
Abstract
Abstract
Induced pluripotent stem cells (iPSCs) from somatic cells can be reprogrammed to provide an unlimited cell resource showing great potential in disease modeling and regenerative medicine. However, the traditional method for reprogramming cells into iPSCs using genome-integrating retro- or lenti-viruses remain an obstacle for its application in clinical settings. We tried the possibility to generate pre-iPSCs from human adipose-derived stem cells (ADSCs) by nongenetic reprogramming using recombinant cell-penetrating proteins OCT4/KLF4/SOX2 (PTD-OKS) and the cocktail of small molecules (VCFZ). Our experimental results demonstrated that PTD-OKS in combination with VCFZ (VCFZ+OKS) could significantly enhance the stemness of ADSCs and easily get pre-iPSCs after 25 days treatments. The pre-iPSCs showed similar morphology to iPSCs, which were positive for alkaline phosphatase staining. Furthermore, RT-polymerase chain reaction analysis showed that VCFZ+OKS could significantly upregulate the expression of OCT4, KLF4, SOX2, and NANOG gene after 25 days treatment. And immunofluorescence staining also showed that the protein makers of pluripotent stem cell were positively expressed in VCFZ+OKS treated group. Our data suggest that nongenetic-mediated reprogramming from ADSCs may be a promising stem cell sources for cell therapy in the near future.
Introduction
I
For example, the transcription factor c-Myc, Klf4, and a cartilage factor Sox9 were used to induce human epidermal fibroblasts into chondrocytes. After transplantation, the chondrocytes were found to be formed in nude mice tumor (Baer and Geiger, 2012). Interestingly, the reprogramming process often results in the production of cells with a stable intermediate “pre-iPSCs” state. Pre-iPSCs exhibit morphology identical to that of iPSCs and express endogenous pluripotency markers (Huang et al., 2015). Thus, pre-iPSCs and iPSCs are not only the starting, intermediate, and final stages of reprogramming, but also represent useful tools for delineating the molecular changes associated with reprogramming and changes in cell fate.
To circumvent the problems associated with traditional reprogramming methods, different approaches such as the use of small molecules and chemicals are being explored (Dai et al., 2014; Desponts and Ding, 2010; Hou et al., 2013; Li et al., 2011; Lu et al., 2013; Lyssiotis et al., 2009). It was first reported the use of histone deacetylase inhibitor, Valproic Acid (VPA) to improve reprogramming efficiencies (Huangfu et al., 2008). Since then, the usage of small molecules and chemicals to facilitate reprogramming through induction or by replacing one or two reprogramming factors have been widely explored (Lee et al., 2012; Park et al., 2015; Zhou et al., 2016). In 2011, Moon shows the possibility of reprogramming mouse fibroblasts with the combination of OCT4 and small molecule purmorphamine was reported (Moon et al., 2011). Moreover, Hou et al. (2013) show that pluripotent stem cells can be generated from mouse somatic cells at a frequency up to 0.2% using a combination of seven small-molecule compounds.
In recent times, protein-based delivery has gained attention, due to the discoveries that iPSCs induction and epigenetic reprogramming could be carried out without the use of traditional viral vectors (Singhal et al., 2015; Thier et al., 2010; Zhang et al., 2012; Zhou et al., 2009). Zhou et al. (2009) reported the generation of mouse iPSCs using four rounds of protein transduction with reprogramming factors, such as OCT4, SOX2, KLF4, and c-Myc, which were tagged with polyarginine in the presence of VPA. By using cell permeable protein transduction approach, a protein could be delivered easily into cells without the use of a permeabilization agent such as streptolysin-O (SLO). This delivery method ensures that no genomic integration takes place and these techniques provide practical reprogramming alternatives in the clinical setting. Furthermore, protein reprogramming allows better control of timing and delivery of reprogramming factors in a concentration-dependent manner; however, the disadvantage of a relatively low efficiency of reprogramming with these methods must be addressed.
An important consideration for iPS technology is determining an appropriate starting somatic cell type. A majority of iPS studies use fibroblast cells, most likely due to their ease of derivation and extension. Various other starting somatic cell populations have been used for iPS induction in the human, including dental pulp cells (Kawano et al., 2017), urine-derived cells (Wang et al., 2016), hematopoietic cells (Eminli et al., 2009), and adipose-derived stem cell (ADSC) (Zapata-Linares et al., 2016). An important observation from these studies is that the somatic cell type chosen had a significant influence on the efficiency of iPS generation and level of reprogramming.
Pollyanna et al. also reported that ADSC as a readily accessible somatic cell type that is an ideal candidate for high-efficiency establishment of iPS cell lines using retroviral transduction of the Yamanaka factors (Goh et al., 2013). In this study, we investigated the effects of cocktail of chemical compounds (VCFZ) and recombinant cell-penetrating reprogramming proteins OCT4/KLF4/SOX2 (PTD-OKS) in reprogramming human ADSCs (hADSCs). The goal was to understand whether the combination of PTD-OKS proteins and small molecules VCFZ can act synergistically and be used as a tool for nongenetic reprogramming of ADSCs.
Ethics statement
Six women with a mean age of 35.1 ± 6.5 years were selected for the study after written informed consent was obtained. The institutional ethical review board of the First Affiliated Hospital of Jinan University approved the protocols. A tumescent solution consisting of a mixture of 0.9 NaCl, 0.1% lidocaine, and 1:100,000 epinephrine was injected using a 50-mL syringe into the fat donor sites of each patient's abdomen. A 2.5-mm-diameter cannula and 20-mL-syringes were used to harvest 200 mL adipose tissue from the abdomen of each patient. All the data used in this study were anonymized.
Materials and Methods
Materials
Culture reagents were from Gibco (Grand Island, NY). Unless otherwise stated, all the other reagents were from Sigma (St. Louis, MO). VPA was from Suju (Guangzhou, China). CHIR99021 (C), Forskolin (F), and DZNeP (Z) were from MedChem Express (NJ). Cell Counting Kit-8 (CCK-8) was from Dojindo (Kyushu, Japan). Klf4 mouse polyclonal antibody was from GeneTex (Alton). Oct4, Sox2, and Nanog rabbit monoclonal antibody were purchased from Cell Signaling Technology (Boston). Goat anti-mouse IgG and goat anti-rabbit IgG were from Santa Cruz Biotechnology (Santa Cruz, CA). EZgeneTM Tissue RNA Miniprep Kit was from Biomiga (San Diego, CA). ReverTra Ace qPCR RT Kit, Blend Taq® and Blend Taq-Plus were from Toyobo (Osaka, Japan). Primers were synthetized by BGI (Shenzhen, China), PTD-OKS was prepared as our previous study (Dai et al., 2014).
Isolation and primary culture of hADSCs
Subcutaneous adipose tissue was obtained from patients undergoing elective abdominal dermolipectomy. No objection certificates were obtained according to bioethics law. Briefly, adipose tissue was repeatedly washed with phosphate-buffered saline (PBS) until blood was completely removed from the tissue, and then incubated with equal volume of Dulbecco's modified Eagle's medium (DMEM) containing 0.01% type I collagenase at 37°C for 1 hour in a shaking incubator at 110 rpm. The suspension was filtered through a 100-mesh nylon membrane and then centrifuged at 300 × g for 10 minutes. The remaining tissues were then rinsed in the culture medium composed of DMEM, followed by centrifugation at 300 × g, 5 minutes, and suspended at a concentration of 1 × 104 cells/mL in a conventional medium (low-glucose DMEM supplemented with 10% fetal bovine serum (FBS), 1%penicillin-streptomycin). The cells were seeded into a 25 cm2 plastic culture flask, fed with 4 mL of media, and incubated at 37°C in a 5% CO2 incubator. The culture medium was changed every second day.
Osteogenic and adipogenic differentiation of hADSCs
ADSCs were plated at 1 × 104 cells/mL and cultured in conventional medium for 24 hours. Afterward, the medium was then changed to an adipogenic induction medium: DMEM/F12, 10% FBS, 1 μM insulin, 200 μM indometacin, 1 μM dexamethasone, 0.5 mM isobutyl-methylxanthine, and 1% antibiotic-antimycotic. Three days later, the medium was refreshed. Adipogenic differentiation was confirmed by staining of lipids by Oil Red O. Cells were plated at 1 × 104 cells/mL and cultured in conventional medium for 24 hours. Afterward, the medium was then changed to an osteogenic induction medium: DMEM/F12, 10% FBS, 0.1 μM dexamethasone, 50 mg/L 2-phosphate ascorbic acid, 10 mM β-glycerophosphate, and 1% antibiotic-antimycotic. The medium was refreshed 3 days a week for 2 weeks. Osteogenic differentiation was confirmed by staining calcium-rich deposits with Alizarin red.
Cell proliferation assay
A CCK-8 was employed to identify the effect of PTD-OCT4, PTD-KLF4, and PTD-SOX2 on the proliferation of ADSCs. Briefly, cells were seeded in 96-well flat-bottomed plate at 1 × 104 cells/well and incubated at 37°C for 24 hours. Subsequently, the cells were treated with increasing concentrations of PTD-OKS (at 30, 40, 50, and 60 μg/mL) for 72 hours. Ten microliters dye added to each well, cells were incubated at 37°C for 2 hours. The absorbance value at 450 nm was determined using the multimode reader. Six parallel experiments in each sample were used to assess the cell proliferation. Similar experiments were carried out to assess the proliferation rate of ADSCs treated with small molecular (CVFZ) and PTD-OKS. Briefly, ADSCs were seeded in 96-well flat-bottomed plate and treated with or without small molecular 3 μM CHIR99021, 1 μM VPA, 10 μM Forskolin, 0.5 μM DZNeP, and PTD-OKS (50 μg/mL) for 72 hours. Ten microliters dyes added to each well, cells were incubated at 37°C for 2 hours. The absorbance value of each well at 450 nm was measured using the Multimode Reader. Six different well per plate were used to assess the cell proliferation.
Generation of human pre-iPSCs
In the VCFZ+OKS group (Fig. 1A), 1 × 104 ADSCs were first incubated with conventional medium for 24 hours. After that, ADSCs were incubated with induction medium (DMEM/F12, 20% KSR, 0.5% ITS, 0.1 mM non-necessary amino acid, 1 mM sodium pyruvate, 2 mM
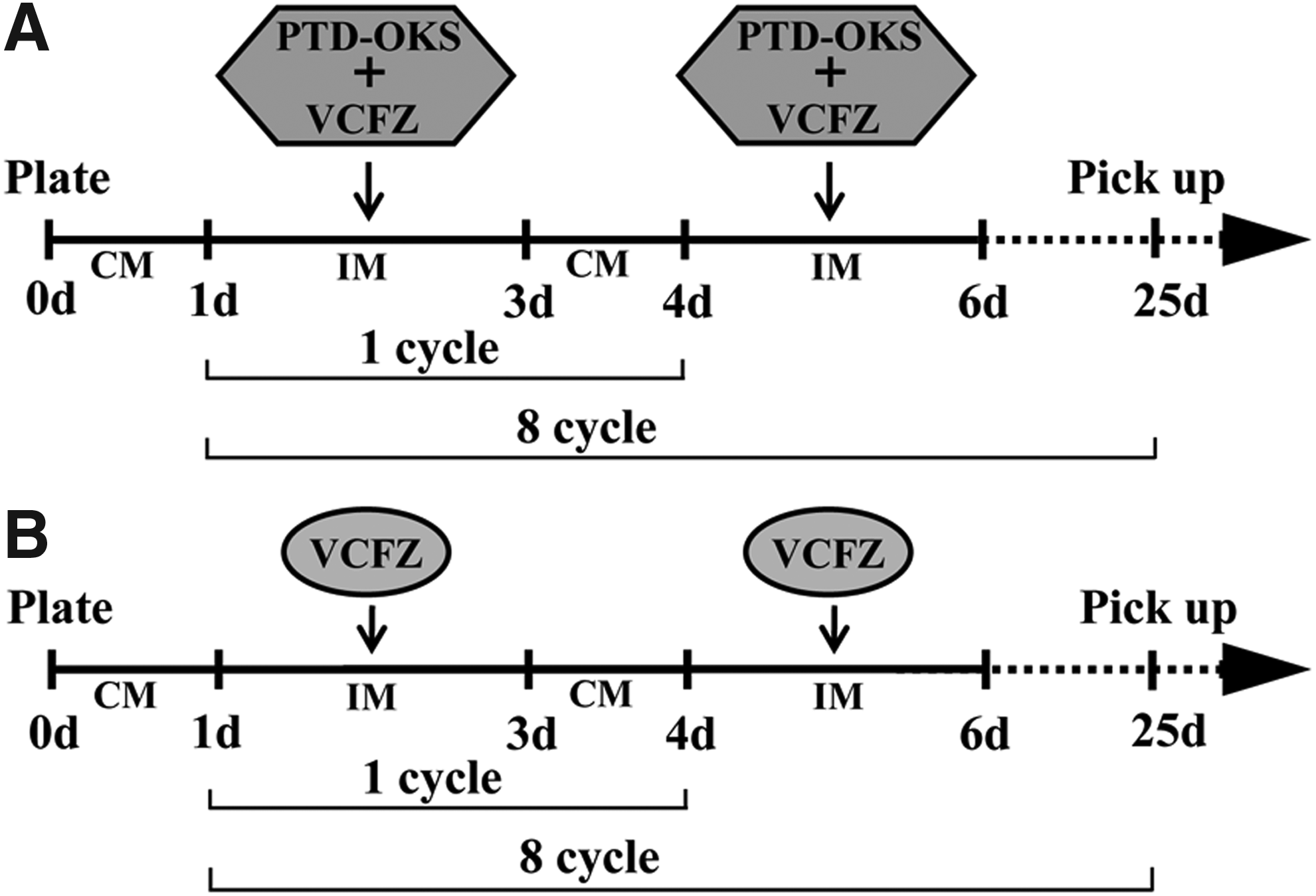
The schematic illustration of reprogramming groups and processes. The schematic illustrated the groups and the processes of non-genetic direct reprogramming through VCFZ+OKS
Reverse transcription–polymerase chain reaction analysis
After 25 days of culture, total RNA from ADSCs was isolated using Tissue RNA Miniprep Kit, and the resulting RNA samples were quantified by measuring the optical density (OD) at 260 nm; the OD 260/280 ratios for all RNA samples were between 1.8 and 2.1. Total RNA (1 μg) was reverse transcribed in a 10 μL reaction mixture containing 2 μL 5 × RT Buffer, 0.5 μL RT Enzyme Mix, 0.5 μL Primer Mix, and 6 μL nuclease-free water at 42°C for 1 hour. One tenth of the RT product was used for subsequent polymerase chain reaction (PCR) with the final concentration of PCR reaction being 1 × Buffer, 0.2 mM dNTPs, 1.25 U Blend Taq in a total volume of 50 μL, using primers shown in Table 1. The PCR mixture was first denatured at 94°C for 2 minutes then amplified for 30 cycles (94°C, 30 seconds; 60°C, 30 seconds; and 72°C, 1 minute) using an authorized thermal cycler (Eppendorf, Hamburg, Germany). After amplification, 5 μL of each PCR product and 1 μL of 6 × loading buffer were mixed and electrophoresed on a 1.5% agarose gel in 0.5 × Tris-boric acid-EDTA containing 0.5 μg/mL ethidium bromide. Gels were photographed and scanned.
Immunofluorescence assay
Immunofluorescence was used to identify the ADSCs. Briefly, after fixation in 4% paraformaldehyde for 30 minutes at room temperature, ADSCs were permeabilized with 0.1% Triton X-100 in PBS for 15 minutes at room temperature, followed by washing three times with PBS and incubation with PBS containing 10% FBS for 30 minutes at room temperature. The cells were then incubated with rabbit monoclonal anti-Oct4 (1:400 dilution), anti-Sox2 (1:400 dilution), anti-Nanog (1:400 dilution), and mouse polyclonal anti-Klf4 (1:400 dilution) for 60 minutes, and then with the goat anti rabbit or mouse secondary antibodies for 60 minutes at room temperature. This was followed by washes with PBS, twice for a duration of three minutes each. DAPI staining was performed to visualize the nucleus. Immunofluorescence was then examined by the microscope.
Statistical analysis
The values were expressed as mean ± SD from three to six samples. Statistical analyses were carried out using Student's t-test and a one-way analysis of variance (SPSS 16.0, Inc., Chicago, IL). Results of p < 0.05 were considered statistically significant. All assays were performed at least with three replicates.
Results
Characterization and differentiation of hADSCs
hADSCs were isolated from human lipoaspirate tissue (Fig. 2A). A confluence of 80%–90% was reached after 10 days of culture, ADSCs were spindle-shaped under phase contrast microscope (Fig. 2B, C). And alkaline phosphatase staining was positive (AP+) under phase contrast microscope (Fig. 2D). Adipogenic differentiation was detected by the formation of lipid droplets in the cytoplasm of the cells, which were stained with Oil Red O. Accumulation of lipid droplets was noticed in the cells, which underwent adipogenic differentiation for 2 weeks (Fig. 2E), while no lipid droplets were noticed in ADSCs (Fig. 2F) (negative control). Osteogenic differentiation was demonstrated by calcification areas shown by Alizarin red staining (Fig. 2G), which was not visible in the negative control (Fig. 2H).

The characterization and differentiation of hADSCs. Primary fibroblast-like ADSCs culture at 3 days
Effect of PTD-OKS and cocktail of chemical compounds on survival of ADSCs
Compared with control media, PTD-OKS moderately promoted the proliferation of ADSCs with the concentrations of 50 μg/mL based on CCK-8 assay. The numbers of ADSCs decreased after incubation with 60 or 70 μg/mL PTD-OKS (Fig. 3A). Compared to control media, small molecular cocktail VCFZ significantly inhibited the proliferation of ADSCs based on CCK-8 assay. When VCFZ was combined with PTD-OKS, slight inhibition of the proliferation ability of ADSCs were noticed in comparison to the control group (Fig. 3B).
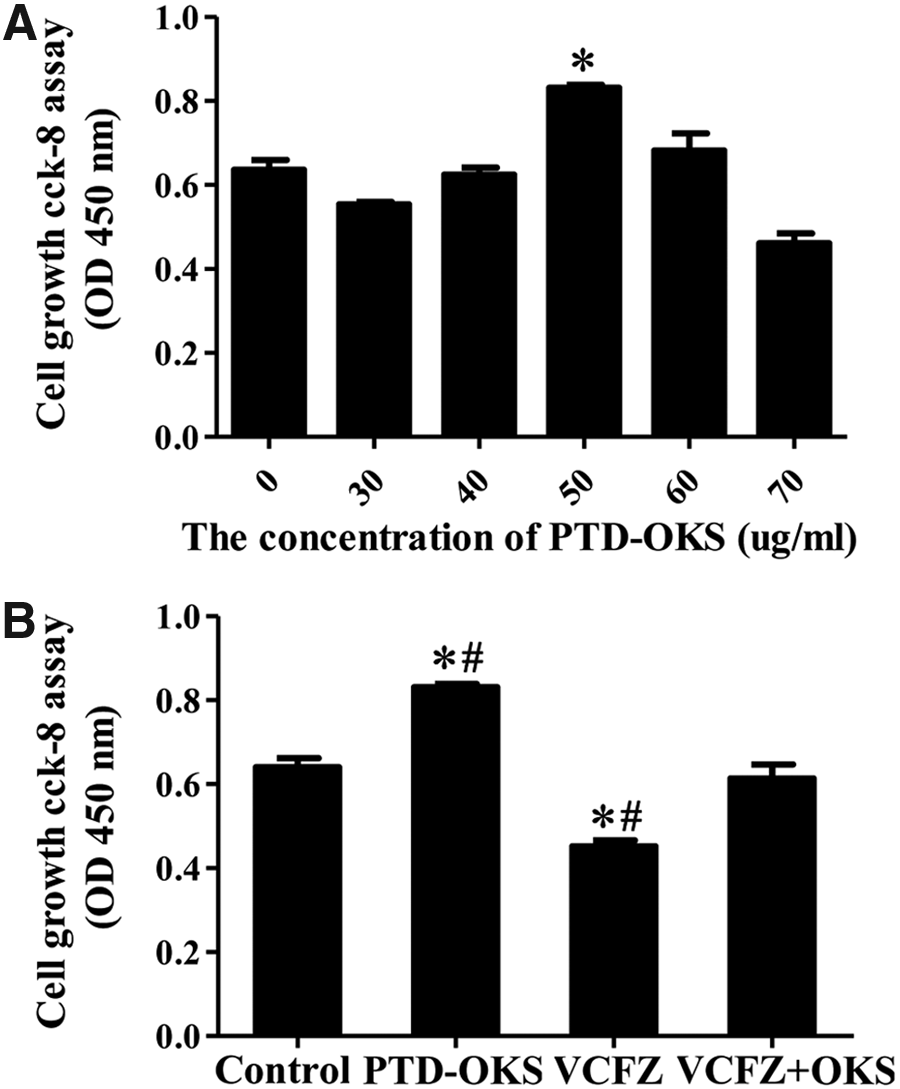
The survival assay of hADSCs treated with reprogramming reagents.
The conversion of hADSCs after non-genetic reprogramming treatment
Primary hADSCs were treated with reprogramming reagents in group VCFZ+OKS, group VCFZ and group control for eight cycles. After 15 days of treatment, ADSCs in the VCFZ+OKS group and VCFZ group exhibited morphological changes of decreased adherence and aggregated growth. Cells proliferated largely and displayed dense colonies in VCFZ+OKS group after 25 days treatment (Fig. 4C), and many apoptotic cells appeared in the VCFZ group (Fig. 4F). However, ADSCs in control group still displayed spindle-shape cellular morphology with the lack of colony formation (Fig. 4I). We also calculated reprogramming by counting the alkaline-phosphatase-positive (AP+) colonies to determine the reprogramming efficiency (Fig. 5). In the VCFZ+OKS group, VCFZ group, and control group, the corrected efficiency of alkaline phosphatase positive (AP+) colony formation were 38.75%, 17.94%, and 9.18%, respectively. Although all treatment groups were significantly different from controls, the combination performed significantly better than either reagent alone. Thus, compared with VCFZ, the combination of VCFZ+OKS could enhance the pluripotency of ADSCs significantly.
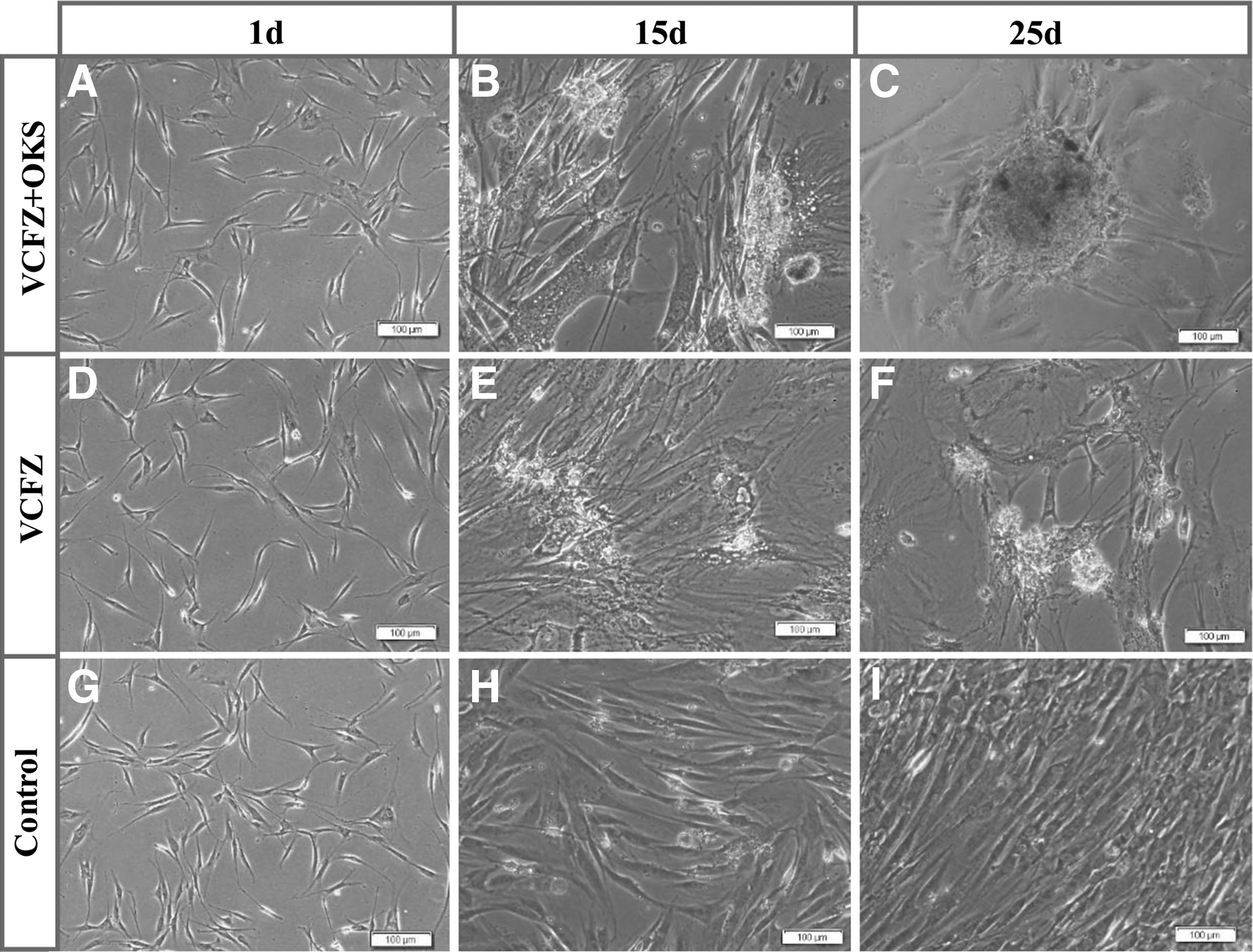
The cellular morphological conversion of hADSCs after the treatment of reprogramming reagents. The adherent hADSCs were cultured after 1
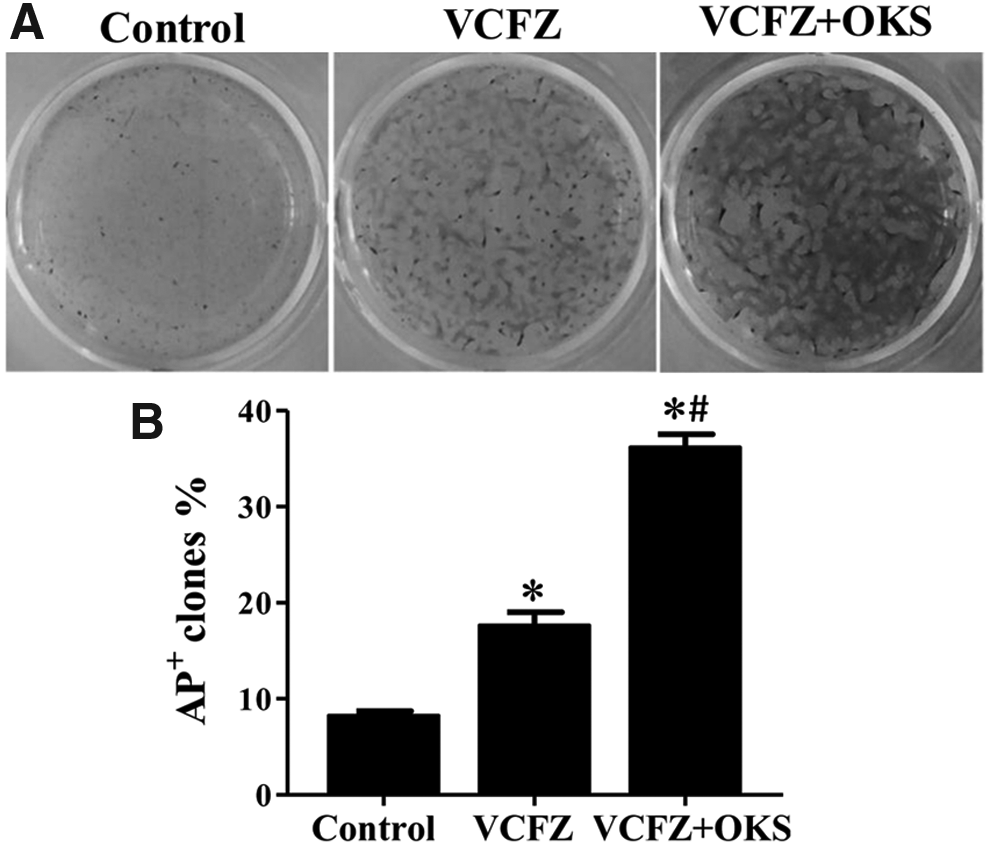
AP staining of ADSCs untreated or treated with reprogramming reagents in triplicate on 6-well plates. The reprogramming efficiencies were determined by counting AP+ colonies on day 25. Three groups were included: reprogramming with the VCFZ protocol, VCFZ+PTD-OKS protocol, and control group. Data are shown as mean ± SD. *p < 0.05 versus control group; #p < 0.05 VCFZ+PTD-OKS versus VCFZ. Difference with p < 0.05 (*/#) was considered statistically significant. VCFZ+PTD-OKS is abbreviated as VCFZ+OKS.
Cocktail of chemical compounds enhance expression of pluripotent genes
A large number of pre-iPSCs colonies appeared after VCFZ combined with PTD-OKS induction for 25 days. And immunofluorescence staining showed that these colonies were positive for pluripotency makers OCT3/4 (Fig. 6A), KLF4 (Fig. 6B), SOX2 (Fig. 6C), NANOG (Fig. 6D), while negatively expressed OCT4 (not show). However, ADSCs weakly expressed KLF4 (Fig. 6F) and NANOG (Fig. 6H) in VCFZ group by immunofluorescence staining and control group does not express anything (Fig. 6I–L). Furthermore, reverse transcription (RT)-PCR analysis showed that the gene transcript of OCT3/4, KLF4, SOX2, and NANOG were positively expressed after 25 days of treatment with VCFZ+OKS in hADSCs spheroids. And, the gene transcript of KLF4 in hADSCs after 25-day VCFZ treatment was positively expressed.
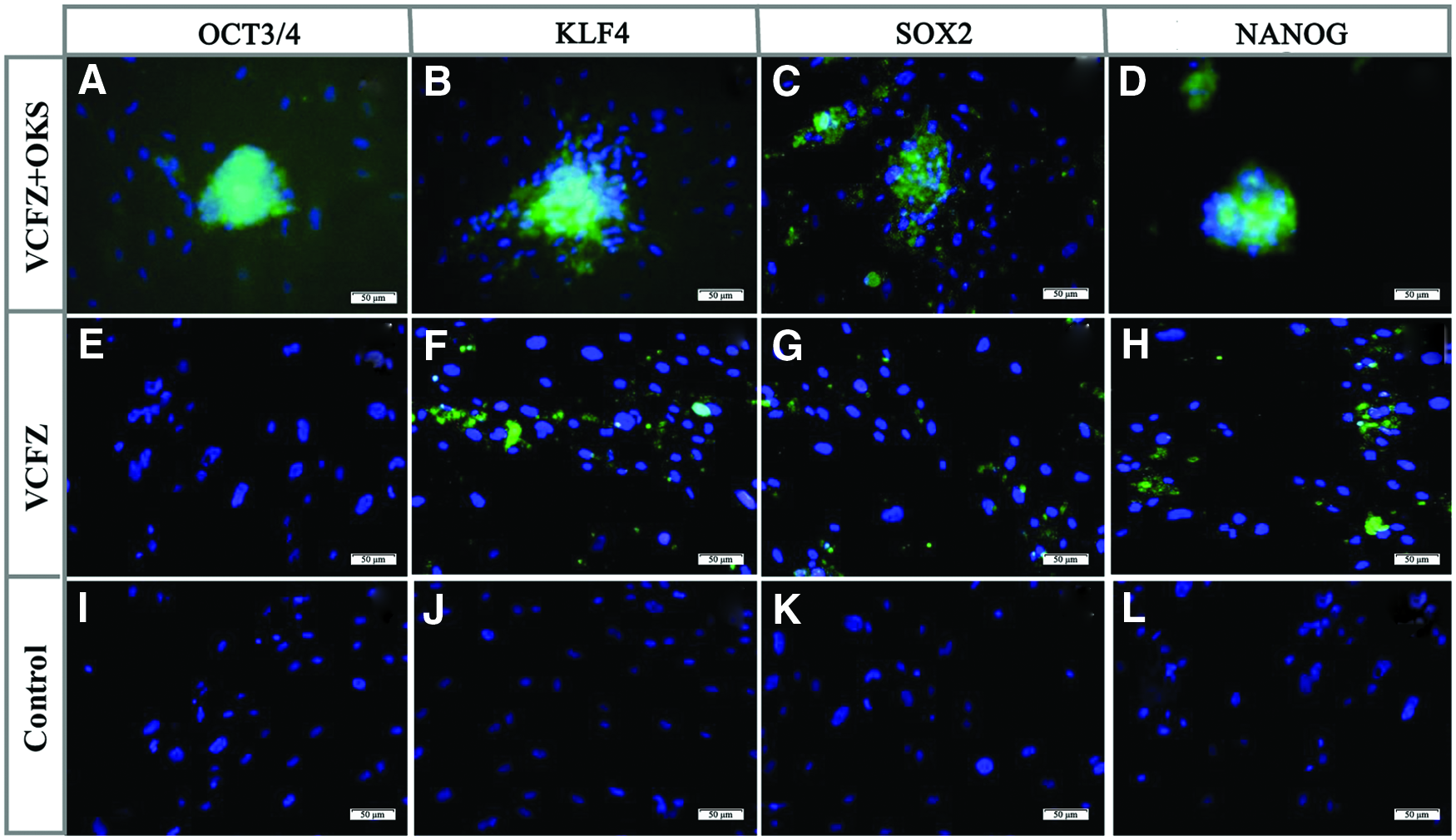
The immunofluorescence staining of hADSCs after the treatment of reprogramming reagents. ADSCs aggregate spheroids in VCFZ+OKS group were positively stained for OCT3/4
However, ADSCs in conventional culture without VCFZ+OKS or VCFZ treatment did not express OCT3/4, KLF4, SOX2, and NANOG (Fig. 7A). GAPDH was expressed in all ADSCs. To understand the regulation of pluripotent genes by VCFZ+OKS on ADSCs, we conducted quantitative PCR experiments to evaluate the gene expression of OCT4, KLF4, SOX2, and NANOG. We found the cocktail of chemical compounds VCFZ combined with recombinant protein PTD-OKS significantly upregulated the expression of OCT4, KLF4, and SOX2 gene, especially NANOG (Fig. 7B, C). These data demonstrate that VCFZ+OKS may induce ADSCs reprogramming by increasing mRNA expression of OCT4, KLF4, SOX2, and NANOG gene.

RT-PCR analysis for the gene expression of OCT3/4, KLF4, SOX2, and NANOG in hADSCs after reprogramming treatment. The gene expressions of OCT3/4, KLF4, SOX2, and NANOG of hADSCs were positively displayed after 25 days treatment of VCFZ+OKS, while hADSCs just express KLF4 gene after 25 days culture with VCFZ treatment, ADSCs in conventional culture without VCFZ+OKS or VCFZ treatment express GAPDH
Discussion
Traditional reprogramming methods are laborious and can cause oncogenic expressions that are unfavorable. Yamanaka factors are able to reprogram somatic cells to become iPSCs. Nonetheless, the generation of iPSCs is laborious and inefficient. Moreover, potential risks exist during integration because of the oncogene expression for iPSCs generation (Belyi et al., 2010; Puzio-Kuter and Levine, 2009). Somatic cell reprogramming has recently prompted the study on direct lineage conversion between two mature cells can generally be accomplished in a short time, while proving to be more efficient and safe (Dai et al., 2014; Gascon et al., 2017; Ii, 2017). Somatic stem- and progenitor cells in adults share similar features as pluripotent stem cells, hence providing an efficient source of iPSCs. For example, immature cell populations of the hematopoietic lineage, in general, give rise to iPSCs at higher efficiencies than terminally differentiated cell types (Eminli et al., 2009).
There are many advantages of using ADSCs as donor cells for reprogramming. First, the isolation and purification of ADSCs is simple and rapid. Second, a large number of ADSCs can be extracted from human adipose tissue and providing a good source for autologous cells. Third, ADSCs express AP activities and along with high endogenous expression level of KLF4, KLF2, KLF5, Esrrb, and c-Myc, which make ADSCs more plastic and fewer barriers for reprogramming. Many studies reveal that the generation of iPSCs from hADSCs is faster and efficient than adult human fibroblasts using Yamanaka factors (Sugii et al., 2010; Sun et al., 2009).
In this study, ADSCs could easily be isolated by collagenase digestion from human lipoaspirate tissues. ADSCs were positively differentiated into adipocytes, osteoblasts, and formed lipid droplets. Our previous studies show that primary hADSCs expressed MSC specific markers including CD29(+), CD44,(+) and CD59(+) but did not express CD45(−) and HLA-DR(−) (Dai et al., 2014; Guo et al., 2015, 2016). Additionally, Primary ADSCs also partly expressed CD34 and CD105, which showed that these ADSCs were comprised of heterogeneous cell populations. CD34 was quiescence stem cell and endothelial cell marker (Gaafar et al., 2014). Usually, CD105 expression was relatively low on the freshly isolated ADSCs. The expression level of CD34 decreased while that of CD105 increased with culture time (Baer and Geiger, 2012).
Recently, many studies have found that reprogramming can be done through new ways of non-gene manipulation (Inagawa and Ieda, 2013). It is safer and more controllable to perform cell reprogramming using small molecule compounds involved in regulating cell status. Recombinant proteins with cell penetrating peptides (CPPs) or protein transduction domains (PTDs) can also achieve non-genomic reprogramming (Thier et al., 2010). The combinative uses of small molecule VPA regimen and recombinant proteins with CPPs or PTDs showed significantly higher reprogramming efficiency than their separate application (Nordin et al., 2017; Zhang et al., 2012). PTD, also known as CPPs, which is a specific short peptide that mediates a variety of molecules (including small molecule compounds, nanomaterials, DNA, siRNA, proteins, etc.), transfected cells.
Several reports of recombinant protein induced iPS were use of arginine-rich peptide as a protein transport system (Chen et al., 2016; Heng and Fussenegger, 2014; Nordin et al., 2017; Singhal et al., 2015; Thier et al., 2010; Zhang et al., 2012; Zhou et al., 2009), such as Ding Sheng laboratory selection of 11R-PTD established the mouse protein-iPS, Kim's Laboratories established the human protein-iPS using 9R-PTD. Our previous study demonstrated that hADSC can successfully direct reprogrammed to corneal endothelial cells using PTD-OKS combined with a small molecular method (Dai et al., 2014). These results not only helped increase iPS security and made a breakthrough but also proved that the use of recombinant protein reprogramming or direct reprogramming is feasible. The disadvantage is that the recombinant protein iPS induction rate is 10 to 100 times lower than the virus-induced system, which greatly limits the application of protein iPS.
Small molecules antagonize or activate reprogramming cell signaling pathways that play a key role in regulating cell fate. Numerous reports indicate that small molecules can improve reprogramming efficiency and even induce iPS cells (Desponts and Ding, 2010; Hou et al., 2013; Li et al., 2011; Lu et al., 2013; Park et al., 2015). Previously, Li et al. (2011) reported that a small-molecule combination “VC6T” [VPA, CHIR99021 (CHIR), 616452, Tranylcypromine], is sufficient to permit reprogramming from mouse embryonic and adult fibroblasts in the presence of a single transcription factor, OCT4, within a period of days. Hou et al. (2013) found VC6T and Forskolin induced some green fluorescent protein (GFP)-positive clusters expressing E-cadherin, however, the expression of OCT4 and Nanog was not detected. To achieve complete chemical reprogramming without the OCT4-inducible system, many small molecules were further tested and they identified that OCT4 expression was enhanced significantly after the addition of DZNeP in chemical reprogramming, and proving that DZNeP was critical for stimulating the expression of OCT4 but not the other pluripotency genes (Hou et al., 2013).
In these small molecules, they found that four essential small molecules VCFZ whose individual withdrawal from the cocktails generated significantly reduced GFP-positive colonies and no CiPSCs. But in our experiments, ADSC cells were treated for 25 days of VCFZ and no cell colony was visible, yet it was noticed that a large number of cells underwent necrosis. Immunofluorescence identification showed that only KLF4 was positively expressed in these ADSCs. There was a negative expression of OCT4, SOX2, and NANOG. Hence, we doubt that VCFZ is inefficient to reprogram ADSCs thereby prompting us to use a combination of PTD-OKS and VCFZ. The combination showed aggregated growth and positive staining.
Overall, using reprogramming proteins PTD-OKS combined with VCFZ, ADSCs easily formed aggregated growth and were positive for AP staining. Our study demonstrates that the combination of PTD-OKS proteins and VCFZ displayed higher cell survival and lower apoptosis than other reprogramming reagents. ADSCs were positive for iPSC markers after 25 days treatment of PTD-OKS proteins combined with VCFZ, such as OCT3/4, KLF4, SOX2, and NANOG. We hypothesize that this method can provide an unlimited cell resource and hold great potential for in vitro cell differentiation and in vivo tissue generation in the future.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.