
Abstract
Abstract
The H19/insulin-like growth factor 2 (Igf2) gene plays vital roles during development, but their expression and methylation status have not been elucidated in male cloned animals. In this study, we investigated the methylation status of imprinting control region (ICR) and differentially methylated region (DMR) of the H19/Igf2 locus in the sperm and testes of naturally reproduced Saanen dairy goats (NG, control), transgenic cloned dairy goats (CG), and the first generation of mating the CGs with the female NGs (FG). Bisulfite sequencing polymerase chain reaction revealed that the H19 ICR was highly methylated in the sperm of all testing groups (97.07% ± 0.47%, 93.73% ± 0.85%, and 96.67% ± 0.54%). Notably, the methylation level of H19 ICR was decreased in the testes of CG goats compared with that of NG goats (73.83% ± 3.28% vs. 97.27% ± 0.71%, p < 0.05), while the Igf2 DMR2 was highly methylated in the sperm of these three groups (89.57% ± 1.74%, 87.23% ± 1.11%, and 83.2% ± 1.27%). However, the Igf2 DMR2 was abnormally hypomethylated in the testes of CG goats (27.33% ± 1.005%, p < 0.001), and moderately methylated in FG goats (53.1% ± 1.47%, p < 0.01) compared with that in NG goats (82.67% ± 0.70%). Furthermore, we found significantly decreased expression of H19 (p < 0.01) and increased expression of Igf2 (p < 0.01) in CG goats. Thus, we infer that somatic cell nuclear transfer has clear effect on genomic imprinting in the testes, but not sperm of cloned goats.
Introduction
G
Somatic cell nuclear transfer (SCNT) permits transformation of a terminally differentiated somatic cell into an undifferentiated zygote with the potential to develop into a newborn animal, thus it holds great potential for applications in agriculture, endangered species conservation, and regenerative medicine (Jaenisch and Young 2008). However, abnormal organs size, respiratory issues, and perinatal death have been described in cloned conceptuses and neonates. Moreover, these pathologies are associated with aberrant DNA methylation and genomic imprinting (Cibelli et al., 2002; Farin et al., 2006; Ogura et al., 2013; Yang et al., 2007).
The H19/insulin-like growth factor 2 (Igf2) locus is one of the most widely studied imprinting clusters, its expression and methylation status have been reported in a number of cloned animals, including mice, cattle, piglets, and goat (Meng et al., 2013; Ogawa et al., 2003; Su et al., 2011; Wei et al., 2010). In both mouse and human genomes, H19 is located adjacent to the paternally expressed Igf2 gene and contains one ICR, located 2 kb upstream of the H19 transcription start site (Barlow 2011; Ideraabdullah et al., 2014). This multifunctional ICR plays roles in insulation, activation, and repression and contains binding sites for the CCCTC-binding factor (CTCF), which binds to the un-methylated allele (Phillips and Corces 2009).
The Igf2 contains two DMRs. DMR1 is a silencer that is inactivated by methylation, whereas DMR2 is an enhancer that is activated by methylation (Constancia et al., 2000; Murrell et al., 2001). On the paternal allele, the methylated H19 ICR inhibits CTCF binding, enabling enhancers to access the Igf2 promoter on the paternal chromosome. On the maternal allele, the ICR is un-methylated. When bound to the maternal H19 ICR, CTCF insulates maternal Igf2 DMR1 and DMR2, preventing methylation, and thus, silencing the Igf2 maternal allele (Lopes et al., 2003).
Because SCNT is often used for the production of specific human proteins in the milk of transgenic animals, consequently, analyses of the H19/Igf2 locus have primarily focused on tissues of female cloned animals. However, the expression and DNA methylation status of the H19/Igf2 locus in male cloned animals has yet to be elucidated. In this study, we investigated the methylation and expression of H19/Igf2 in the sperm and testes of cloned goats, to determine the epigenetic status of imprinted genes in the reproductive system of male cloned goats and their offspring.
Materials and Methods
Experimental design
The present investigation was conducted on normal Saanen dairy goats (NG), transgenic cloned dairy goats (CG), and the first generation of mating CGs and female NGs (FGs) (Fig. 1A). Three bucks each from the three experimental breeds were selected for collection of semen and testes. The age, reproductive history, and recent status of these bucks used in this study were listed in Table 1.

Schematic representation of the experiment design and target DNA sequence of the H19/Igf2 DMR.
Goats were provided ad libitum access to clean drinking water and seasonal green fodder depending on the availability along with the mixture of maize and oat silage. Vaccination, deworming, regular checkup for communicable diseases, and other herd-health programs were followed as per standard farm schedule, to protect the animals from diseases and produce quality semen.
All the experimental procedures involving animals were in accordance with the National Research Council's publication “Guide for the Care and Use of Laboratory Animals” and approved by Institutional Animal Care and Use Committees at Nanjing Agricultural University.
Production of transgenic cloned goats
The transgenic cloned dairy goats were produced by SCNT, which has been previously described (Meng et al., 2013; Wan et al., 2012). Briefly, metaphase II (MII)-stage oocytes were enucleated by standard micromanipulation techniques. Cycle-synchronized donor fibroblast cells were placed between the zona and cytoplast membrane of the enucleated oocytes by using a 20 μm (inner diameter) needle.
Incorporation of the donor cell nuclei into the cytoplasm of the enucleated oocytes was accomplished by exposing cytoplast/karyoplast couplets to two 1.2 kV/cm DC pulses lasting for 20 μsec in the presence of a calcium-rich fusion medium [0.25 M d-sorbitol, 0.5 mM (CH3COO)2MgN4H2O, 0.1 mM (CH3COO)2Ca, 0.5 mM HEPES, and 1 mg/mL bovine serum albumin; pH 7.3]. Reconstructed embryos were activated by treating with 5 mM ionomycin for 5 minutes, followed by incubation in 2 mM 6-DMAP for 4 hours, and culturing in M16 Media (Sigma) for 8 hours. Then, two-cell SCNT embryos were surgically implanted (approximately eight embryos per goat) into the oviducts of recipient local goats.
Semen collection
Ejaculates were collected from each buck twice a week as per the routine of the semen station. A total of four ejaculations from each goat were collected, after examination of sperm motility and concentration (Table 2), one around the average was selected for the tests. Thus, a total of nine ejaculations were used. After collection, the samples were immediately brought back to the lab and kept in the water bath at 37°C for density gradient centrifugation.
Sperm gradient
To remove dead sperm and antibodies in the seminal plasma and acquire high quality sperm, Percoll gradient was performed as previously described (Claassens et al., 1998), with slight modification. Briefly, a stork isotonic Percoll solution was prepared by adding 1.0 mL of 10 × concentrated phosphate-buffered saline (PBS; Solarbio, Beijing, China) to 9.0 mL of Percoll (Pharmacia AB, Uppsala, Sweden). The bottom fraction of the Percoll gradient (90%) was mixed by 9.0 mL of the Percoll stock solution and 1.0 mL of 1 × concentrated PBS. The middle fraction (45%) consisted of 4.5 mL of Percoll stock solution mixed with 5.5 mL of 1 × concentrated PBS. Solutions were stored at 4°C until used.
To prepare the gradient, 800 μL of 90% Percoll was pipetted into the bottom of the centrifuge tube, 800 μL of 45% Percoll was carefully layered over the bottom fraction and 200 μL of semen was layered over the middle fraction. The tubes were centrifuged at 400 × g for 25 minutes at room temperature. After the supernatant was discarded, samples were washed twice in 1 × concentrated PBS and resuspended in 50 μL of Sample Protector for RNA/DNA (TaKaRa, Dalian, China) and stored at −80°C for further use.
Tissue collection
Tissues were collected immediately after castration of CG, NG, and FG goats and were excised into slice, and stored at −80°C in Sample Protector for RNA/DNA (TaKaRa) until further use. Transgenic goats were identified by polymerase chain reaction (PCR) screening and southern blotting (unpublished data).
DNA isolation
Sperm and testes genomic DNA was isolated from each sample using the DNeasy Blood & Tissue Kit (No. 69504; Qiagen, Hilden, Germany) following the manufacturer's instruction, with the exception that DNA was eluted in 20 μL elution. Concentration of DNA yielded from the sperm and testes was determined using a Nanodrop Spectrophotometer (Thermo Scientific, Waltham, MA). Sample was deemed acceptable if 260/280 ratios were >1.8 and 260/230 ratios were >1.0.
DNA methylation status analysis
Putative DMR identification and primers design
The DMRs of imprinted genes have highly conserved regions across various mammal species. To identify the counterparts of these DMRs in goat genome, the well-known DMRs of H19 and Igf2 in human, bovine, and ovine genomes were compared and the distribution of CpG islands was identified using an online software MethyPrimer with restrictive conditions (island size >100, GC percentage >50.0%, CpG observe/expect >0.60) (Fig. 1B). The specific primers for amplification and sequencing of DMRs of H19 and Igf2 are listed in Table 3.
R, A/G.
Igf2, insulin-like growth factor 2.
Bisulfite treatment of oligonucleotides
Genomic DNA was treated with sodium bisulfite using the EZ DNA Methylation-Direct Kit according to the manufacturer's recommendation (Zymo Research Corp., Orange, CA). Briefly, 500 ng DNA in 20 μL volume was added to 130 μL of CT conversion reagent solution in a PCR tube. Bisulfite conversion was performed on a thermo cycler as follows: 98°C for 8 minutes, 64°C for 3.5 hours, and 4°C storage for up to 20 hours. The bisulfite-treated DNA was recovered by Zymo-Spin™ IC Column and subsequently sequenced to confirm the efficiency of bisulfite conversion. Only the DNA with more than 99% cytosine conversions was used for bisulfite PCR and sequencing.
Bisulfite PCR and sequencing
For the amplification of the H19 and Igf2 putative DMRs, touch down PCR was performed using 20 ng of the bisulfite-converted genomic DNA as a template. The PCR procedure was as follows: one cycle of 95°C for 5 minutes; two cycles of 95°C for 30 seconds, 64°C (annealing temperature) for 1 minute, and 72°C for 1 minute; two cycles of 95°C for 30 seconds, 62°C for 1 minute, and 72°C for 1 minute and so forth; 20 cycles of 95°C for 30 seconds, 54°C (annealing temperature) for 1 minute, and 72°C for 1 minute, and a final cycle of 72°C for 7 minutes. Individual PCR products were purified with a Gel Extraction Kit, and subsequently cloned into the pMD19-T vector (TaKaRa).
Plasmid DNA was isolated using AxyPrep Plasmid Miniprep Kit (Axygen Biosciences, Union City, CA) and sequenced by Big Dye (ABI) on an ABI PRISM Model 3730. The DNA methylation patterns derived from more than 10 clones with 99% cytosine conversions were analyzed using the BIQ Analyzer software.
Gene expression analysis
Total RNA was extracted from the goat testes and sperm using the RNAprep Pure Tissue Kit (Tiangen) and TRIzol reagent, respectively. Reverse transcription (RT) was performed using the PrimeScript RT reagent kit with gDNA Eraser (TaKaRa) according to the manufacturer's protocol.
Quantitative real-time PCR (qPCR) was performed on an ABI 7300 Real-Time PCR System (Applied Biosystems). The PCR reaction consisted of 10 μL of SYBR Green PCR Master Mix, 10 pM of forward and reverse primers, and 1.0 μL template complementary DNA (cDNA), in a total volume of 20 μL. The PCR conditions were as follows: initial denaturation at 95°C for 10 minutes, followed by 40 cycles at 95°C for 15 seconds, annealing at 60°C for 30 seconds, and extension at 72°C for 30 seconds. A melting curve was performed to confirm the identity of each PCR product. Expression was normalized to Gapdh as a housekeeping gene. A complete list of primers used in this study is described in Table 4. The relative amount of each transcript gene that presented in each cDNA sample was calculated using the 2−ΔΔCT method.
Statistical analysis
All qPCR reactions were performed in triplicate. Data are presented as mean ± standard error of the mean. DNA methylation status of goat sperm and testes, and the gene expression were tested by one-way analyses of variance and multiple comparison between the groups was performed using S-N-K method (SPSS, Inc., Chicago, IL). A difference with p-value of <0.05 was considered statistically significant.
Results
DNA methylation status of the H19 ICR
The promoter and DMR of H19 (GenBank accession No. EF577239.1) were predicted, and this putative DMR was deemed an ICR based on its position (Fig. 1B). The putative ICR was located upstream of the promoter and consisted of 363 nucleotides and 1 CTCF (zinc-finger protein putative binding site) motif (CCGNNGGNGGC). The seventh and eighth CpG sites were located within the CTCF motif (Fig. 1B). BLAST search indicated that this ICR shared 99% identity with that of Ovis aries (GenBank accession No. AJ566210.1) and 88% identity with the Bos indicus breed Sahiwal H19 upstream ICR (GenBank accession No. GU371442.1).
As expected, the putative ICR was highly methylated in the sperm of NGs (93.73% ± 0.85%), CGs (97.07% ± 0.47%), and FGs (96.67% ± 0.54%), with no statistical difference (Fig. 2). Notably, the CTCF motif was completely methylated in the sperm of all three groups. Conversely, the 11th and 13th CpG sites in goat NG1, the 11th CpG site in goat NG2, and the 24th CpG site in goat NG3 were completely un-methylated (Fig. 2C).

Bisulfite sequencing analysis of the H19 ICR in the sperm.
In the testes of NG goats, the H19 ICR was almost completely methylated (97.27% ± 0.71%, Fig. 3B). Compared with the NG goats, the methylation level of H19 ICR was significantly lower in the testes of CG goats (73.83% ± 3.28%, p < 0.05; Fig. 3C, E) but showed no significant difference in the testes of FG group (91.57% ± 2.46%, p > 0.05; Fig. 3D, E). Notably, the CTCF motif was completely methylated in the testes of NG goats (Fig. 3B).

Bisulfite sequencing analysis of the H19 ICR in the testes.
DNA methylation status of the Igf2 DMR
The putative Igf2 DMR is located within the extron 4 (between nucleotides 7209 and 7663 nucleotides) and covers 22 CpG sites within 345 nucleotides (GenBank accession No. NC_030836.1) (Figs. 1B and 4A). This DMR exhibited 95% identity with that of Bos taurus (GenBank accession No. EU518675.1) and 97% identity with that of O. aries (GenBank accession No. CP011906.1). Based on its position, this DMR was designated as DMR2.
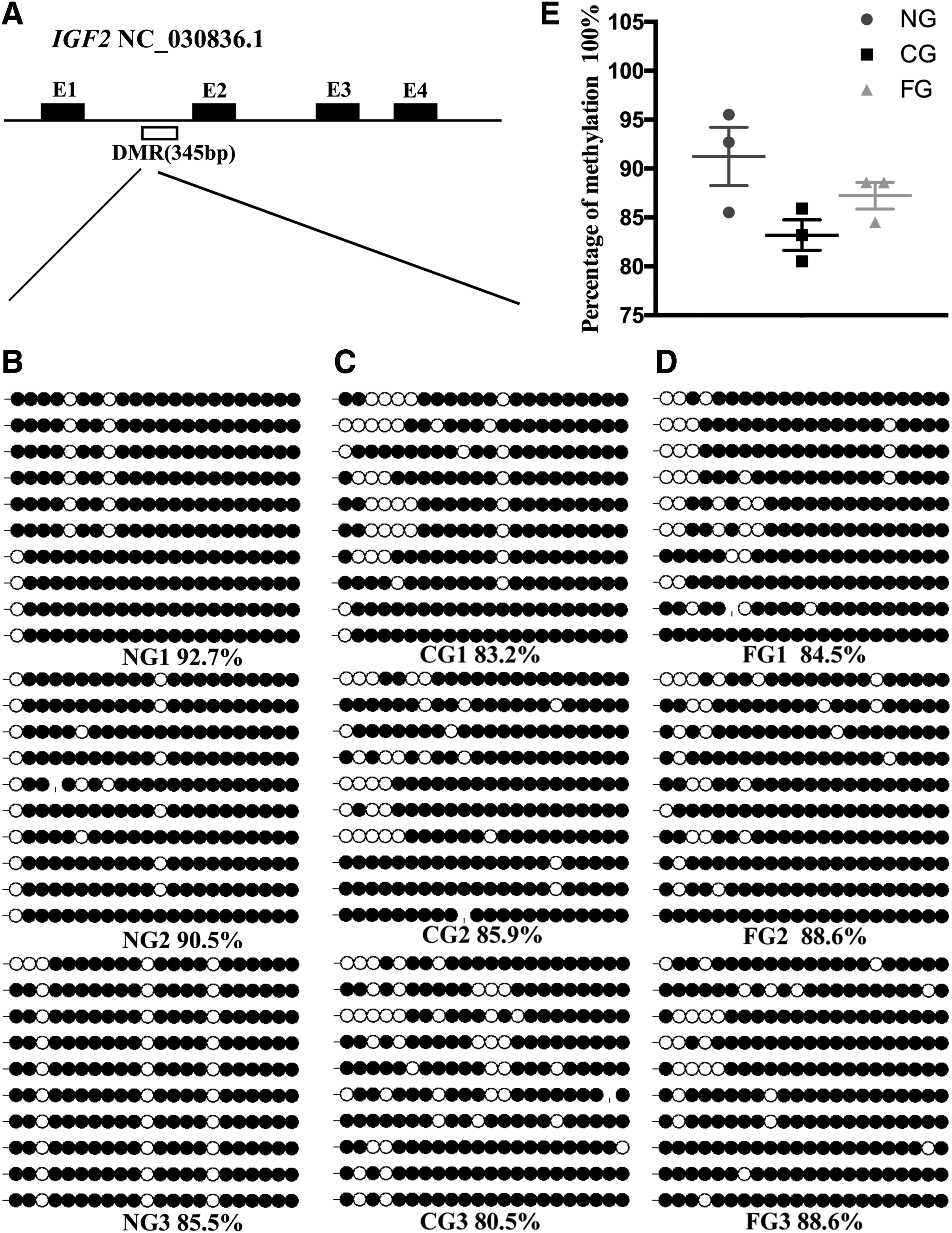
Bisulfite sequencing analysis of the Igf2 DMR2 in the sperm.
As shown in Figure 4, the DMR2 was hypermethylated in NG, CG, and FG sperm (89.57% ± 1.74%, 83.2% ± 1.27%, and 87.23% ± 1.11%), with no significant difference. Notably, the 3rd, 11th, and 16th CpG sites were completely un-methylated in NG goats.
In the testes, the Igf2 DMR2 were extensively methylated in the NG group (82.67% ± 0.70%), moderately methylated in the FG group (53.1% ± 1.47%), and hypomethylated in the CG group (27.33% ± 1.005%). Compared with the NG group, the methylation level was significantly lower in the CG and FG groups (p < 0.001, p < 0.01, Fig. 5; respectively). In addition, the percent methylation of Igf2 DMR2 in the CG group was also significantly lower compared with the FG group (p < 0.01; Fig. 5C–E). Moreover, the third and seventh CpG sites in CG1, the seventh and eighth CpG sites in CG2, the fourth CpG site in FG2, and the seventh CpG site in NG2 and NG3 were completely un-methylated (Fig. 5).

Bisulphite sequencing analysis of the Igf2 DMR2 in the testes.
Expression of imprinted genes
When studying nuclear reprogramming, it is important to examine the correlation between aberrant methylation with abnormal expression of imprinted genes. Here, we investigated two imprinted genes in the testes and sperm. As shown in Figure 6, compared to the NG goats, the expression of H19 was significantly decreased in both the sperm (p < 0.01) and testes (p < 0.01) of CG goats, whereas the expression of Igf2 was significantly increased (p < 0.01) in testes, but remained unaltered in the sperm of CG goats. In the sperm and testes of FG goats, the expression levels of these two genes were similar to that in NG goats.
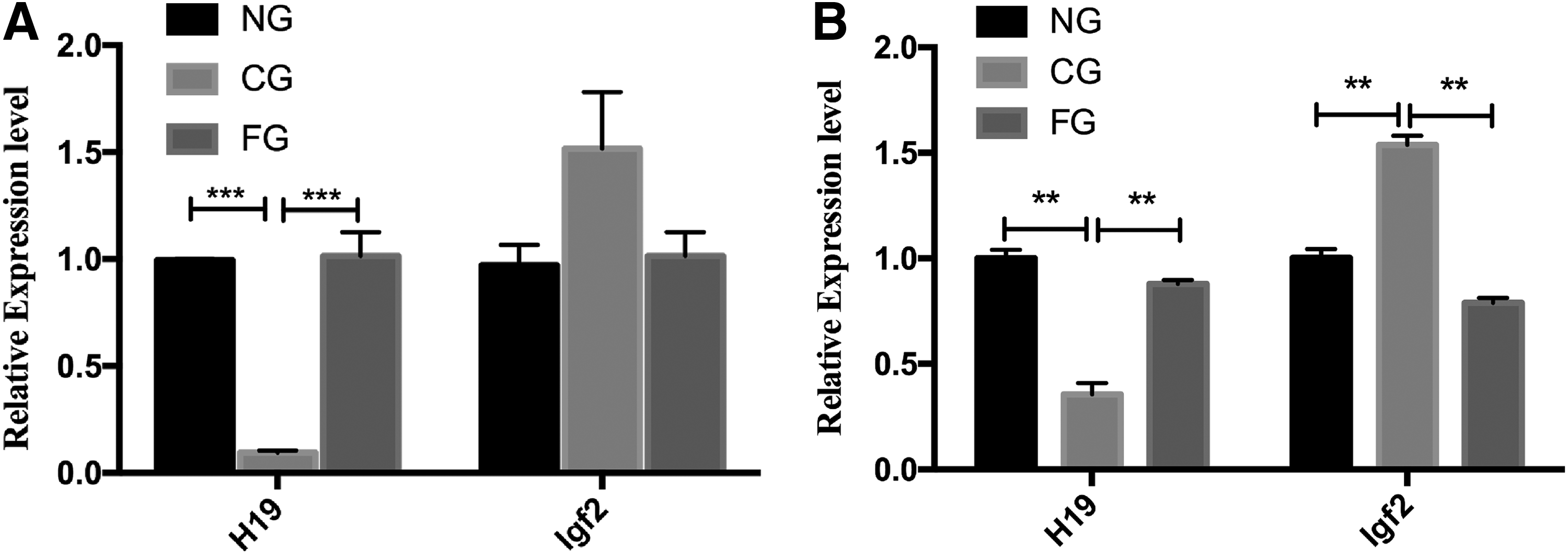
Quantitative analysis of H19/Igf2 in the sperm and testes of naturally reproduced goats, cloned goats, and their offspring.
Discussion
In the present study, we selected the H19/Igf2 locus as a representative for imprinted genes (Zhao et al., 2013) and their DNA methylation and expression were analyzed in the sperm and testes of cloned goats, to determine the epigenetic status of imprinted genes in the reproductive system of male cloned goats and their offspring.
Our data revealed that the expression of H19 and Igf2 in the testes of the first-generation progeny, from mating of male cloned goats with naturally reproduced female Saanen goats, was similar to that in naturally reproduced goats. However, the expression of H19 was downregulated in the sperm and testes of cloned Saanen dairy bucks and the expression of Igf2 was upregulated in the testes of cloned bucks, consistent with the result in the placentas of deceased cloned cattle (Su et al., 2011).
The correlation between aberrant methylation and abnormal expression of imprinted genes have been reported in cloned embryos and their subsequent offspring. Generally, increased DNA methylation in transcriptionally regulatory regions leads to a closed chromatin conformation that blocks access by transcription factors, resulting in downregulation of gene expression (Feldmann et al., 2013). Putative DMRs of goat Igf2 were deduced by alignment with the known bovine DMRs, although the putative DMR1 locus was very similar to the bovine locus, but was almost completely un-methylated in both parental alleles (Deng et al., 2017). Therefore, we sought to determine the methylation status of Igf2 DMR2.
The Igf2 DMR2 exhibited a differential methylation pattern in the porcine genome (Shen et al., 2012), and it was hypermethylated in the sperm of both cloned goats and their offspring in the present study, consistent with the results of a previous study involving porcine sperm (Han et al., 2008). These results suggest that genomic imprinting in sperm is normal. However, Igf2 DMR2 methylation was downregulated in the testes of cloned goats, whereas their offspring exhibited moderate methylation, at a level ∼30% less than that of control goats. In line with this, the expression of Igf2 was increased in the testes of cloned goats. Our data revealed that the Igf2 DMR2 exhibited abnormal methylation in the testes of cloned bucks, but this abnormality is moderated in the offspring.
The long non-coding RNA gene H19 is maternally expressed and associated with growth suppression. In naturally reproducing animals, the H19 ICR is only partially methylated in the tissues but highly methylated in sperm (Ibala-Romdhane et al., 2011; Meng et al., 2013; Su et al., 2011; Zhao et al., 2013). In the sperm of cloned pigs, the H19 ICR at the third CTCF sites was shown to be hypermethylated (Han et al., 2008), consistent with the results of this study involving goats.
The results of several bisulfite sequencing studies support the hypothesis that allele-specific DNA methylation of the H19 ICR is faithfully replicated during preimplantation development in mice (Inoue et al., 2002). Thus, the methylation status of the H19 ICR could reflect the reprogramming efficiency of cloned embryos. The results of previous studies, in conjunction with our present results, suggest that genomic imprinting might be normal in the sperm of live cloned goats.
In the present study, we observed decreased expression of the H19 gene in both the sperm and testes, consistent with our previous study (Meng et al., 2014). Inconsistently, DNA methylation of the H19 ICR in the testes of cloned goats was decreased. It was reported that the evolutionarily conserved CTCF plays an active role in H19 imprinting (Phillips and Corces 2009). The CTCF binds to the un-methylated H19 ICR, promoting H19 maternal allele (Ideraabdullah et al., 2008). Using the CCGNNGGNGGC motif, we found four putative CTCF sites in goat H19, a number equal to that in the sheep (Young et al., 2003).
Moreover, the H19 ICR at the third CTCF site was hypermethylated in the testes of all testing goats, thus, silencing the H19 paternal allele. However, the downregulation of H19 in cloned bucks still needs further investigation. One of the reasons is that relationship between methylation and gene expression is complex than previously thought and expression of the H19 gene might be epigenetically controlled at multiple levels (Zhou et al., 2015).
Notably, in the offspring of male cloned goats, the methylation status of the H19 ICR and the expression level of H19/Igf2 was similar to that of naturally reproduced goats. These data clearly indicate that methylation of specific imprinted DMRs is normal in the cloned goat offspring.
Conclusion
Although the H19/Igf2 locus is one of the most widely studied imprinting clusters, the identification of the uncharacterized DMRs of the H19 and Igf2 genes in male cloned goats and their offspring has not been established before. We found that methylation of the H19 ICR and Igf2 DMR2 in the sperm of live cloned goats and their offspring was similar to naturally reproduced goats, but was significantly lower in the testes of cloned bucks. Thus, we concluded that genomic imprinting might be normal in the sperm of cloned goats, but is abnormal in the testes of cloned goats and their offspring and this abnormality could be overcome by natural reproduction.
Footnotes
Acknowledgments
This study was financially supported by the National Natural Science Foundation of China (Nos. 31301973; 31272443) and by the National Major Special Projects on the Fundamental Research Funds for the Central Universities (No. KJQN201401).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist,