
Abstract
Abstract
p53 is a barrier to somatic cell reprogramming. Deletion or transient suppression of p53 increases the efficiency of reprogramming of somatic cells into induced pluripotent stem cells. Whether p53 represents an obstacle to a similar process transdifferentiation of somatic cells is unknown. However, it is predicted that inhibition of p53 would promote transdifferentiation of fibroblasts into cardiomyocytes. In this study, the effect of p53 on the capacity of cardiogenic transdifferentiation is evaluated using p53 wild-type (p53+/+), p53 heterozygous mutant (p53+/−), and p53 homozygous mutant (p53−/−) mouse embryonic fibroblasts (MEFs). Repression of p53 in MEFs increases the expression level of mesoderm transcription factors Brachyury (T) and MESP1. The cardiac-specific markers, Myh6 (Myosin, Heavy Chain 6), Myh7 (Myosin, Heavy Chain 7), and cTnI (cardiac muscle troponin I), show elevated expression in p53+/− and p53−/− MEFs compared with wild-type MEFs, but cardiac muscle troponin T (cTnT) showed a lower expression level when p53 was inhibited. After induction to cardiac differentiation, cTnT expression increased and markers of endoderm and ectoderm decreased in p53+/− and p53−/− MEFs. The effect of an important reprogramming factor Oct4 on cardiac transdifferentiation was also evaluated in the allelic series of p53 MEFs. We found that overexpression of Oct4 significantly enhanced Mesp1, Tbx5, and Isl1 expression in p53+/+ and p53+/− MEFs. Oct4 also enhanced cTnT expression in all three cell lines, especially in p53+/− MEFs. Thus, inhibition of p53 expression and viral expression of Oct4 both promote transdifferentiation of MEFs into cardiomyocytes, establishing reciprocity of action in the process.
Introduction
H
Both embryonic stem (ES) cells (Laflamme et al., 2007; Shiba et al., 2012; van Laake et al., 2007) and induced pluripotent stem (iPS) cells (Seki et al., 2014; So et al., 2011; Wang et al., 2016; Zhang et al., 2009) and other tissue-derived stem cells have been effectively differentiated into functional cardiomyocytes, and applied to improve heart function, but unfortunately, transplanting cells into an infarcted heart yields only transient and marginal benefits (Burridge et al., 2012).
Direct reprogramming of somatic cells into cardiomyocytes provides another strategy to generate cardiomyocytes. Recently, cardiomyocyte-like cells have been obtained from fibroblasts by introducing three lineage-specific transcription factors: gata4, mef2c, and tbx5 (Ieda et al., 2010). These cardiomyocyte-like cells displayed characteristics of cardiomyocytes, including gene expression profiles and electrophysiological function (Burridge et al., 2012). The expression ratio of gata4, mef2c, and tbx5 in the cells influenced the efficiency of cardiac transdifferentiation (Wang et al., 2015a, 2015b). Subsequently, it was shown that cardiomyocytes could be generated from cardiac fibroblasts, which predicts an approach for in situ transdifferentiation of resident cardiac fibroblasts into cardiomyocytes. However, the efficiency of the transdifferentiation process is low (Jiang et al., 2013), which will limit clinical application.
Efforts have been undertaken to increase the efficiency of transdifferentiation by including additional cardiac-specific factors, however, although improvement is observed, the rate of conversion is still low (Hirai et al., 2013). Clearly, additional approaches are required to overcome obstacles, likely to be epigenetic, to improve the efficiency of transdifferentiation. It has been demonstrated that inhibition of p53 expression significantly promoted the reprogramming of mouse and human somatic cells to iPS cells (Chu et al., 2015; Hong et al., 2009; Shi et al., 2015; Tidball et al., 2016; Wang et al., 2012). Studies have shown that up to 10% of transduced mouse embryonic fibroblasts (MEFs) lacking p53 were reprogrammed to iPS cells, even in the absence of Myc from the Yamanaka cocktail (Kawamura et al., 2009).
Subsequently, it was shown that p53 deletion promoted induction of integration-free mouse iPS cells generated by plasmid transfection (Rasmussen et al., 2014). The efficiency of reprogramming depends on the level of p53, which appears to act as a cell plasticity barrier (Hong et al., 2009). Oct4, a key transcription factor, plays a pivotal role not only in maintaining pluripotency and self-renewal of ES cells, but also plays emerging roles in somatic cell reprogramming (Di Stefano et al., 2009; Takahashi and Yamanaka, 2006; Yu et al., 2007). Oct4 alone, combined with small molecules, could induce cardiac differentiation of fibroblasts (Wang et al., 2014). Thus, Oct4 expression possibly promotes transdifferentiation efficiency.
Transdifferentiation of fibroblasts into cardiomyocytes has been successfully achieved, although with low efficiencies. This represents a bottleneck for therapeutic applications. Thus, strategies are required to overcome this limitation. We have taken a two-pronged approach to address the problem; one is to remove barriers to cellular plasticity (p53), and two to incorporate a degree of reprogramming into the process (Oct4).
In this study, p53+/+, p53+/−, and p53−/− MEFs were used as a model to investigate the effect of p53 on cardiac transdifferentiation. In addition, ectopic expression of Oct4 in p53+/+, p53+/−, and p53−/− MEFs was used to investigate the relationship between p53 and Oct4 on MEF transdifferentiation. Reciprocal actions between Oct4 and of p53 on MEF transdifferentiation toward cardiomyocytes were demonstrated in the process.
Materials and Methods
All surgical operations related to mice in the experiments were performed under anesthesia, and mice were sacrificed by euthanasia. These experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of Baylor College of Medicine and The University of Texas at Austin.
MEF isolation and maintenance
To prepare MEFs, embryos were harvested from mice of p53+/− C57BL6/129 heterozygous crosses at E13.5 followed by removal of head, four limbs, and gut. The remaining tissue was minced, then digested with trypsin, followed by resuspension of the cells in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) and 2 mM
Differentiation of MEFs toward cardiomyocytes
Transdifferentiation medium was composed of IMDM/M199 (4:1), 10% FBS, 5% horse serum 1% penicillin/streptomycin, 1 × glutamine, 1 × nonamino acid, 1 × sodium pyruvate (Invitrogen). MEFs were cultured in this medium and medium was changed every 2–3 days until cells were harvested.
Cell proliferation assay
1 × 105 MEF cells of the genotype p53+/+, p53+/−, p53−/− were plated in wells of six-well plates, and then cell numbers were counted every day using hemocytometer. Proliferation rate was evaluated by a growth curve.
RNA isolation and analysis
Total RNAs were isolated from the samples using TRIzol reagent (Cat. No. 15596018; Invitrogen). RNA samples were analyzed with quantitative real-time polymerase chain reaction (qRT-PCR) with SYBR Green Q-PCR reagent (Cat. No. 208056; Qiagen) and PCR primers listed in Table 1.
Afp, Alpha fetoprotein; CTnI, cardiac muscle troponin I; CTnT, cardiac muscle troponin T; Gapdh, Glyceraldehyde-3-phosphate dehydrogenase; Gfap, Glial fibrillary acidic protein; Myh6, Myosin, Heavy Chain 6; Myh7, Myosin, Heavy Chain 7.
Fluorescence-activated cell sorting
MEFs at day 21 of culture in transdifferentiation media were dispersed with 0.05% trypsin-EDTA in DMEM, and washed with Dulbecco's phosphate-buffered saline (DPBS) twice, and then stained with anti-cTnT (cardiac muscle troponin T) antibody (Cat. No. 8295; Abcam) diluted 1:200 as the primary antibody; normal mouse IgG as isotype control; and then stained with goat anti-Mouse IgG–Alexa Fluor® 647 (Cat. No. ab150115; Abcam) diluted 1:200 as secondary antibody. cTnT-positive cells were analyzed with the Flow Cytometer BD LSRFortessa (BD Biosciences).
Western blot analysis
Total cell lysates were subjected to western blot analyses. The following antibodies were used: primary antibodies against: cTnT (Cat. No. ab8295; Abcam), p53 (Cat. No. AF1355; R&D Systems), β-actin (Cat. No. A1978; Sigma), secondary antibodies: goat anti-mouse IgG-horseradish peroxidase (HRP, Cat. No. sc-2005; Santa Cruz), Rabbit anti-goat IgG-HRP (Cat. No. sc-2768; Santa Cruz). HRP activity was detected with a chemiluminescent methodology using the Pierce ECL Western Blotting Substrate Kit (Thermo Scientific). The emitted light was detected by photographic film.
Oct4 transfection
pMXs-based retroviral vectors-Oct4 (Cat. No. 13366; Addgene) were introduced into each 10 cm plate with Plat-E cells, respectively, using FuGENE 6 Transfection Reagent (Cat. No. 11814443001; Roche) according to the manufacturer's recommendations. One day before infection, MEFs were seeded at 8 × 105 cells per 10 cm gelatin (0.1%)-coated plate. After 24 hours of transfection, virus-containing supernatants derived from these Plat-E cultures were filtered through a 0.45 μm cellulose acetate filter and supplemented with 4 mg/mL polybrene. MEFs were incubated in the virus/polybrene-containing supernatants overnight. MEFs were infected twice. After 24 hours of infection, media were replaced with 10 mL fresh medium.
Statistical analysis
All data were presented as mean ± standard deviation. Student's t-test was performed to determine the differences among grouped data. ** Indicates statistical significance with p < 0.05.
Results
Inhibition of p53 expression promotes mesoderm differentiation of MEFs
MEFs were dissociated from mouse embryos generated by p53+/− C57BL6 and p53+/− 129 heterozygous crosses at E13.5. An allelic p53 series was generated, p53+/+, p53+/−, and p53−/− MEFs were obtained and confirmed by genotyping of the cultures. When cultured on tissue culture plates, the cell morphologies of each genotype were not significantly different at early passages (no more than three passages) (Fig. 1A–C). After extended in vitro culture (2–3 weeks), cell clusters were observed in p53−/− MEFs (Fig. 1F), whereas the morphologies of p53+/−and p53−/− MEFs did not show significant changes (Fig. 1D, E), which implied that the cellular abnormalities associated with p53−/− MEFs increase with long-term culture in vitro.

Characteristics of p53+/+, p53+/−, and p53−/− MEFs.
The expression levels of p53 mRNA and protein were verified by qRT-PCR and western blot, respectively. p53 mRNA was expressed at a very low level (Fig. 1G) in p53−/− MEFs, and protein expression was not detectable (Fig. 1H), whereas the level of both mRNA and protein expression was approximately half in the p53+/− MEFs compared with those in p53+/+ MEFs (Fig. 1G, H). Inhibition of p53 also led to increased cell proliferation rates (Fig. 1I).
Next, we detected the expression of the mesoderm marker genes, Brachyury (T) and MESP1 and the pluripotency factor Oct4 in the MEFs. T defines mesoderm during embryonic gastrulation and regulates transcription of genes required for mesoderm formation and cellular differentiation (Wilkinson et al., 1990; Wilson et al., 1995). Mesp1 is expressed in the heart precursor cells and regulates cardiac-specific gene expression (Chan et al., 2013; Saga et al., 1999). In early passages, p53−/− or p53+/− MEFs showed a higher level of T expression than that in wild-type MEFs (p53+/+) and the level of T mRNA expression negatively correlated with the level of p53 expression (Fig. 1J). The Mesp1 expression pattern is similar to T (Fig. 1K).
The results of this early mesoderm marker gene analysis showed that inhibition of p53 in MEFs promotes MEFs toward mesoderm and cardiac differentiation. Recently, it was demonstrated that Oct4 plays a role in mesoderm formation (Blin et al., 2010). To investigate the effect of p53 levels on Oct4 expression, we analyzed Oct4 mRNA expression in p53+/+, p53+/−, and p53−/− MEFs by qRT-PCR. The results show that Oct4 expression increased when p53 was inhibited in p53+/− and p53−/− MEFs (Fig. 1L).
Expression of cardiac marker genes increased in p53-decreased MEFs
The expression of cardiac marker genes is critical for cardiac transdifferentiation. To test cardiac gene expression after inhibition of p53 in MEFs, we analyzed mRNA expression of the cardiomyocyte marker genes: MYH7 (Myosin, Heavy Chain 7), MYH6 (Myosin, Heavy Chain 6), cTnT, and cardiac muscle troponin I (cTnI), in early passages (passages 2–3). The level of mRNA expression of MYH7, MYH6, and cTnI were upregulated in p53+/− and p53−/− MEFs relative to p53+/+ MEFs (Fig. 2A–C). In contrast, cTnT mRNA expression levels in p53+/− and p53−/− MEFs were lower than those in p53+/+ MEFs, and there was no significant difference between p53+/− and p53−/− MEFs (Fig. 2D). A negative correlation was found between the levels of MYH7 and MYH6 and cTnI expression and the level of p53 expression.

The expression of cardiac lineage marker genes at early passages (not more than three passages).
Inhibition of p53 enhanced MEF transdifferentiation toward cardiomyocytes and repressed endoderm and ectoderm gene expression
To systematically investigate the effect of inactivation of p53 on differentiation trends, the MEFs were grown in the specific media M199 plus Iscove's modified Dulbecco's medium (IDDM) that was used to transdifferentiate fibroblasts into cardiomyocytes (Ieda et al., 2010). To induce the MEF cultures to differentiate toward cardiomyocyte lineages, the inductive medium (M199+IDDM) was used to culture the MEFs; 21 days later the cells were dispersed into single cells with trypsin-EDTA and were analyzed by fluorescence-activated cell sorting (FACS) fr cTnT expression. The results showed that the percentage of cTnT-positive cells in the various cultures was 3.98%, 9.71%, and 7.83% in the differentiated p53+/+, p53+/−, and p53−/− MEFs, respectively, indicating that decreased p53 expression led to increased numbers of cTnT-positive cells during induction (Fig. 3A–G).

Expression of marker genes of the three germ layers after 21 days of induction.
Inhibition of p53 promoted MEF transdifferentiation into cardiomyocytes. However, we did not know if p53 levels effected endoderm and ectoderm differentiation and their respective marker gene expression. To address this gap the endoderm markers, Alpha fetoprotein (AFP) and Sox17, and the ectoderm markers, Nestin and Glial fibrillary acidic protein (Gfap), were analyzed after 2–3 weeks of MEF transdifferentiation. The results showed that all four markers were downregulated in p53+/− and p53−/− MEFs, which implied that inhibition of p53 not only promoted cardiac transdifferentiation, but also simultaneously repressed endoderm and ectoderm differentiation in this inducible system.
Overexpression of Oct4 enhanced the expression of cardiac transcription factors in MEFs
During ES cell differentiation, an Oct4-positive population of cells demonstrated a potential to differentiate toward cardiac lineages (Blin et al., 2010). To test whether Oct4 promoted cardiac differentiation of fibroblasts, pMXs-Oct4 retroviral vector was packaged and introduced into p53+/+, p53+/−, and p53−/− MEFs. Three weeks later, the expression of cardiac transcription factors was tested. As observed before, T expression was upregulated in p53+/− and p53−/− MEFs compared with p53+/+ MEFs; however, overexpression of Oct4 did not significantly change the level of T expression in p53+/+, p53+/−, and p53−/− MEFs. This analysis suggested that Oct4 was unable to affect T expression, and thus mesoderm formation, in these three cell lines (Fig. 4A).
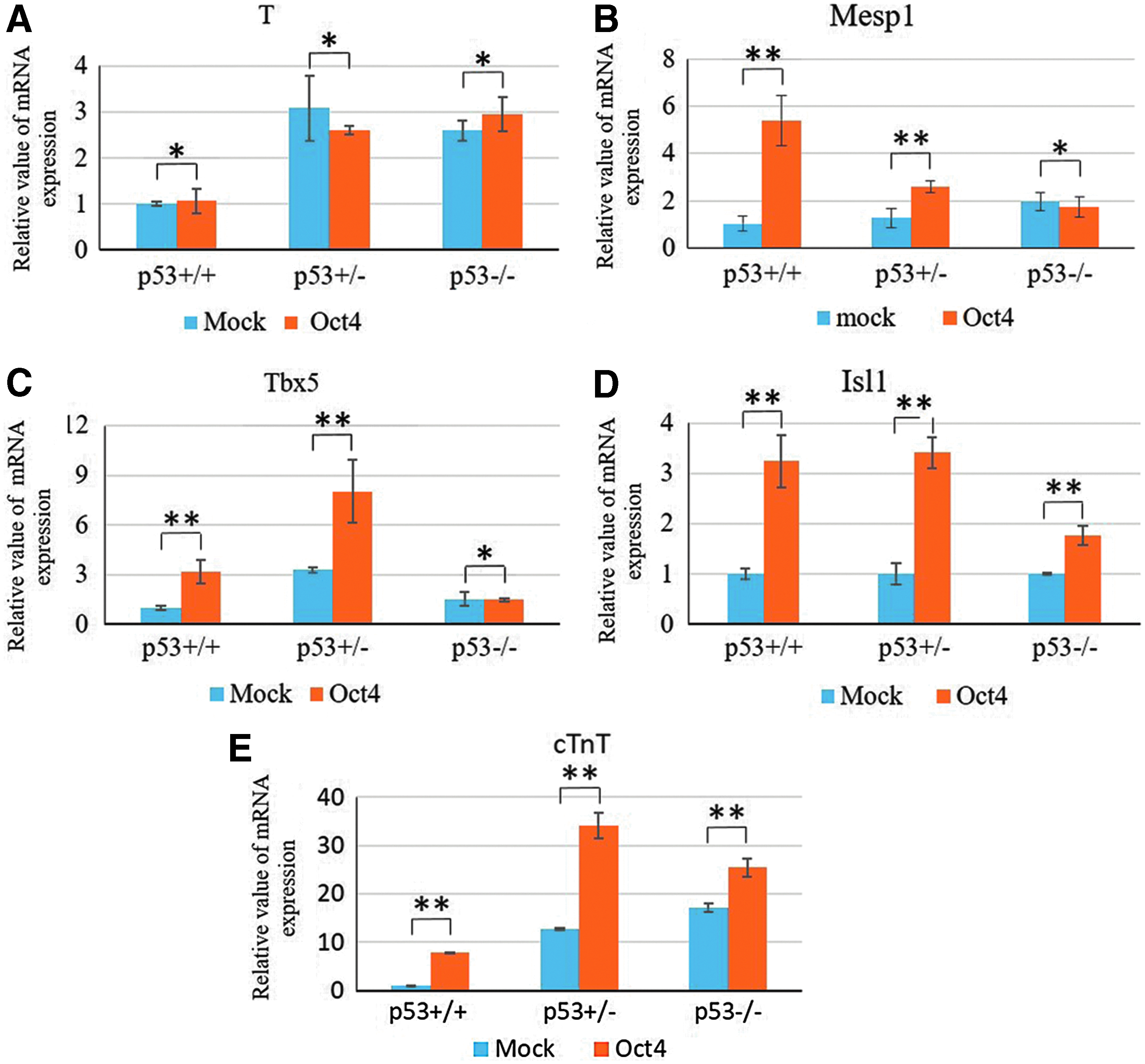
The effect of Oct4 on the expression of cardiac transcription factors verified by qRT-PCR.
The levels of Mesp1 expression were upregulated in p53+/− and p53−/− MEFs compared with wild-type (p53+/+) MEFs. Oct4 significantly enhanced Mesp1 expression in p53+/+ (p > 0.05) and p53+/− (p > 0.05) MEFs, whereas the level of Mesp1 expression was not significantly affected by Oct4 expression in p53−/− MEFs (p < 0.05) (Fig. 4B) The Tbx5 expression pattern is similar to Mesp1; however, Oct4 increased the expression of Tbx5 to its highest levels in p53+/− MEFs (Fig. 4C). Oct4 enhanced Isl1 as well as cTnT expression levels in all three cell lines (Fig. 4D, E). These results suggested that p53 expression is necessary for Oct4 promotion of the expression of cardiac transcription factors, Mesp1, Tbx5, and Isl1 and cardiac-specific marker cTnT.
Functional reciprocity between p53 and Oct4 in promoting cardiac transdifferentiation
To further investigate whether Oct4 promoted cardiomyocyte differentiation in MEFs with decreased p53, we infected the allelic series of MEFs with pMX-Oct4 retrovirus. Oct4 expression was not significantly different among p53+/+, p53+/−, and p53−/− MEFs, whereas decreased levels of p53 promoted endogenous Oct4 expression in p53+/− and p53−/− MEFs as observed before (Fig. 5A). However, the level of p53 expression was not significantly affected by overexpression of Oct4 among these cell lines (Fig. 5B). To further validate the potential of Oct4 to promote cardiomyocyte (CM) differentiation, FACS was used to analyze cTnT-positive cells after Oct4 was introduced into the MEFs.

Functional reciprocity between p53 and Oct4 in promoting cardiac transdifferentiation.
The results showed that Oct4 increased the number of cTnT-positive cells in all three cell lines (Fig. 5C, D). Western blot displayed the highest levels of protein in p53+/− cell line, which was in accordance with the levels of mRNA and the results of FACS. Although we demonstrated that inhibition of p53 upregulated cTnT expression at the level of mRNA and protein expression, overexpression of Oct4 significantly upregulated cTnT expression, and the level of cTnT mRNA and protein expression was at its highest levels when Oct4 is overexpressed in p53+/− MEFs (Fig. 5C–5E).
Discussion
This study is the first to show the reciprocity between Oct4 and p53 in transdifferentiation of MEFs toward cardiomyocytes. Decreased expression of p53 and overexpression of Oct4 both promote transdifferentiation. Oct4 plays a pivotal role in maintaining stem cell pluripotency and its levels affect cell fate decisions during early ES cell differentiation. Direct transdifferentiation of fibroblasts into cardiomyocytes has been achieved using defined lineage factors or Oct4 combined with small molecules (Wang et al., 2014). The gatekeeper of the genome, p53, is also a key factor regulating both somatic cell reprogramming and pluripotency during pluripotent stem cell differentiation (Akdemir et al., 2014; Lin et al., 2005).
In this study, we demonstrate that inhibition of p53 or overexpression of Oct4 enhanced the expression of cardiac-specific genes during transdifferentiation of MEFs toward the cardiomyocyte lineage. Furthermore, reciprocity between Oct4 and of p53 expression was found in this process. The findings raise the possibility of reprogramming endogenous fibroblasts that normally exist in the heart into functional cardiomyocytes for regenerative purposes by targeting these factors.
p53 has been verified by several groups to be a barrier to somatic cell reprogramming. Suppression of p53 increases the efficiency of direct reprogramming of fibroblasts into iPS cells (Alexandrova and Moll, 2013; Hong et al., 2009; Takenaka et al., 2010; Tidball et al., 2016; Wang et al., 2012). A recent study demonstrated that suppression of p53 enhanced the efficiency (5- to 20-fold) of direct reprogramming of human primary fibroblasts to dopaminergic neuron-like cells (Liu et al., 2014). In our study, repression of p53 promoted upregulation of lineage-restricted transcription factors, including Brachyury and Mesp1 that determine mesoderm development and cardiac lineage commitment. The cardiac-specific genes, Mhy7, mhy6, and cTnT, were also upregulated in this process.
It is obvious that inhibition of p53 promotes MEF to cardiomyocyte transdifferentiation. However, it is well known that p53 is a tumor repressor, so deletion of p53 may result in critical safety issues because it was found to cause genomic instability when mutated or deleted (Chen et al., 2012). Transient suppression of p53 with nonintegrative plasmids was also shown to improve the reprogramming efficiency of human fibroblasts (Okita et al., 2011, 2013) without affecting apoptosis and DNA damage repair (Rasmussen et al., 2014). We also found that p53 null MEFs displayed morphological instability, but hemizygous p53 expression levels in p53+/− MEFs showed morphological stability and a higher cTnT-positive population after induction.
Evidence showed that Oct4- and SSEA-1-positive cells derived from BMP2-induced differentiation of primate pluripotent stem cells give rise to endo/mesodermal cells secreting cardiogenic factors, further directing the cell fate toward cardiac lineages, including cardiomyocytes as well as smooth muscle and endothelial cells (Blin et al., 2010). Precise levels of Oct4 expression have been shown to decide the fate of differentiating ES cells. A <2-fold increase from normal expression levels causes differentiation into ectoderm and mesoderm, whereas inhibition of Oct4 results in rapid differentiation of ES cells and a significant increase in transcription of genes associated with trophoblastic and endoderm lineages (Hay et al., 2004; Niwa et al., 2000; Wang et al., 2013).
Overexpression of Oct4 promoted human ES cells and iPS cells to differentiate into cardiac progenitors by activation of Sox17 (Stefanovic et al., 2009). Overexpression of Oct4 in MEFs upregulated the mesoderm gene, T and cardiac transcription factors, Tbx5, Nkx2.5, and Mef2c, which would promote the expression of cardiac-specific genes, such as cTnT, and thus enhanced MEF differentiation toward cardiac lineages (Addis et al., 2013).
Although overexpression of Oct4 did not downregulate p53 expression, increased expression of Oct4 in p53 null or heterozygous MEFs promoted transdifferentiation toward cardiac lineages compared with increased Oct4 expression alone or downregulation of p53 alone. This in turn lead to higher levels of Mesp1, Isl1, Tbx5, and cTnT expression in hemizygous p53 (p53+/−) MEFs compared with p53 null (p53−/−) MEFs. Oct4 alone enhances the expression of important transcription factors: Mesp1, and Tbx5 in p53+/+ MEFs and p53+/− MEFs other than p53 null (p53−/−) MEFs, which demonstrates that p53 is probably necessary for the process of cardiomyocyte reprogramming and that complete deletion of p53 would limit cardiomyocyte transdifferentiation.
In addition, p53-deleted fibroblasts displayed overgrowth and tumorigenesis, leading to an inability to completely differentiate into the normal cell type, which was probably another reason for the relatively low efficiency of transdifferentiation. To avoid genomic instability caused by complete deletion of p53 and sequential tumorigenesis, transient inhibition of p53 or partial inhibition of p53 would be an optimal strategy for directly reprogramming without affecting genome stability (Rasmussen et al., 2014).
Directly reprogramming somatic cells into cardiomyocytes has been achieved, however, its efficiency and safety should be considered. Increasing transcription factor expression could improve the efficiency of reprogramming, but the more factors used in the process will lead to more potential for genomic instability, which will bring more risks to patients. In our experiments, decreased p53 (p53+/− MEFs) alone promotes about 10% fibroblast transdifferentiation into a cTnT-positive population, and cTnT-positive population elevated to about 15% after p53+/− MEFs were treated with Oct4.
Our findings suggest that a combination of increasing Oct4 and inhibiting p53 are reciprocal for promoting cardiomyocyte transdifferentiation. Furthermore, the advantage of p53 inhibition may reduce the required amount of exogenous genes used in transdifferentiation, which will reduce the risk of tumorigenesis for patients, thus promoting transdifferentiation of fibroblasts into cardiomyocytes for clinical purposes.
Another issue that should be considered is that atrial or ventricular lineages owe their individual traits to a combination of electrophysiology, gap junction, lineage markers, etc. To gain the functional cardiomyocytes in damaged heart is pivotal and large hurdles to restore the cardiac function for clinical therapy remain. Furthermore, transplantation of mixed cardiac lineages possibly causes arrhythmia (Liao et al., 2010), thus development of transdifferentiation methodologies to generate chamber-specific cardiomyocytes to reach clinical purposes need to be resolved.
Footnotes
Acknowledgment
This work was supported in part by an NIH grant (R01 HL121294-01A1) to T.R.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.