT-qPCR product was performed using the comparative 2−ΔΔCT method (Livak and Schmittgen, 2001). Primers used in this study are as follows: Krox-20, F: 5′-AGATACCATCCCAGGCTCAGT-3′, R: 5′-CTCTCCGGTCATGTCAATGTT-3′; oct-6, F: 5′-AGGTCCTGTTGGAGATGATATGTT-3′, R: 5′-TTGGGAAATGAATTGTCAAGAAA-3′; p0, F: 5′-GAATTCATGGCTCCCGGGGCTCCCT-3′, R: 5′-AAGCTTCTATTTCTTATCCTTGCGAGACTCC-3′; PMP-22, F: 5′-CCCAACTCCCAGCCACCATG-3′, R: 5′-TCATTCGCGTTTCCGCAGGATC-3′.
Statistical analysis
All experiments were conducted in triplicate. The measurement data were expressed by mean ± SD. Independent continuous variables were compared using the Student t-test. It was considered to be statistically significant when p-value was less than 0.05. All calculations were made using SPSS 18.0.
Result
Inhibition of Wnt/β-catenin signaling pathway could inhibit SC marker proteins
Two weeks after induction, Western blotting analysis was performed to detect Wnt/β-catenin pathway-related proteins such as β-catenin, Dsh, GSK-3β, c-myc, and CyclinD1. Results are shown in Figure 1. uPDLSCs were used as control group. When FzB was used, the expression levels of Wnt/β-catenin pathway-related proteins were significantly inhibited, p < 0.05, indicating the successful interference for Wnt signaling pathway. As shown in Figure 2, in the dPDLSCs group, S100, GFAP, and P75NTR were detected as normal, whereas in the dPD+FzB group, the mentioned protein expression levels were depressed obviously, p < 0.05, suggesting that the differentiation from PDLSCs to SCs was disturbed when the Wnt/β-catenin signaling pathway was inhibited by FzB, so resulting in SC marker proteins reduction.
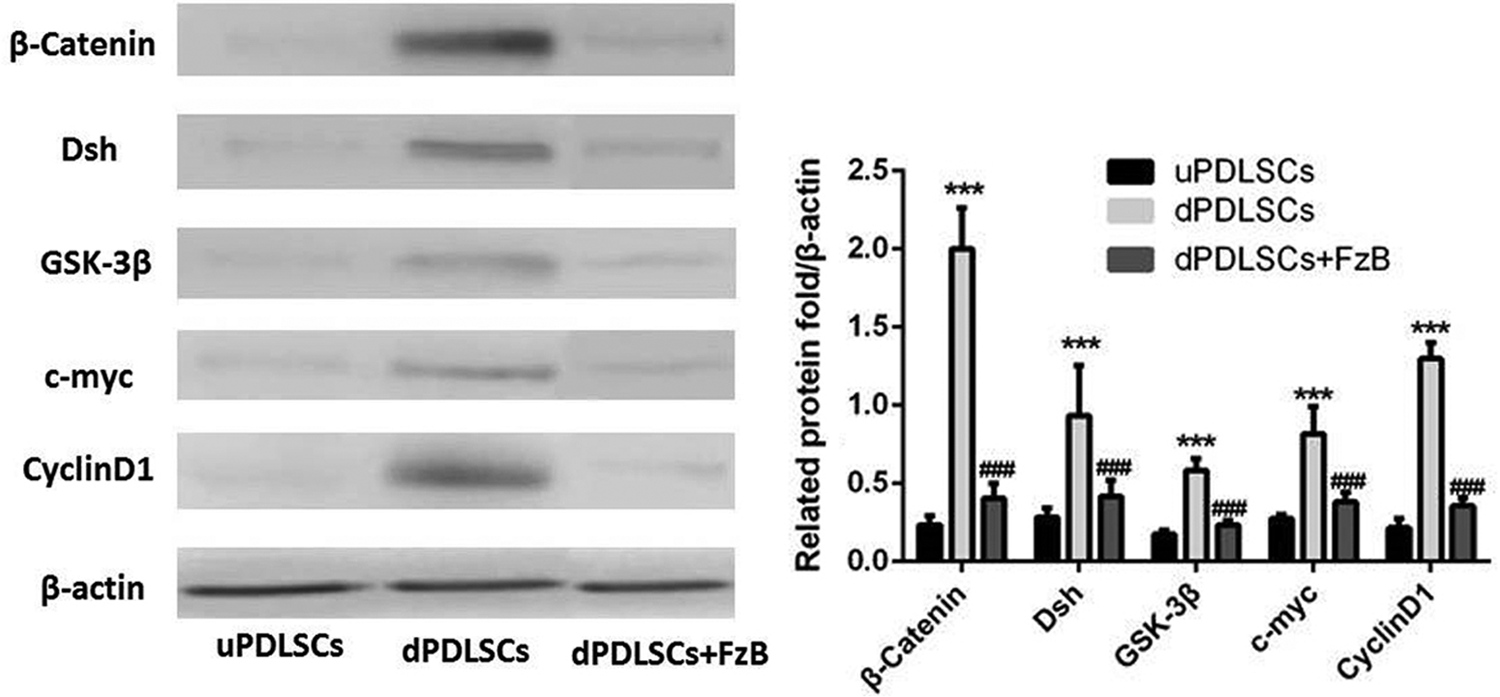
In uPDLSCs and dPDLSCs+FzB groups, β-catenin, Dsh, GSK-3β, c-myc, and CyclinD1 protein expression levels were negative, whereas in dPDLSCs group, those protein expression levels were obvious. ***p < 0.05, compared with the uPDLSCs; ###p < 0.05, compared with the dPDLSCs. dPDLSC, differentiated periodontal ligament stem cells; uPDLSCs, undifferentiated PDLSCs.
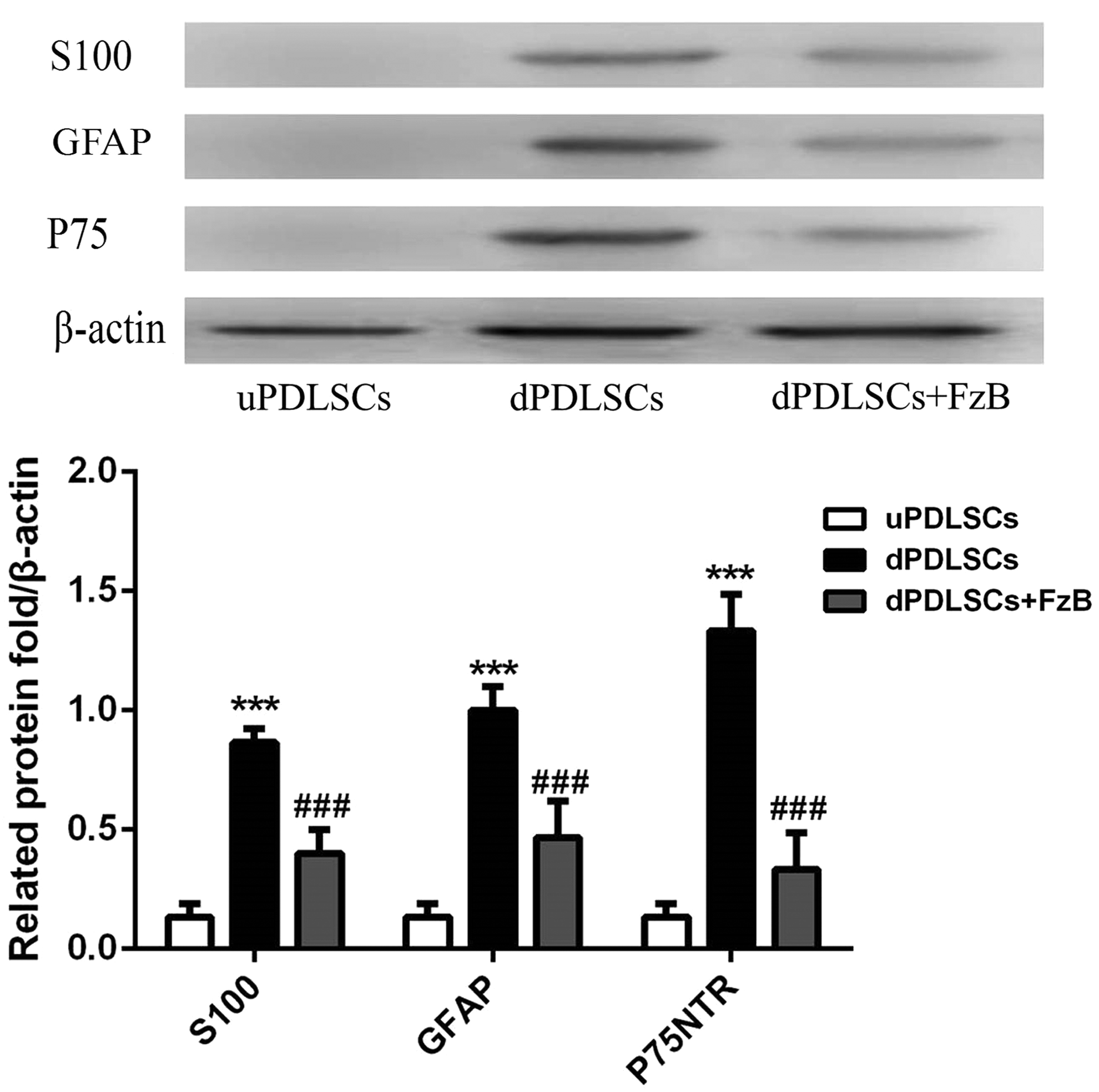
In dPDLSCs group, S100, GFAP, and P75NTR were detected as normal, whereas in dPD+FzB group, the mentioned protein expression levels were depressed obviously. ***p < 0.05, compared with the uPDLSCs; ###p < 0.05, compared with the dPDLSCs. GFAP, .
Further immunofluorescence study also demonstrated similar results. As shown in Figure 3, dPDLSCs group appeared SC shape obviously after 2 weeks induction, and cell bodies stretched out to form synapse and contacted with each other with green fluorescence in blue DAPI labeled cells cytoplasm, indicating strong positive for S100 antigen (A); dPD+FzB group also partly appeared SC shape after 2 weeks induction, and some blue DAPI labeled cells negative for S100 antigen (B); dPDLSCs group showed strong green fluorescence in blue DAPI labeled cells cytoplasm, indicating strong positive for GFAP antigen (C); while in dPD+FzB group, most DAPI labeled cells showed no green fluorescence in cytoplasm, indicating negative or weak positive for GFAP antigen (D); dPDLSCs group showed strong green fluorescence on DAPI labeled cells membrane, indicating strong positive for P75NTR antigen (E); while in dPD+FzB group only a few DAPI labeled cells showed weak green fluorescence on membrane, indicating weak positive or negative for P75NTR antigen (F).
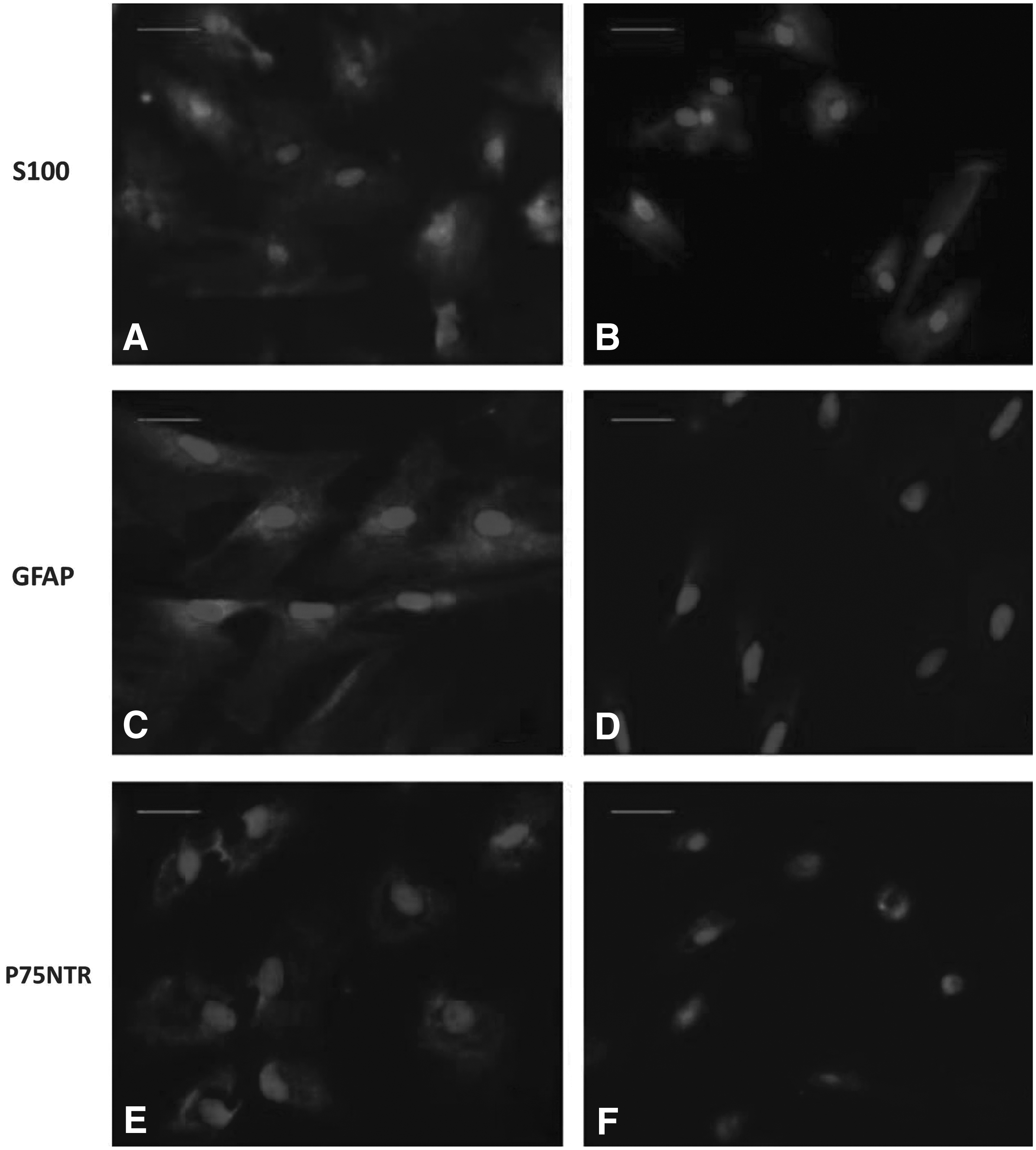
Immunofluorescence detection for S100, GFAP, and P75NTR protein of dPDLSCs and dPD+FzB group.
Inhibition of Wnt/β-catenin signaling pathway could inhibit SC myelinogenesis-related genes
To study effects of Wnt signaling pathway on expression of SC myelinogenesis-related genes krox-20, Oct-6, P0, and PMP-22, PCR method was used to determine mRNA level of the mentioned factors. As shown in Figure 4, expression of krox-20, Oct-6, P0, and PMP-22 was downregulated in the dPD+FzB group, p < 0.05, suggesting that Wnt/β-catenin signaling pathway inhibition disturbed PDLSCs differentiation into SCs. Meanwhile, expression levels of SC myelin sheath-related genes were also downregulated and finally led to the failure of SC myelinization.
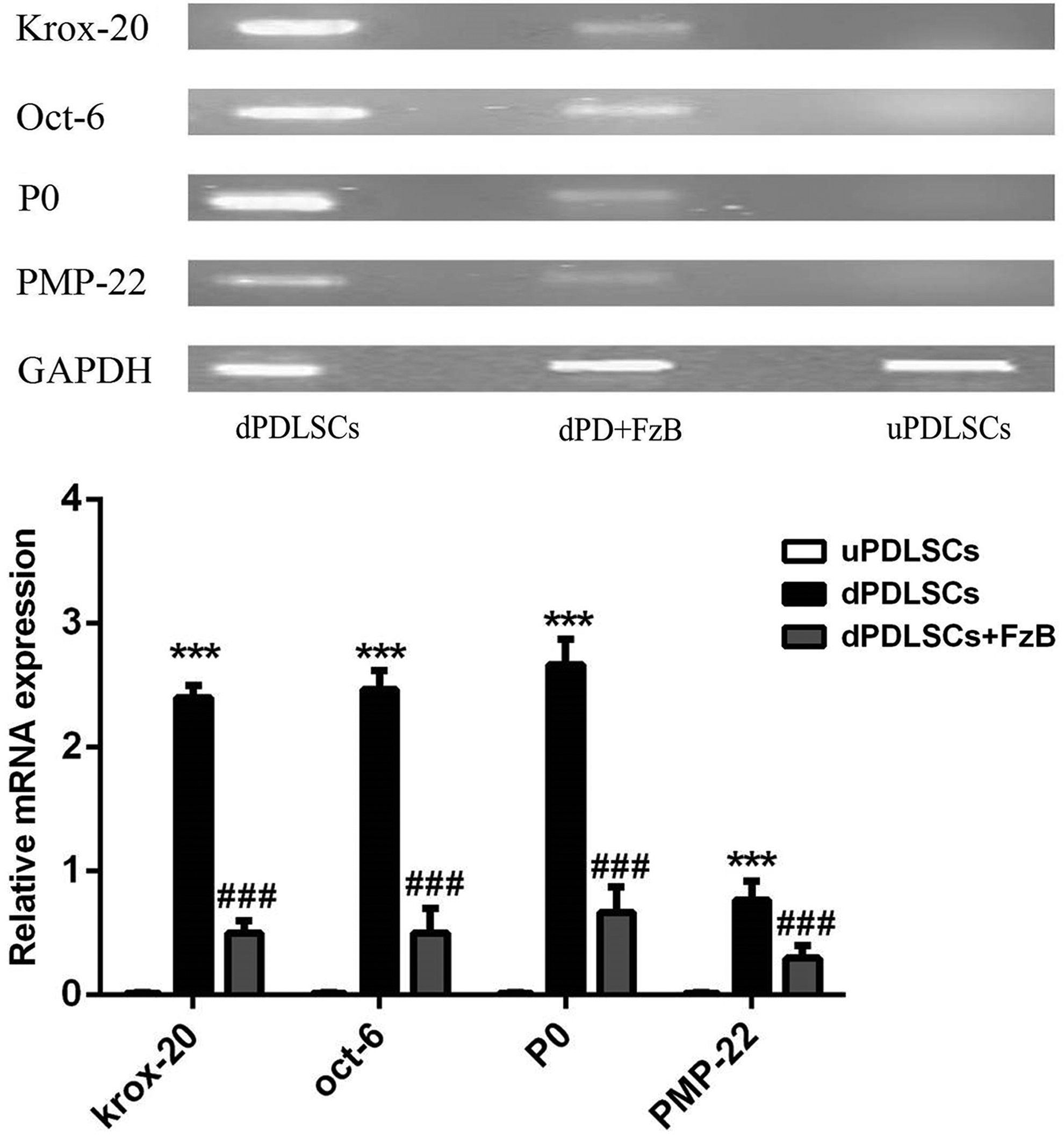
PCR agarose gel electrophoresis results. Krox-20, oct-6, p0, and PMP-22 gene expression was detected in dPDLSCs, dPD+U0126, and uPDLSCs groups. GAPDH as house-keeping gene. ***p < 0.05, compared with the uPDLSCs; ###p < 0.05, compared with the dPDLSCs. PCR, polymerase chain reaction.
Discussion
Neurotrophin plays an important role in SCs development and injured peripheral nerve regeneration, many extracell factors regulate nerve development and regenerate by activating Wnt signaling pathway. NGF is one of the earliest neurotrophic factors discovered by researchers, which is very important in promoting SC myelinization and axon growth, also in accelerating SCs maturation (Hempstead and Salzer, 2002); BDNF, with a molecular weight of 13 kD, is one kind of neurotrophic factor existing in homodimer, which is also vital in injured peripheral nerve repair, neuron survival, axon regeneration, and myelinization (Levi-Montalcini et al., 1996). Neurotrophic factor produces its marked effect mainly by two kinds of receptors, one is tyrosine-related kinase (Trk), a high affinity receptor, and the other is P75, a low affinity receptor. Trk is a transmembrane receptor with three subtypes such as TrkA, TrkB, and TrkC, which binds to neurotrophic factors selectively. NGF binds to TrkA, and BDNF binds to TrkB, then form dimeride, promote protein tyrosine kinase phosphorylation, activate downstream signaling molecule, and finally promote neuron growth, synapse shaping, and nerve repair. Trk signaling pathway participates in SC development by activating PLC and PI3K/Akt, etc. (Harrisingh et al., 2004).
However, there is another signaling pathway more important in SC development; it is Wnt signaling pathway that is a complicated regulation network, including three branches (Huelsken and Behrens, 2002): (1) classic Wnt pathway, activating genetic transcription by β-catenin; (2) Wnt/PCP pathway, regulating cell framework rearrangement by activating c-Jun N terminal kinase (JNK) with G proteinum; and (3) Wnt/Ca2+ pathway, affecting cytoadherence and gene expression through intracellular Ca2+ release. There is a dynamic degradation complex of classic Wnt pathway in cytoplasm, which is composed of β-catenin, Axin, APC, GSK 3β, CK1, etc.
The activation or depression of classic Wnt pathway is decided by the concentration of β-catenin in cytoplasm. The activation of classic Wnt pathway begins from the combination of secreting Wnt glycoprotein and Frizzled receptor. Frizzled is a kind of receptor protein on the surface of plasmalemma with seven transmembrane structures, combining with LRP5/6 to form a specific binding target of ligand Wnt, then activating protein Dsh with a PDZ structural domain, releasing signal to depress the degradation activity of β-catenin by GSK 3β. During the process, Dsh activates guanine nucleotide binding protein (GBP), then GBP liberates GSK 3β from degradation complex, inducing β-catenin to accumulate in cytoplasm, sequentially entering into cell nucleus to activate transcription factor LEF/TCF, switching on the transcription of target gene (Mulholland et al., 2005).
In this study, we combined neurotrophic factors, such as NGF and BDNF, to induce PDLSCs to differentiate into SCs like cells. Our results showed that dPDLSCs could express SC special markers such as S100, GFAP, and P75NTR protein. Although after inhibition of Wnt signaling pathway by FzB, expression of S100, GFAP, and P75NTR was significantly decreased, suggesting that the Wnt pathway probably participated in the differentiation process.
PCR results showed that expression of SC myelinization-related genes, Oct-6, krox-20, P0, and PMP-22 was downregulated in the dPD+FzB group, suggesting that the Wnt pathway probably participated in myelinization during the differentiation from PDLSCs to SCs. We also found that although the Wnt pathway was inhibited, some PDLSCs still differentiated into SCs like cells. This result might be due to other signaling pathways participating in the differentiation, such as NRG-1, ERK1/2, and Notch pathways. So in future studies, we intend to use gene knockout animal models to further investigate the mechanisms, meanwhile other signaling pathways cross-talking with Wnt are under consideration in further investigation.
Conclusion
We applied FzB to depress Wnt pathway during the induced differentiation from PDLSCs to SCs. Results showed that expression of SC-related protein, S100, GFAP, and P75NTR, was significantly reduced, and expression of SC myelinization-related genes, Oct-6, krox-20, P0, and PMP-22, was significantly downregulated. Our results suggested that the Wnt pathway participated in the differentiation from PDLSCs to SCs, and to depress the Wnt pathway could disturb the differentiation partly.
Footnotes
Acknowledgment
The study was approved by the Zhejiang Province Natural Science Foundation (491020+N21129).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.