
Abstract
Abstract
As a potential cure for diabetes, more and more attentions have been paid to organ transplants to replace insulin therapy. As a result, many researchers have explored out many programs to get insulin-producing cells (IPCs) to replace the defective β cells. Currently, more and more new induction methods are being proposed, and at the same time, more and more possible induction molecular mechanisms are being revealed. The purpose of this study was to explore whether and how the two factors pdx-1 and myocardin affected the differentiation of rat mesenchymal stem cells (rMSCs) into IPCs. In this study, we investigated the process of transfecting myocardin and/or pdx-1 in rMSCs in vitro. The results showed that rMSCs were able to secrete insulin after cotransfected with myocardin and pdx-1. At the same time, we explored the possible mechanism that myocardin and pdx-1 coinduced rMSCs into IPCs by forming a complex to promote the transcriptional activity of insulin. Our results may provide a theoretical basis to the study of islet transplantation in the future.
Introduction
D
Many kinds of pluripotent stem cells can be used to be induced into IPCs as embryonic stem cells, umbilical cord blood, and mesenchymal stem cells (MSCs) (Bose et al., 2012; Kadam et al., 2010; Kim et al., 2012; Milanesi et al., 2012; Parekh et al., 2009; Raikwar and Zavazava, 2013). In particular, MSCs are very concerned, due to the low immunogenicity and multienergy (Pittenger et al., 1999; Ren et al., 2012; Jiang et al., 2002; Kobayashi et al., 2009). Although there have existed a lot of induced methods, the further effective methods and distinct mechanisms of MSCs differentiation into IPCs should also be concerned about.
Myocardin can form a stable ternary complex with SRF to activate target genes, by binding to the highly conversed CarG box (CC(A/T)6GG sequence) (Wang et al., 2001). Interestingly, there exists a CarG box in the promoter of insulin. This may indicate that myocardin probably interacts with insulin. Moreover, pancreatic duodenal homeobox 1 (pdx-1) is a well-known master gene in pancreas development and crucial for early pancreatic differentiation (Fujitani et al., 2006; Holland et al., 2005). It has been proved that stable IPC lines could be isolated from MSCs by transduced with the pdx-1 (Karnieli et al., 2007; Li et al., 2007).
So far the relationship between myocardin and pdx-1 is not clear. In this article, we investigated the effect of the two factors on the inducting of rat MSCs (rMSCs) differentiation and the molecular mechanisms of their interaction.
Materials and Methods
Cell isolation and culture
Adult bone marrow-derived MSCs were isolated from the femurs and tibias of male Sprague–Dawley rats (weight 90–100 g, aged 4 weeks) with a method described by Wang et al. (2013a). In brief, bone marrow mononuclear cells were obtained by Percoll (1.073 g/mL) density gradient centrifugation. The cells were seeded in Dulbecco's modified Eagle's medium-low glucose (DMEM-LG, Hyclone Co.) supplemented with 10% fetal bovine serum (FBS, Gibco) at 37°C in humified air with 5% CO2. Cos-7 cells (ATCC, Number CRL-1651) were cultured in DMEM containing 10% FBS.
Plasmids used and cell transfection
The CDS (sequence coding for amino acids in protein) regions of myocardin and pdx-1 were cloned into pLVX-Puro and the vector of pLVX-Puro as control. For transfection experiments, the rMSCs were cultured in growth medium without antibiotics at 60% confluence for 2 days and then transducted with lentiviral(MOI:5).
Then insulin promoter region was cloned into pGL-3 luciferase reporter vector and amplified by polymerase chain reaction (PCR) with sense primer containing MluI: GACATACGCGTGAAAGGGTGTGATCCACAAAAACTG, and antisense primer containing HindIII: AGGGAAGCTTAGAAAATCCTCACATTTATCTACAA. Before transfection, the rMSCs should be cultured for 2 days in medium without antibiotics at 60% confluence. Then the luciferase analysis, immunocytochemistry assay, real-time PCR, and western blotting were performed as follows.
Immunocytochemistry assay
This experimental method is based on our earlier introduction (Wang et al., 2013a). In brief, the cells after treatment were fixed in 4% paraformaldehyde for 15 minutes and then blocked with normal goat serum for 20 minutes at room temperature (RT). After incubation with primary antibodies (rabbit anti-pdx-1 [Protein Tech]) in a humid chamber over night, cells were incubated with appropriate secondary antibodies (FITC-conjugated goat anti-rabbit IgG, Santa Cruz) for 30 minutes at 37°C. After washing with PBS, the samples were observed under laser scanning confocal microscope (OLYMPUS, Japan). DAPI stain (blue) highlights the total nuclei.
Reverse-transcription PCR and real-time PCR
This experimental method is based on our earlier introduction (Wang et al., 2013a). In brief, total RNA was isolated from cells using TRIzol Reagent (Invitrogen), and two micrograms of the sample were reverse-transcribed using M-MLV reverse transcriptase (Promega). Real-time PCR was performed in an Applied Biosystems StepOne™ Real-Time PCR System. Fast SYBR® Green Master Mix was obtained from Applied Biosystems. The reverse-transcription PCR (RT-PCR) primers sequences are listed in Table 1. Data were shown as relative expression level after being normalized by GAPDH. Primers were used for PCR as listed in the table.
Western blotting
This experimental method is based on our earlier introduction (Wang et al., 2013b). In brief, proteins prepared were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. The membranes were immunoblotted with rabbit anti-myocardin (Protein Tech), rabbit anti-pdx-1 (Protein Tech), and rabbit anti-insulin (Protein Tech) antibodies overnight at 4°C and then incubated with IRDyeTM-800 conjugated anti-mouse and anti-rabbit secondary antibodies (Li-COR Biosciences) for 30 minutes at RT. The specific proteins were visualized by Odyssey Infrared Imaging System (Gene Company Limited). β-actin expression was used as an internal control to show equal loading of the protein samples.
Luciferase assay
This experimental method is based on our earlier introduction (Wang et al., 2013a). In brief, luciferase activity was measured using the Dual-Luciferase Assay System (Promega). Results were normalized by dividing the firefly luciferase activity with the Renilla luciferase activity of the same sample.
Measurement of insulin using enzyme-linked immunosorbent assay
This experimental method is based on our earlier introduction (Jian et al., 2015). In brief, for intracellular insulin quantification, rMSCs were cotransducted with myocardin and/or pdx-1 for 7 days, then differentiated cells were treated with DMEM-LG (5.6 mM) for 12–16 hours, washed with PBS, and then stimulated by DMEM-HG(25 mM) for 2 hours at 37°C. The supernatant was collected, and secreted insulin was quantified using rat insulin enzyme-linked immunosorbent assay (ELISA) kits (WUHAN BEINGLAY BIOTECH CO.) according to the manufacturer's instructions.
Chromatin immunoprecipitation assays
All chromatin immunoprecipitation (CHIP) assays were performed by SimpleChip® Enzymatic Chromatin IP Kit (Cell Signaling Technology, Danvers, MA). In brief, after 48 hours 4 × 105 transducted Cos-7 cells were crosslinked by formaldehyde (1% for 10 minutes) and then collected. After ultrasound, chromatin was sheared into 200–1000 bp length. 10% supernatant was used as 10% input; other chromatin was immunoprecipitated with 5 μg anti-myocardin antibody. The protein–antibody complexes were recovered on protein/G beads. Immunoprecipitates were washed three times, and crosslinks of chromatin were reversed by incubating samples 30 minutes at 65°C.
The resulting DNA was purified and analyzed by PCR. The following pairs of primers were used for each indicated promoter region as follows: CarG box, 5′-TCCAATCCCAGAGAGACCAG-3′ and 5′-TGGGAAGGGTGGACTCTTGG-3′, size 196 bp. Abox, 5′-GACCTCTGAAAAAATTTAAATATCA-3′ and 5′- GAAAGGAGCCTTGTAGTGTAGCTAG-3′, size 167 bp.
Coimmunoprecipitation
Lysates were collected after lentiviral of myocardin and pdx-1 transducted into rMSCs after 48 hours, pLVX-Puro as control. The myocardin antibody (Protein Tech) was used to precipitate myocardin, and pdx-1 antibody (Protein Tech) was used to precipitate pdx-1. The resulting mixture was washed, subjected to SDS-PAGE, transferred to a NC (nitrocellulose filter) membrane (Pall), first revealed by myocardin antibody to visualize myocardin, and then striped and reprobed with pdx-1 antibody to visualize pdx-1.
Statistical analysis
Data are expressed as mean ± SE, accompanied by the number of experiments performed independently, and analyzed by t-test. Differences at p < 0.05 were considered statistically significant.
Results
Myocardin and pdx-1 could synergistically promote expression of insulin in rMSCs
First, the impact of myocardin and pdx-1 on the differentiation of rMSCs were explored by testing the expression of insulin. To determine that, after overexpression of myocardin and/or pdx-1 in rMSCs by lentiviral transduction, we tested the mRNA and protein levels of the key genes by RT-PCR, real-time PCR, and western blotting. The results showed that the mRNA and protein levels of insulin were increased in the group of pdx-1 overexpressed and coexisted in the group of two factors. Especially in the cotransfection group, the improvement was most remarkable (Fig. 1A–C and Supplementary Figs. S1 and S3).
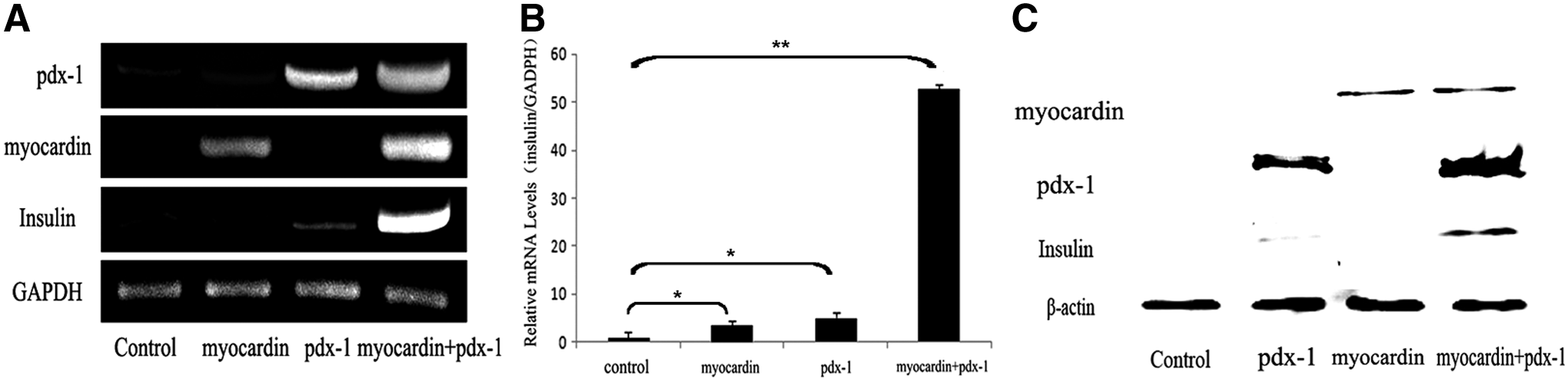
Myocardin and pdx-1 synergistically promote expression of insulin in rMSCs. PLVX-Puro (as control) myocardin and/or pdx-1 were transducted into rMSCs for 48 hours, the mRNA levels of myocardin, pdx-1, and insulin were determined by reverse-transcription-PCR
Insulin release in response to the expression of myocardin and pdx-1
We observed the change of insulin in molecular level after overexpression of myocardin and pdx-1 in rMSCs above. How about the secretion of insulin in rMSCs after transduction? The rMSCs after transduction were subjected to immunocytochemistry, using the antibody specifically against insulin. In the control group, cells transduced with pLVX-Puro were not detected with insulin secretion, while a strong insulin expression was found in the rMSCs cotransducted with myocardin and pdx-1 (Fig. 2A and Supplementary Fig. S2).
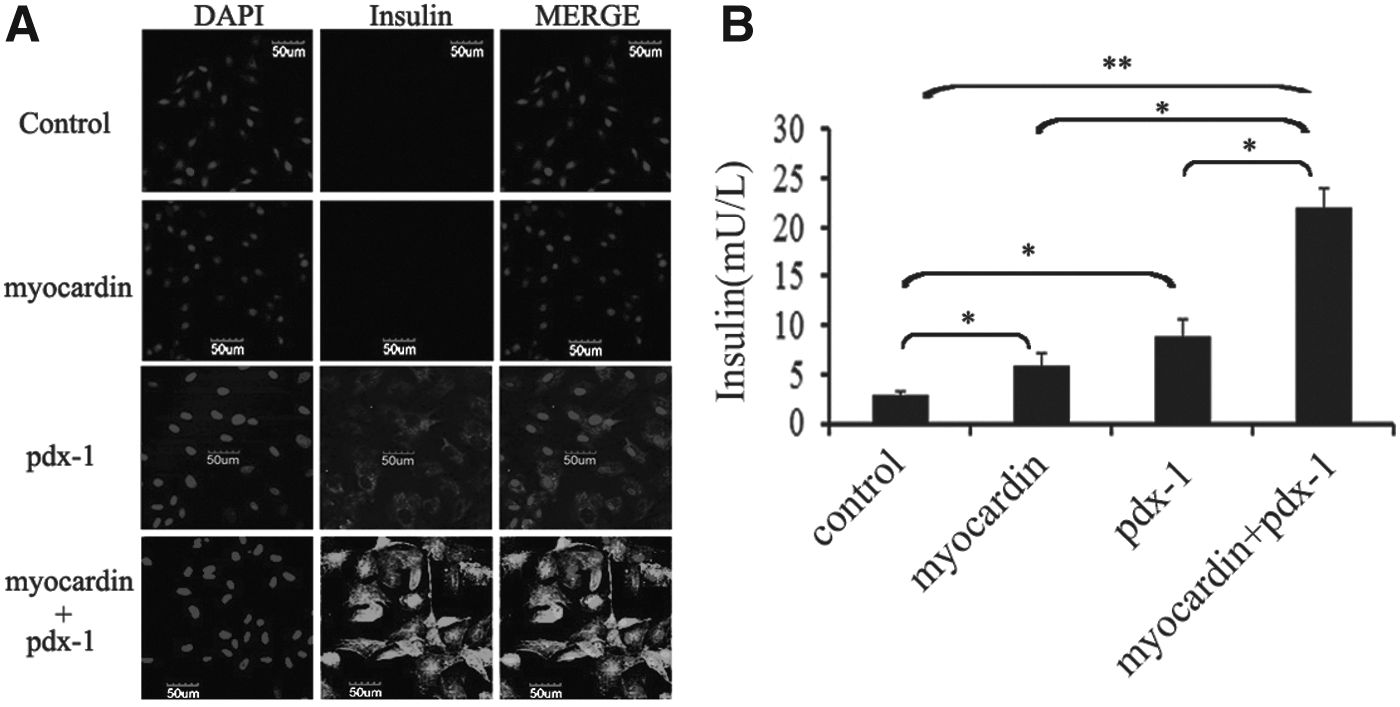
Insulin release in response to the expression of myocardin and pdx-1. Immunofluorescence assay for insulin showed that rMSCs can exhibit insulin positive expression. Nuclei were stained with DAPI
Based on the above results, enzyme-linked immunosorbent assay was used to evaluate the secretion of insulin. The results were in line with previous studies, insulin production was increased in response to the coexpression of myocardin and pdx-1 than undifferentiated cells (Fig. 2B). These results showed that rMSCs could be induced into IPCs, which was similar to the pancreatic β cells under the overexpression of myocardin and pdx-1.
Physical interaction between myocardin and pdx-1 in vitro
Since myocardin and pdx-1 could synergistically induce rMSCs into IPCs, we hypothesized that there existed a link between myocardin and pdx-1 in vivo. Coimmunoprecipitation assays were used to measure the link after myocardin and pdx-1 overexpressed in Cos-7 for 48 hours. In our results, a direct physical interaction between the two factors was detected in vivo (Fig. 3). Thus, myocardin and pdx-1 could form a complex in vivo.
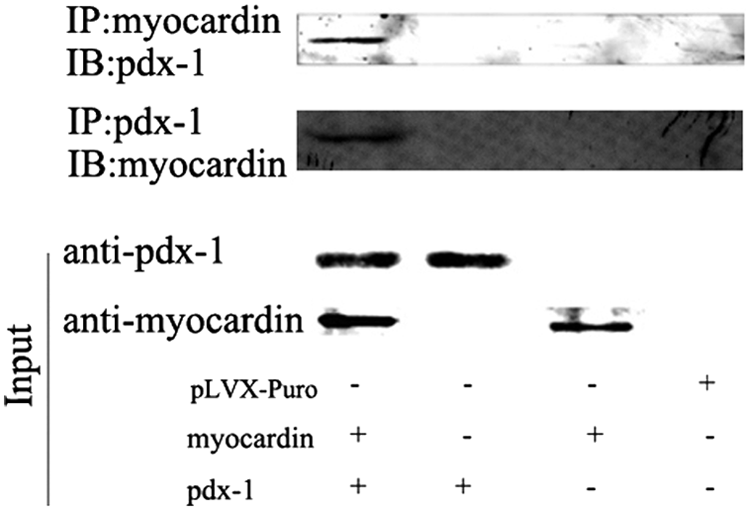
Physical interaction between myocardin and pdx-1 in vivo. Cotransducted with myocardin and pdx-1 for 48 hours. Coimmunoprecipitation assays were used to identify the interaction of myocardin and pdx-1 in cellular environment.
The synergy of two factors on insulin expression was mediated via pdx-1/myocardin/SRF/CarG box pathway
Unlike pdx-1 which is directly bonded to A box (TAAT), myocardin must first bind with SRF and then activate the target genes with CarG box-containing promoters. As shown in Figure 4A, myocardin and pdx-1 could increase the transcriptional activity of insulin. In an attempt to detect the mechanism, myocardin and pdx-1 mediated regulation of insulin expression, the CarG box, and A box in insulin promoter was mutated (5′-CCTATAAAGG-3′ to 5′-AATAGGGAAA-3′ and 5′-TAAT-3′ to 5′-TTTT-3′). We got a negative result in the group of cotransfection of pdx-1 and myocardin with the CarG box-mutated insulin promoter as well as the CarG box and A box double-mutated insulin promoter, compared with the wild type (Fig. 4B, D), although the luciferase activity in the group of cotransfection of two factors was not affected when the A box was mutated (Fig. 4C).
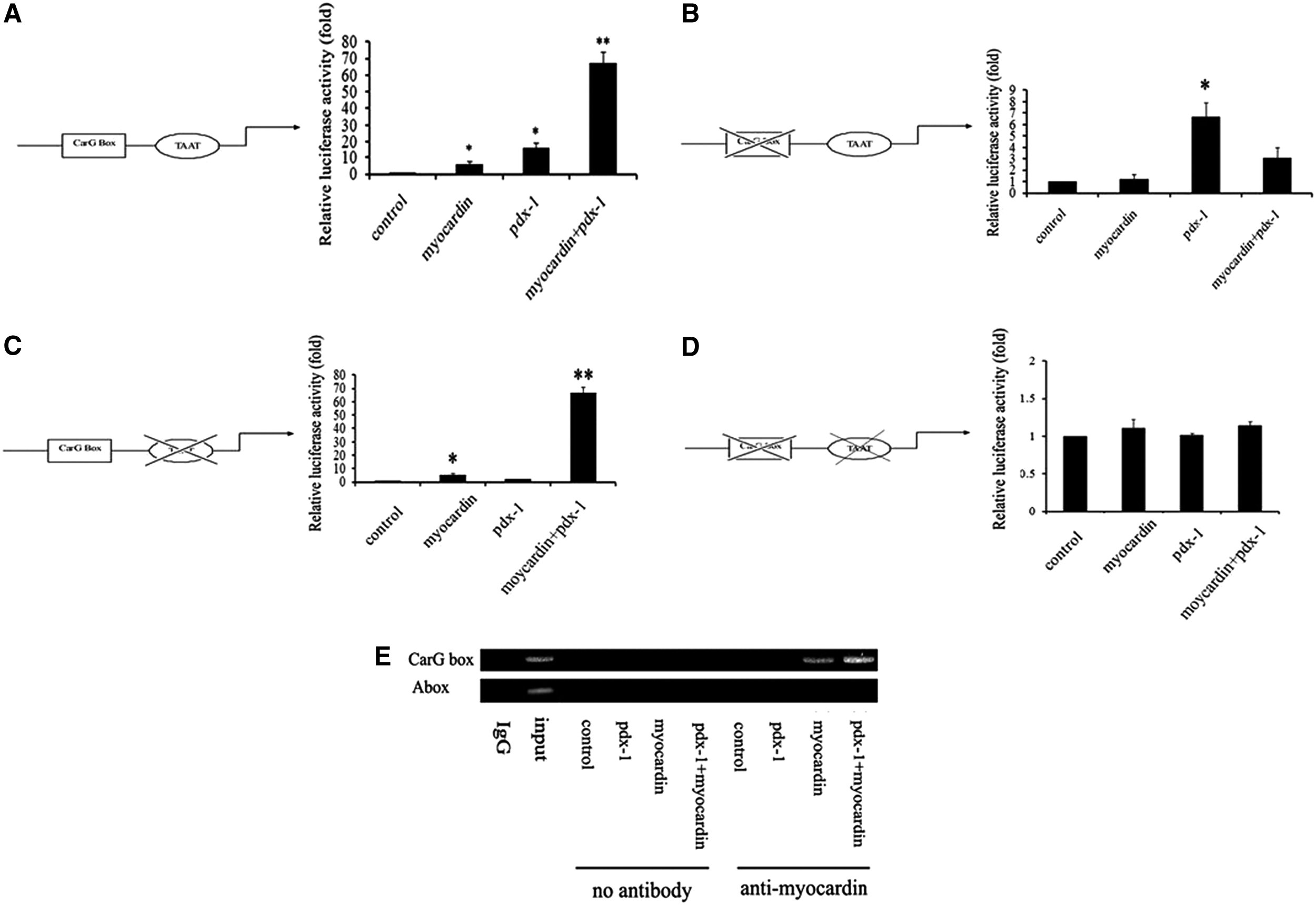
Transactivation of insulin induced by myocardin, and pdx-1 was mediated via myocardin-SRF-CarG box signal transduction. The activation effect of myocardin and pdx-1 on wild type
Based on above results, we suspected that the active effects of insulin transcriptional activity mediated by myocardin and pdx-1 might be due to the myocardin-SRF-CarG box signal transduction. To further confirm our hypothesis, ChIP experiments were carried out after Cos-7 cells were transfected with myocardin, pdx-1, and the insulin promoter. As shown in Figure 4E, myocardin and pdx-1 could bind to the CarG box region of insulin promoter. So we have reasons to believe that the upregulation of insulin induced by myocardin and pdx-1 was mediated via the myocardin-SRF-CarG box signal transduction.
Discussion
In this study, our main focus was to evaluate the potential of myocardin and pdx-1 in inducing rMSCs to differentiate into IPCs. Also, we hoped to find a possible link between the two factors. Fortunately, we found a synergy of two factors on insulin secretion with a short period about 7 days, which was even better than many differentiation conditions previously reported (Santamaria et al., 2011; Wang et al., 2011).
Recently, there have been some new induction schemes. Galivo made GBCs (gallbladder and cystic ducts) be reprogrammed into insulin-producing pancreatic β-like cells by a combined adenoviral-mediated expression of hallmark pancreatic endocrine transcription factors PDX1, MAFA, NEUROG3, and PAX6 (Galivo et al., 2017). Some researchers create three-dimensional organotypic pancreatic cultures, named pancreatoids, composed of embryonic day 10.5 murine epithelial progenitors and native mesenchyme treated with (−)-indolactam-V or phorbol 12-myristate 13-acetate, two protein kinase C activators. They observed roughly 23% insulin+ cells at day 7 (Marissa et al., 2017). Compared with the above schemes, our induction method may be effective and easy to implement.
It had been confirmed that overexpression of pdx-1 could be a direct way to induce MSCs into β cells in vitro (Karnieli et al., 2007), which was consistent with our data. What was interesting in this article was that we found myocardin and pdx-1 could synergistically induce rMSCs to differentiate into insulin secreting cells. The two factors had an obvious effect not only in the expression of insulin but also the secretion of insulin. Of course, the most puzzled question was why and how myocardin and pdx-1 synergistically induce differentiation of rMSCs.
In this article, our data demonstrated a direct physical interaction between myocardin and pdx-1 in vivo. Myocardin and pdx-1 could form a complex to promote the transactivation of insulin. In addition, as we know, myocardin combined with SRF binding into CarG box to activate transcription, while pdx-1 directly bonded to the A box. Precisely, there were one CarG box and one A box in the insulin promoter. Luciferase reporter assay and CHIP assay coconfirmed that the synergy of two factors on insulin expression was mediated via pdx-1/myocardin/SRF/CarG box pathway.
In simple terms, cotransfecting myocardin and pdx-1 resulted in synergistic activation of the mutation of the A box in insulin promoter, while mutation of the CarG box in insulin promoter potently reduced the ability to respond to myocardin and pdx-1. Mutation of the CarG box almost completely eliminated responsiveness to myocardin and pdx-1.
Generally speaking, we first confirmed that co-overexpression of myocardin and pdx-1 could make rMSCs differentiate into insulin-secreting cells within a short day in vitro. At the same time, our data established a molecular link between myocardin and pdx-1. Obviously, further studies are needed to clarify whether the differentiated cells could treat diabetic nude mice model. As a conclusion, myocardin and pdx-1 could contribute to the expression and secretion of insulin in a very short period of time. Also, the synergy of two factors on insulin expression was mediated via pdx-1/myocardin/SRF/CarG box pathway.
Funding
This work was financially supported by the Wuhan University of Science and Technology school fund (No.2015XZ050) and National Natural Science Foundation of China (No.31471282).
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.