
Abstract
Abstract
To improve the cloning efficiency of somatic cell nuclear transfer (SCNT) and to establish nuclear transfer embryonic stem cells (NT-ESCs) reliably, it is necessary to produce high-quality blastocysts derived from mice SCNT embryos. Therefore, the present study aims to investigate an optimal method for mouse SCNT embryo production and NT-ESCs derivation by comparing the effects of two methods: the treatment of histone deacetylase inhibitor PXD101 after SCNT, embryo aggregation and their combination treatment. The results suggest that embryo aggregation at four-cell stage and 50 nM PXD101 treated for 10 hours during and after activation could improve both mouse SCNT embryos' development (PXD101: 40.0% vs. 18.5%; p < 0.05; aggregation: 40.2% vs. 18.5%; p < 0.05) and also enhance the isolation rate of NT-ESCs (PXD101: 38.2% vs. 12.5%; p < 0.05; aggregation: 39.0% vs. 12.5%; p < 0.05). The combination of their treatments had a higher development rate (43.6%) and significantly higher NT-ESCs isolation rate (54.7%), therefore, we concluded that the combination of these two methods (50 nM PXD101 treated for 10 hours after SCNT and then aggregated at four-cell stage) is considered as the optimal way for the in vitro development of SCNT embryo and subsequent NT-ESCs isolation in mice, providing a new approach for the practical improvement of mouse cloning techniques and opening new opportunities to improve cloning efficiencies in other species.
Introduction
S
Several studies have indicated that embryo aggregation is a promising method to improve the developmental competence of porcine blastocysts (Lee et al., 2007; Saadeldin et al., 2014, 2015; Siriboon et al., 2014; Terashita et al., 2011). Lee et al. (2007) observed in porcine IVF (in vitro fertilization) embryos that the rate of blastocyst formation and total cell number of blastocysts were higher in aggregated embryos than in control IVF embryos. The study by Terashita et al. (2011) found that the aggregation of embryos at the four-cell stage improved the quality of miniature pig SCNT blastocysts. Siriboon et al. (2014) produced viable cloned miniature pigs by aggregation of handmade cloned embryos at the four-cell stage. Saadeldin et al. (2014, 2015) suggested that aggregation of blastomeres is an efficient alternative for the development of parthenogenetic porcine embryos.
In addition, studies indicated that embryo aggregation is also a promising method to improve the efficiency of cloned porcine ESCs derivation (Lee et al., 2006, 2013). In mice, our previous study (Qiu et al., 2017) indicated that the embryo aggregation improved the development of parthenogenetic blastocyst and parthenogenetic ESCs (pESCs) derivation, which is consistent with the study by Shan et al. (2012). However, there is limited study about the effect of embryo aggregation on the development of mice SCNT embryo and NT-ESCs derivation efficiency. Therefore, the first aim of this study is to improve the blastocyst formation and the quality of SCNT blastocysts by the aggregation of genetically identical clones at the four-cell stage.
In addition to embryo aggregation, studies have indicated that application of the histone deacetylase inhibitors (HDACis) is also an effective method to improve the development of SCNT embryo in animal species, including mice, pigs, cattle, and sheep [see our review article (Qiu et al., 2016)], and our previous study (Qiu et al., 2017b) investigated the effect of a novel HDACi-PXD101 [belinostat, N-hydroxy-3-(phenylsulfamoylphenyl)acryl amide] on mouse SCNT embryo and concluded that the treatment of 50 nM PXD101 for 10 hours after activation could most effectively improve the in vitro developmental capacity of mouse SCNT embryos.
Therefore, in the present study, besides the investigation of the effect of embryo aggregation on the development of mouse SCNT embryo, we compared its effect with the treatment of PXD101, and the effect of these two treatments (aggregation and PXD101) on subsequent establishment of NT-ESCs, aiming to do the optimization of mouse SCNT embryo production and NT-ESCs derivation.
Materials and Methods
Unless otherwise indicated, all reagents were purchased from Sigma.
Animals
Kunming female mice were used as oocytes and somatic cell (cumulus cell) donors. All animals were maintained in accordance with the Animal Experiment Hand Book at the Center for Developmental Biology, and experiments were carried out in accordance with the UK Animals (Scientific Procedures) Act, 1986 and associated guidelines.
Collection of oocytes and cumulus cells
Mature oocytes were collected from the oviducts of females 8–12 weeks of age that were induced to superovulate with 5 IU of pregnant mare serum gonadotropin, followed by 5 IU of human chorionic gonadotropin (hCG) 48 hours later. Oocytes were collected from oviducts 16 hours after hCG injection, placed in Hepes-buffered CZB (H-CZB) medium, and treated with 0.1% hyaluronidase until the cumulus cells dispersed. The oocytes were transferred to fresh droplets of H-CZB and denuded of almost all cumulus cells by gentle pipetting. The denuded oocytes were then washed twice in H-CZB and then kept in K-modified simplex optimization medium (KSOM) containing nonessential amino acid (EAA) and EAA supplemented with 1 mg/mL bull serum albumin, covered with paraffin oil, and cultured at 37°C in an atmosphere of 5% CO2 in air with 100% humidity until use.
Cumulus cells were removed from the hyaluronidase drops and placed in an Eppendorf tube with H-CZB medium to be washed by centrifugation at 250 × g. The cumulus cell pellet was then resuspended in a small volume of 3% polyvinylpyrrolidone in H-CZB and kept on ice until the moment of the nuclear injection.
Production of cloned embryos by SCNT
The method for nuclear transfer referred to the previous description (Wakayama et al., 1998). Our self-made beveled injection needle was used for easier operation. Enucleated oocytes were injected individually with a cumulus cell nucleus. After nuclear transfer, the reconstructed oocytes were activated using 10 mM SrCl2 in Ca2+-free CZB medium in the presence of 5 μg/mL cytochalasin B (whether supplemented with 50 nM PXD101 depending on the experimental procedure). After three washes in KSOM, cloned embryos were cultured in the same medium for development. Usually, one droplet (10–15 μL) contained about 20 embryos and was cultured in KSOM medium until the end of the experiment.
At the end of the culture, embryos that reached the blastocyst stage at 96 hours were washed three times in phosphate-buffered saline (PBS) and placed onto slides in a drop of mounting medium (mixture of glycerol and PBS [9:1] containing 25 μg/mL of Hoechst 33342) (Jin et al., 2015). A coverslip was placed on top of the blastocysts, the edge was sealed with nail polish, and the number of nuclei was counted under ultraviolet light.
Aggregation of embryos
Our previous study about the effect of different pronase treatments and different aggregation systems on embryo aggregation indicated that aggregation of the embryos treated by 0.25% pronase for 10 minutes in our self-made concave hole system had the highest aggregation rate, development potential, and blastulation rate (Qiu et al., 2017a). Therefore, on day 2 after in vitro cultivation, the zonae pellucidae (ZPs) of four-/eight-cell cloned embryos were removed by 0.25% pronase for 10 minutes. Aggregation was accomplished by placing 2× cloned embryos (4-/8-cell) with the ZP free within each of our self-made concave hole and cultured to the blastocyst stage.
Isolation of NT-ESCs
Cloned embryos were produced as described above and, when they reached the blastocyst stage, they were used to isolate the NT-ESCs as previously described (Wakayama et al., 2001; Wang et al., 2007). When the inner cell masses (ICMs) were growing normally and had been passaged five to six times, they formed ESC-like colonies. The karyotype analysis and teratoma experiments were used to evaluate the NT-ESCs. NT-ESCs were injected subcutaneously into 4-week-old immunodeficient mice to test for teratoma formation. Four weeks after injection, the mice were sacrificed and the resulting teratomas were excised, fixed in 4% paraformaldehyde, embedded in paraffin, sectioned, and stained for histological examination.
The real-time polymerase chain reaction (PCR) method below was used to detect the expression of the pluripotent genes (POU5F1, NANOG, CDX-2, and REX-1) of isolated NT-ESCs.
Total RNA extraction, reverse transcription-polymerase chain reaction (RT-PCR), and real-time PCR
Total RNA was isolated from previously frozen (−80°C) NT-ESCs using the easy-spin™ (DNA free) Total RNA Extraction Kit according to the manufacturer's instructions. cDNA was produced from 1 μg of total RNA from NT-ESCs samples using the Omniscript RT Kit (Qiagen, Hilden, Germany). Quantitative RT-PCR (qRT-PCR) was performed using a LightCycler 2.0 (Roche, Germany). PCR products for four genes (POU5F1, NANOG, CDX-2, and REX-1) were detected in three replicates by SYBR Green probes, and each reaction consisted of 2 μL of cDNA, 2 μL of SYBR Green I Master Mix (Toyobo, Japan), 2 μL each of the forward and reverse primers, and was adjusted to a total volume of 20 μL using distilled water.
The amplification conditions consisted of initial preincubation (95°C for 30 seconds) for hot-start DNA polymerase activity, followed by 45 amplification cycles at 95°C for 5 seconds, 60°C for 20 seconds, and 72°C for 20 seconds. The fluorescence values were collected after each elongation step to determine the threshold cycle and the cycle during the log linear phase of the reaction at which fluorescence increased above background. At the end of the amplification cycle, a melting curve analysis was performed to determine the specificity of amplification. The melting protocol included holding at 60°C for 15 seconds and then heating from 60°C to 95°C. The fluorescence values were collected during the whole procedure with a temperature ramp rate at 20°C/min.
Primer sequences and the approximate sizes of the amplified fragments of all transcripts (POU5F1, NANOG, CDX-2, and REX-1) are shown in Table 1. Each transcript sample was quantified in three replicates and their relative expressions were analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Statistical analyses
Each experiment was repeated three times. Data (%) were analyzed using the Chi-square test. The number of nuclei was analyzed using an analysis of variance. Statistical analyses were performed using SPSS 16.0 software (SPSS, Inc., Chicago, IL). p Values <0.05 were regarded as statistically significant.
Experimental Design
Investigation of the effect of four-/eight-cell aggregation on the development of SCNT embryos
We evaluated the effect of SCNT embryo aggregation at the four- and eight-cell stages. The rate of aggregated embryo (evaluation: no separation within 0.5 hours after aggregation) and the rate of blastocyst formation were assessed.
Investigation of the effect of PXD101 and embryo aggregation on the development of SCNT embryos and on the establishment of NT-ESCs
Our previous study (Qiu et al., 2017b) had investigated the effect of PXD101 on mouse SCNT embryos and concluded that the treatment of 50 nM PXD101 for 10 hours after activation could most effectively improve the in vitro developmental capacity of mouse SCNT embryos. Therefore, the treatment of 50 nM PXD101 for 10 hours after activation was used in this study for the comparison of its effect with embryo aggregation treatment and also their combinations on mouse SCNT embryo development and on the establishment of NT-ESCs. The blastocyst formation rate, cell number of the blastocyst, and rate of NT-ESCs established were compared among aggregation or PXD101-treated groups or their combinations or untreated (control) groups.
Investigation of the effects of PXD101 and embryo aggregation on the pluripotent gene expression of established NT-ESCs
The expression of pluripotent genes (POU5F1, NANOG, CDX-2, REX-1) were detected using real-time PCR. The relative expression levels of these genes were compared among aggregation or PXD101-treated groups or their combinations or untreated (control) NT-ESCs groups. Each transcript sample was quantified in three replicates and their relative expressions were analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Results
Investigation of the effect of four-/eight-cell aggregation on the development of SCNT embryos
There is no significant difference between aggregation at four-cell stage and at eight-cell stage for the rate of aggregation and the rate of blastocyst in our self-made concave hole aggregation system (Table 2). Therefore, the aggregation at four-cell stage was chosen for the following effect in comparison with the PXD101 treatment.
Different lowercase superscripts in the same column indicate difference at p < 0.05.
Investigation of the effect of PXD101 and embryo aggregation on the development of SCNT embryos and on the establishment of NT-ESCs
The NT-ESCs have a smooth surface and obvious boundary with the surrounding MEF cells and exhibit morphology typical of undifferentiated ESCs (Fig. 1A). The karyotype analysis showed that the observed NT-ESCs had a normal diploid karyotype (Fig. 1B). In addition, 4 weeks after injection of NT-ESCs into 4-week-old immunodeficient mice, the teratoma (2 cm in size) was formed (Fig. 1C), and histology showed nerve (epiblastic), fat (mesoblastic), and gland (endoblastic) (Fig. 1D–F), further demonstrating the pluripotency of the isolated NT-ESCs.
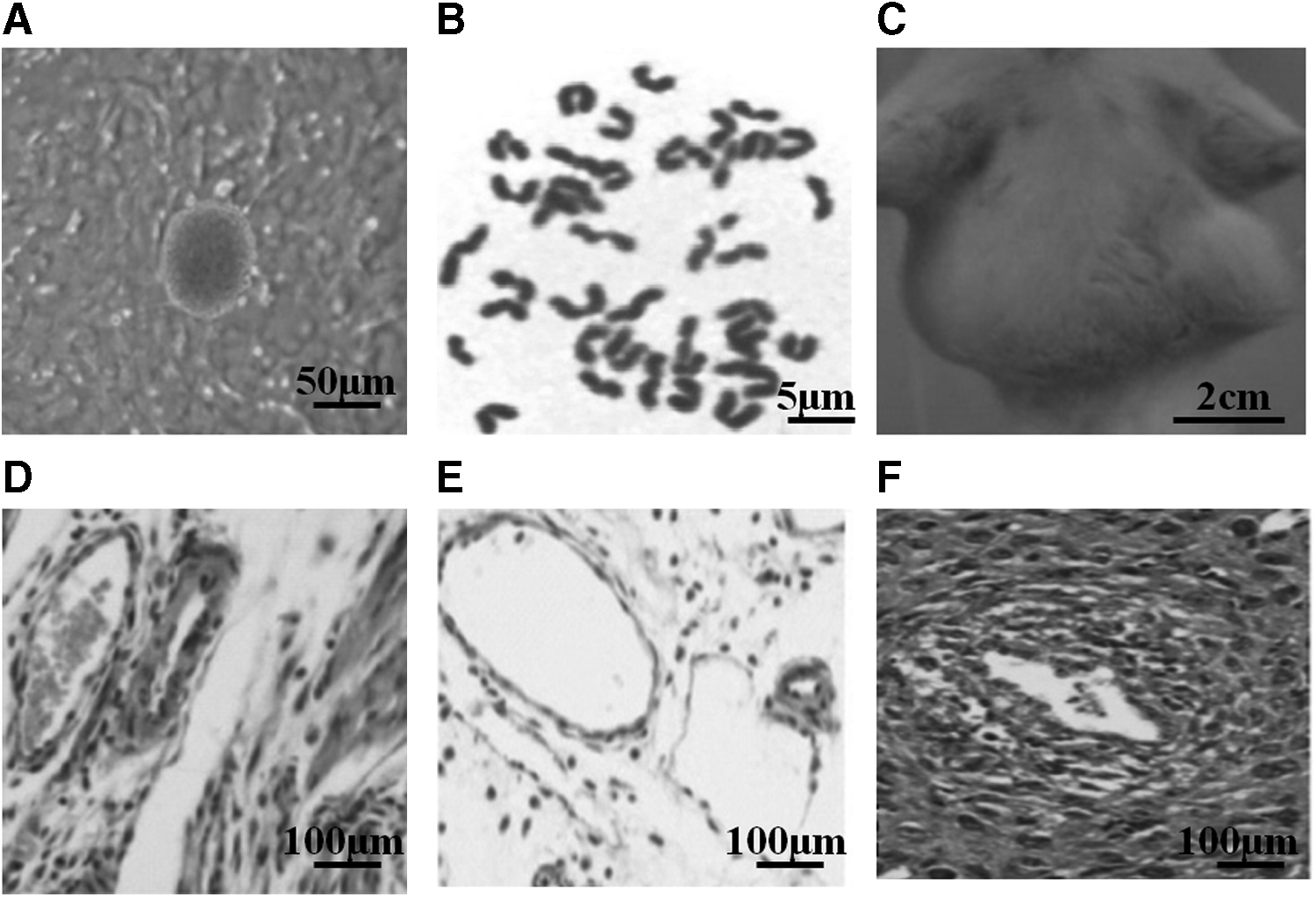
Evaluation of NT-ESC.
There is an effect of PXD101 treatment and embryo aggregation on the development of mouse SCNT embryos and on the isolation of NT-ESCs.
Compared with the control (untreated SCNT embryo), treatment with 50 nM PXD101 for 10 hours (during [6 hours] + after [4 hours] activation) and four-cell SCNT embryo aggregation could significantly improve the blastocyst formation, the total cell number per each blastocyst, and the subsequent establishment of NT-ESCs (p < 0.05; Table 3), but there was no significant difference between both (p > 0.05; Table 3). However, the treatment of their combination (50 nM PXD101 treated during [6 hours] + after [4 hours] activation first, and then aggregated at four-cell stage) has a higher rate of blastocyst formation, although not significant, but significantly more total cell number of each blastocyst and significantly higher rate of the subsequent establishment of NT-ESCs than that by their separate treatment (p < 0.05; Table 3).
Different lowercase superscripts in the same column indicate difference at p < 0.05.
50 nM PXD101 treated 10 hours first and then aggregated at four-cell stage.
NT-ESC, nuclear transfer embryonic stem cell; PN, pronucleus.
Investigation of the effects of PXD101 and embryo aggregation on the pluripotent gene expression of established NT-ESCs
There is an effect of different treatments on the pluripotent gene expression of established NT-ESCs. The relative expression levels of pluripotency-related genes (POU5F1, NANOG, CDX-2, and REX-1) were similar between PXD101 and aggregation-treated groups, but both were significantly higher than the untreated (control) group (Fig. 2), suggesting that PXD101 and aggregation have a similar effect on the increase of the rates of establishing NT-ESCs (Fig. 2 and Table 3). However, for the combination of PXD101 and aggregation, although the relative expression levels of CDX-2 and REX-1 were similar with their separately treated group, the expression of genes POU5F1 and NANOG was significantly higher than their separate treatment (Fig. 2). It was concluded that the combination of PXD101 and aggregation treatment has the highest expression level of pluripotency-related genes (POU5F1, NANOG, CDX-2, and REX-1) of established NT-ESCs.
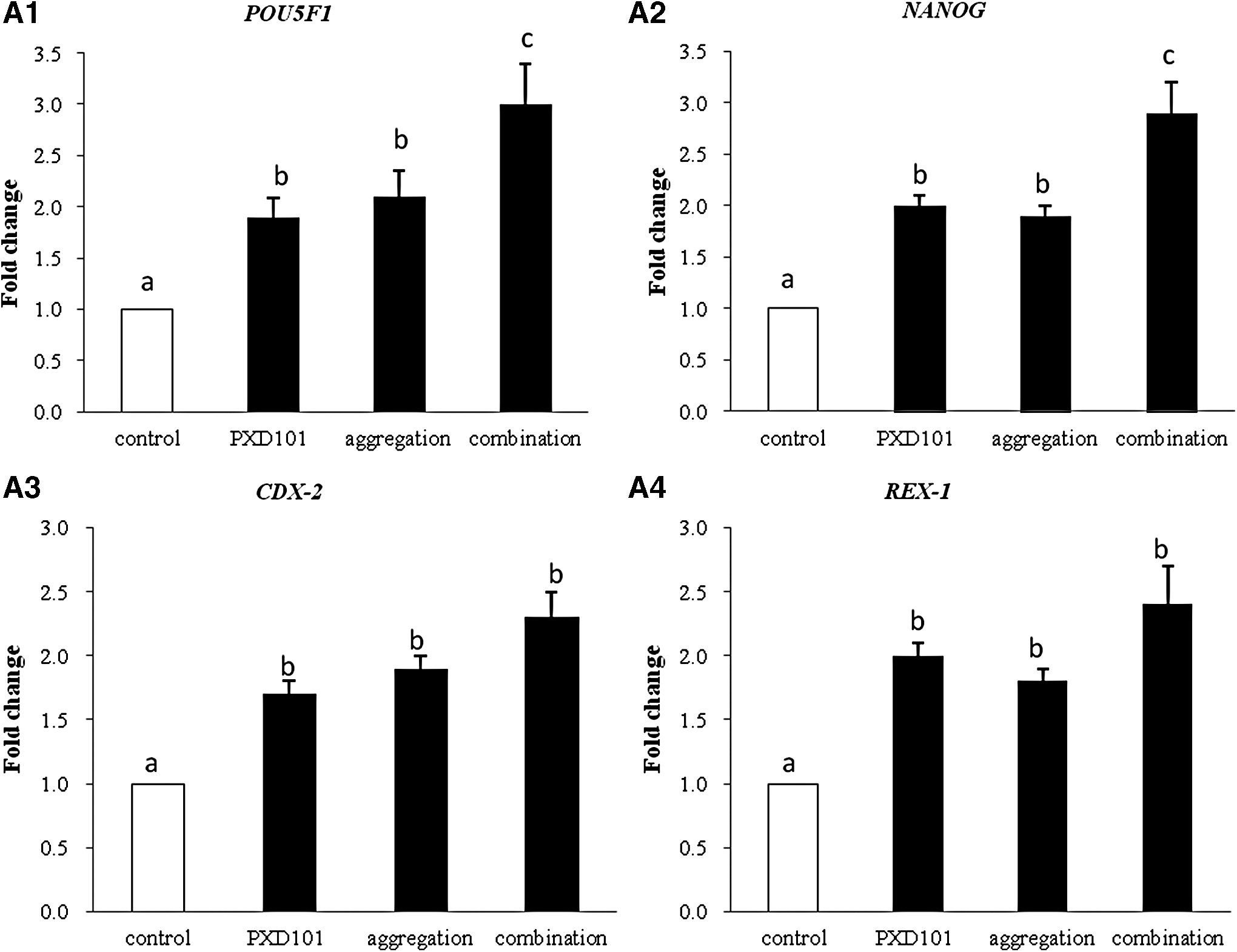
Discussion
To optimize the method to increase the in vitro developmental competence of mouse SCNT blastocysts, the present study compared the effects of embryo aggregation method and application of HDACis (PXD101) on the development potential of mouse SCNT embryos and on subsequent NT-ESCs establishment. The results suggest that both methods have a similar effect on the improvement of SCNT embryo development; however, the combination of their treatments (50 nM PXD101 treated for 10 hours during and after activation and then aggregation of the four-cell embryos) had the best development rate and subsequent NT-ESC isolation rate and the pluripotency-related gene expression.
Studies suggested that there are so far two main methods to improve the in vitro development of blastocysts: embryo aggregation and application of HDACis (Lee et al., 2006, 2007; Qiu et al., 2016). However, most of the recent studies about the effect of embryo aggregation on the in vitro developmental competence of blastocysts, including IVF embryo, SCNT embryo, and parthenogenetic embryo, were carried out in pigs (Lee et al., 2006, 2007, 2013; Saadeldin et al., 2014; Siriboon et al., 2014; Terashita et al., 2011). Our previous study in mice (Qiu et al., 2017a) indicated that the embryo aggregation improves the development of parthenogenetic blastocyst and pESCs derivation, and we also investigated the effect of a novel HDACi-PXD101 on in vitro developmental capacity of mouse SCNT embryos (Qiu et al., 2017b), but there is limited study about the comparison of these two methods on mouse SCNT embryo, therefore, we carried out this study for method optimization.
In the present study, we concluded that embryo aggregation improved the in vitro development potential of mouse embryos (Table 3), which is consistent with previous studies in cattle (Wells and Powell, 2000), mice (Tang and West, 2000), rhesus monkey (Schramm and Paprocki, 2004), and pigs (Lee et al., 2006, 2007; Lee et al., 2013; Saadeldin et al., 2014; Siriboon et al., 2014; Terashita et al., 2011). In addition, we generated an increased blastocyst formation rate (40.2%) by 2× embryo aggregation (Table 3), which is similar to the result (40%) by Terashita et al. (2011), but lower than that by Siriboon et al. (2014) (63.0%), which used a different system (the well-of-the-well system). The blastocyst formation rate was 40.0% by 50 nM PXD101 treated during (6 hours) + after (4 hours) activation (Table 3), which is also similar with our previous study (40.2%) (Qiu et al., 2017b).
In addition, we suggested that aggregation at four-cell stage is better for the rate of aggregation, rate of development, and rate of blastocyst, although there is no significant difference with that at eight-cell stage, which is also consistent with the previous studies in pigs (Lee et al., 2007; Siriboon et al., 2014; Terashita et al., 2011), where it was demonstrated that 4-cell stage embryos are the best stage for aggregation, whereas those at the 2-cell or beyond the 16-cell stage have a low aggregation or development rate (Lee et al., 2007). It is suggested that embryo aggregation could enhance their physical contact and interactions between the embryos.
The intercellular communication between blastomeres is demonstrated to be through gap junction, which is a prerequisite for the embryo development (Lee et al., 2007; Siriboon et al., 2014). The gap junctions were first apparent at the eight-cell stage, and increased in number from compaction onward in the mammalian embryo (Goodall and Johnson, 1984; Lo and Gilula, 1979). Therefore, it is conceivable that the intercellular communication between aggregated embryos can occur subsequent to aggregation—as early as the four- and eight-cell stages, at which point the effect becomes apparent (Boiani et al., 2003; Lee et al., 2007). Therefore, the cell and cell interactions through gap junctions may overcome the possible defect in vitro, which may be the possible reason why aggregation of the four-cell stage embryos could enhance the development potential of embryos.
It was observed from this study that compared with the control untreated embryo, the total cell number of blastocyst was significantly increased by two methods (aggregation and PXD101) (Table 3), but it may be attributed to different mechanisms involved.
It is understandable that the embryo aggregation could result in the increase of cell number of blastocyst as well as the intercellular communication. As it was stated, the formation of intercellular gap junctions between blastomeres, which allows electrical and biochemical coupling, such as the ions (e.g., K+ and Ca2+), intracellular signaling molecules (e.g., cAMP and cGMP), and small metabolite (e.g., glucose) exchanges (Lawrence et al., 1978; Siriboon et al., 2014), is critical for normal development during the morula stage, therefore, the enhanced intercellular communication resulted by aggregation facilitated the in vitro development of mouse SCNT embryos.
While for the HDACi-PXD101 in the present study, it also increased the total cell number of embryos (Table 3). In fact, increased histone acetylation due to the function of the HDACi, has been associated with a more effective formation of DNA replication complexes (Costa-Borges et al., 2010; de Ruijter et al., 2003), which can facilitate cell proliferation and could explain the results in the present study.
In this study, it was found that embryo aggregation and PXD101 treatment after SCNT both significantly improved the subsequent establishment of NT-ESCs and also the expression levels of pluripotency-related genes (POU5F1, NANOG, CDX-2, and REX-1) in comparison to the untreated group (Table 3 and Fig. 2). Nowadays NT-ESCs are attracting more public attention due to their application in regenerative medicine (Hwang et al., 2004, 2005). However, there is a limit for its application due to its low establishing rates. Our study here suggests that a new technique of PXD101 treatment and embryo aggregation can be applied to improve the rates when establishing NT-ESCs.
Our present study concluded that the embryo aggregation at four-cell stage and application of HDACi (PXD101) (50 nM; treated for 10 hours after SCNT) could both improve the in vitro development potential of mouse SCNT embryos and subsequent NT-ESCs establishment, whereas their combination had the best effect, therefore, the combination of these two methods is considered as the optimal treatment for the in vitro development of SCNT embryo in mice, providing a new approach for the practical improvement of mouse cloning techniques and opening new opportunities to improve cloning efficiencies in other mouse strains or species. Future study should be focused on the health of the combination treated cloned mice and the molecular mechanism underlying its improvement.
Conclusion
The combination of embryo aggregation and application of PXD101 treatment (50 nM PXD101 treated for 10 hours after SCNT and then aggregated at four-cell stage) is considered as the optimal way to improve the in vitro development of SCNT embryo and subsequent NT-ESCs isolation in mice.
Footnotes
Acknowledgments
This study was supported by the Scientific Research Funds (PhD Foundation) of Southwest University (grant no. SWU116037), the Fundamental Research Fund for the Central Universities (grant no. XDJK2017C011), and the National Natural Science Foundation of China (grant no. 31572488).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.