
Abstract
Abstract
In this study, we found that the measles virus (MV) can infect human-induced pluripotent stem cells (hiPSCs). Wild-type MV strains generally use human signaling lymphocyte activation molecule (SLAM; CD150) as a cellular receptor, while vaccine strains such as the Edmonston strain can use both CD150 and CD46 as receptors. It is not yet known how early in the embryonal differentiation stages these receptors are expressed. We established two hiPSCs (BGU-iPSCs and EMF-iPSCs) which express CD46 and CD150. Both cell types can be infected by MV to form persistent, noncytopathic cell lines that release infectious MV particles. Following MV persistent infection, BGU-iPSCs and EMF-iPSCs remain pluripotent and can differentiate in vitro into the three germ layers. This includes cells expressing the neuronal differentiation markers: NF68 and miRNA-124. Since the MV does not integrate into the cell's genome, it can be utilized as a vehicle to systematically introduce genes into iPSC, to dissect and to define factors regulating lineage differentiation.
Introduction
M
MV initially infects a variety of lymphoid and nonlymphoid peripheral organs. More rarely, the MV can persistently infect cells within the central nervous system (CNS). MV proliferates in epithelial cells, then infects secondary target cells, such as dendritic cells, lymphocytes, and monocytes, and ultimately spreads throughout the body via the blood and lymphatic fluids (Dörig et al., 1993; Yokota et al., 2008). Vaccine strains of MV such as the Edmonston strain use human CD46 as a cellular receptor. CD46 is an ubiquitously expressed complement regulatory protein that protects host cells from injury by preventing accidental complement deposition on host cells (Liszewski and Atkinson, 2015). As CD46 is expressed on all nucleated human cells, vaccine strains of MV can infect almost any human cell type (Evlashev et al., 2000; Horvat et al., 1996; Manchester and Rall, 2001).
In contrast, wild-type strains of MV instead use CD150 (also known as signaling lymphocyte activation molecule [SLAM]) as receptor for infection (de Swart et al., 2007). CD150, generally considered as a cellular receptor for both vaccine and wild-type strains of MV (de Swart, 2008), is a costimulatory molecule in lymphocyte activation, and its expression is restricted to activated T and B lymphocytes, immature thymocytes, mature dendritic cells, and activated monocytes (Hashimoto et al., 2002; Ohno et al., 2007; Yanagi et al., 2006). CD147/EMMPRIN (extracellular matrix metalloproteinase inducer) has been identified as a third MV receptor on epithelial cells (Watanabe et al., 2010).
MV has a well-established propensity to invade the CNS and develop an apparently noncytopathic persistent infection within neurons (Chakraborty et al., 2010; Gopas et al., 1992; Segev et al., 1995). Some studies in vitro have demonstrated MV persistent infection of non-neural cell lines such as HeLa cells, Hep2 cells, and monkey kidney cells (Doi et al., 2016; Rima and Duprex, 2005). The factors that allow the MV to persist remain largely unknown; studies suggest that miRNAs are among the host molecules that viruses co-opt to suppress their own replication to evade immune elimination and establish a persistent infection (Mahajan et al., 2009).
miRNAs are a class of ∼22 nucleotide long noncoding RNAs that are transcribed from the genomes of all multicellular organisms and some DNA viruses (Bartel, 2004; Cullen, 2004). Specific miRNAs have been implicated in diverse biological processes, including development, cellular differentiation, proliferation, apoptosis, and oncogenesis (Bushati and Cohen, 2007). Expression of miR-124 increases over time in the developing nervous system (Smirnova et al., 2005), and neuronal differentiation is enhanced following ectopic expression of miR-124 in mouse neuroblastoma cells (Makeyev et al., 2007), mouse embryonal carcinoma cells, and mouse embryonic stem cells (ESCs) (Krichevsky et al., 2006), as well as neuronal differentiation of postnatal neural stem cells (Silber et al., 2008).
MV infection may spread to the CNS causing several types of devastating neurological diseases in a small percentage of measles cases, such as fatal subacute sclerosing panencephalitis (Buchanan and Bonthius, 2012). The molecular mechanisms of CNS infection and the specific lineage target of MV tropism are not well understood, but based on studies in transgenic rodent models, a complex interplay of innate and adaptive immune system responses is involved (Schneider-Schaulies et al., 2003).
In this work we investigated whether pluripotent stem cells can be persistently infected with MV. Human stem cells or human-induced pluripotent stem cells (hiPSCs) are considered to be a powerful in vitro system for studying differentiation and generation of human tissues (Thomson et al., 1998) and for unveiling the mechanism of development of monogenic and complex human diseases (Nakamura et al., 2013; Richard and Maragakis, 2015; Sanchez-Danes et al., 2012), including viral diseases (Berger et al., 2015). Generating hiPSCs involves reprogramming of fibroblasts by infecting these cells with lentivirus containing human Oct4, Sox2, Klf4, and c-MYC genes. A novel reprogramming process based on vectors derived from the nonintegrating vaccine strain of MV was established (Driscoll et al., 2015).
Differentiating hiPSCs in vitro generate embryoid bodies (EBs) that develop into embryonic three germ layers (endoderm, mesoderm, and ectoderm) (Thomson et al., 1998). There are protocols for induction of differentiation into terminal cell types like cardiomyocytes (Nakamura et al., 2013), neuronal progenitors (Chambers et al., 2009; Cohen et al., 2007; Liu et al., 2012), GABAergic neurons (Liu et al., 2013), and dopaminergic neurons (Sanchez-Danes et al., 2012).
Human ESCs (hESCs) and iPSCs provide an opportunity to study human neural development (Petros et al., 2011; Reubinoff et al., 2000; Thomson et al., 1998) and may be useful for unveiling the neuronal lineage relevant to MV tropism persistence and disease.
In this study, we describe that iPSCs express the MV receptors CD46 and CD150; they can be persistently infected by MV and produce infective MV particles. Infective particles are produced both in nondifferentiated and differentiated iPSCs. The infected cells remain pluripotent and can differentiate in vitro into the three germ layers.
Materials and Methods
Reprogramming
For generating the line BGU-hiPSCs, a layer of peritoneum that surrounds the abdominal organs was obtained from a patient that underwent gallbladder surgery in the Soroka University Medical Center. The hospital's committee of ethics approved this study, and informed consent was obtained from the patient (Health office No.: 920100231).
The tissue was minced with scissors and scalpels into small (<3 mm) pieces, followed by digestion with 0.5 mg/mL collagenase type I (Worthington Biochemical, Lakewood, NJ). Digested tissue was centrifuged at 1000 rpm. Supernatant containing adipocytes were removed, and the cell pellet was resuspended in DMEM (Biological Industries, [BI], Israel) supplemented with 10% fetal bovine serum (FBS; BI). Resuspended cells were filtered through 100 μm cell strainer (Becton Dickinson, Franklin Lakes, NJ) followed by passage through a 40 μm cell strainer. Cells were resuspended and plated in DMEM supplemented with 10% FBS.
To generate the line EMF-iPSCs, human foreskin fibroblasts (EMF; kindly provided by Prof. Joseph Itskovitz-Eldor; Technion, Haifa, Israel) were used. For reprogramming, 40,000 cells per well in a 24-well plate were transduced with lentivirus containing human Oct4, Sox2, Klf4, and c-MYC produced in 293T cells as described before (Takahashi et al., 2007). After viral transduction, cells were allowed to recover between 24 and 48 hours before being transferred onto plates with mitomycin treated mouse embryonic fibroblast (MEF) as feeder cells (36,000 cells/cm2). Transduced cells were cultured in the defined stem cell media NutriStem (BI). hiPSC colonies (termed in this study as BGU-iPSCs and EMF-iPSCs) were picked after 3 and 5 weeks of culture and subcultured clonally on MEFs in six-well tissue culture plates.
ESC-like colonies with distinct light refractive property were picked out and seeded on Matrigel surface (BD Matrigel HESC Qualified; BD Biosciences, San Diego, CA; www.bdbiosciences.com) in a new culture dish. Each single picked colony was maintained and expanded following routine ESC passaging and culturing protocols and were established as a hiPSC line in NutriStem medium.
Cell culture
The established BGU-iPSC and EMF-iPSC lines were grown on Matrigel surface in NutriStem medium and were characterized by flow cytometry, in vitro, EB formation, and in vivo teratoma assays. The cells had a normal diploid karyotype (data not shown).
EB assay
hiPSCs were grown in ultralow attachment plates in suspension culture for 4–8 days with DMEM/F12 (1:1) containing 20% knockout serum (Invitrogen), 4.5 g/L
Teratoma assay
All mouse procedures were conducted under local ethical guidelines and after gaining permission from the local Animal Ethics Committee of the Ben-Gurion University of the Negev (IL04-06-2014). The mice were NOD SCID (Harlan laboratories). BGU-iPSCs were grown to near confluence, harvested using collagenase IV treatment, washed in NutriStem, and resuspended in NutriStem supplemented with 30% Matrigel (BD Biosciences). Methoxyflurane-anesthetized mice were intramuscularly injected in the hind leg with 50 μL of ∼1 × 106 cells at one site only. Before teratoma removal, mice were sacrificed by Methoxyflurane. Teratomas were surgically removed, fixed in 4% formalin, and embedded in paraffin. Sections were mounted onto slides, stained with hematoxylin/eosin, and examined for the presence of representatives of the three germ layers.
Persistent infection
BGU-iPSCs and EMF-iPSCs (1 × 106/well) were seeded on a six-well Matrigel covered plate in 2 mL stem cell media (NutriStem) and grown overnight. Infection with 2 × 106/mL PFU of GFP labeled MV wild-type strain, IC323-EGFP (GFP-MV; kindly provided by Dr. Yusuke Yanagi) (Hashimoto et al., 2002), in RPMI 1640, and 2% FCS for 60 minutes at 37°C, was performed and the plates were shaken every 15 minutes and washed thrice with stem cell media. Two milliliters of stem cell media was finally added to each well. Dead cells were removed, and the surviving cells were carried further in stem cell media for 30 days. The remaining attached cells were GFP-MV positive indicating that they were persistently infected.
To show that infected hiPSCs produce infectious virus, BGU-iPSCs and EMF-iPSCs were persistently infected with GFP-MV (2 × 106/mL PFU) as mentioned above. Thirty days postinfection the cells spontaneously differentiated in vitro and formed embryonic bodies (EBs) for 4 days and were then plated on gelatin for additional 21 days. Supernatants at 4 days of differentiation (before plating on gelatin) and 21 days of differentiation on gelatin were collected and coincubated with VERO-SLAM cells, expressing CD150 kindly provided by Dr. Yusuke Yanagi (Hashimoto et al., 2002), 0.5 × 105 cells per well in 24-well plates in triplicate. GFP fluorescence of VERO-SLAM viable cells was followed for 2 weeks, and pictures were taken by confocal microscopy.
Quantitative PCR (qPCR) analysis of miRNAs in BGU-iPSCs and MV-BGU-iPSCs
cDNA libraries were prepared using miScript Reverse Transcription Kit (Qiagen, Hilden, Germany). Briefly, 1 μg of total RNA was poly-adenylated by poly (A) polymerase and converted into cDNA by reverse transcriptase with oligo-dT in a single step, at 37°C for 1 hour and at 95°C for 5 minutes. The cDNA libraries were used in the qPCR analysis for miRNAs using miScript SYBR Green PCR Kit (Qiagen) with a forward specific miRNA primer (sequences were taken from the databases (DS) analysis; Metabion Martinsried, Germany) and the miScript Universal reverse primer. Forward primers with high G/C content at the 3′ end were extended with one or two “A” nt at their 3′-end. qPCRs were performed using LightCycler 480 system (Roche Applied Science, Manheim, Germany). miRNA hsa-124a-3p was used as a reference gene (sequence of the forward primer: 5′-UAAGGCACGCGGUGAAUGCC-3′).
Flow cytometry
Immunostaining was performed according to the following protocol: BGU-iPSCs were washed with phosphate-buffered saline (PBS) containing 0.1% FBS (FACS buffer), fixed in paraformaldehyde (4% in FACS buffer) for 10 minutes at room temperature (RT), washed in FACS buffer, and incubated with primary antibody against stage-specific mouse anti-human embryonic antigen-4 (SSEA4); (1:150; Santa Cruz Biotechnology, Inc.). The cells were then washed and incubated with the following secondary antibodies: Alexa 488 goat anti-mouse (KPL 202-03-18-06 DyLight 488 Labeled; Epitomic, Inc.). Cells were analyzed by flow cytometry (BD FACSCanto™ II; BD Biosciences). FACS for MV receptor CD46: mouse anti-human CD46 (BioLegend); Anti-human SLAM/CD150 (R&D Systems). Second antibody: goat anti-mouse polyclonal IgG DyLight 650 (red like Cy5) (Epitomic, Inc).
Immunocytochemistry
BGU-iPSCs grown on Matrigel (BD Biosciences) or gelatin-coated 24-well tissue culture plates were washed with PBS and fixed with cold ethanol at −20°C for 15 minutes. After the cells were air-dried, they were blocked with hESC medium (Knockout DMEM; Rhenium, 20% Knockout Serum Replacement; Rhenium, 4 ng/mL bFGF; Rhenium, 1% Pen-Strep-Ampho Solution; BI, 1%
The cells were washed and incubated with secondary antibody in hESC medium for 1 hour at RT. Second antibody for Oct3/4 and SSEA4 was Alexa 488 goat anti-mouse KPL 202-03-18-06 DyLight 488 Labeled, and second antibody for NF68, AFP, and SMA was goat α Rabbit-FITC Santa Cruz Biotechnology, Inc. The cells were washed and visualized under a fluorescent microscope (Olympus IX70).
Gene expression analysis by qPCR
Total RNA was isolated by the RNeasy Mini Kit (Qiagen Cat. No. /ID 74104). The concentration and purity were measured on a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific). RNA from each sample was reverse transcribed into cDNA using Verso cDNA Reverse Transcription Kit (AB-1453; Thermo Scientific). Gene expression analysis was performed using TaqMan gene expression assays (Applied Biosystems). Reactions were run on ABI-7500 and reaction conditions included 40 cycles of 95°C for 10 seconds, followed by 60°C for 30 seconds. Relative quantity (RQ) of gene expression of target genes was calculated as RQ of expression in MV infected cells compared to uninfected cells, using GAPDH as the normalization gene (Pfaffl, 2001; Pfaffl et al., 2002). Bars represent ± STDEV (standard deviation) or SE (standard error).
Results
Characterization of BGU-iPSCs
We generated a hiPSC line from human cells derived from the layer of peritoneum that surrounds the abdominal organs (termed in this study as BGU-iPSCs) that fulfills all the criteria for pluripotency (Senju et al., 2009). These cells were reprogrammed using lentivirus containing a polycistronic reprogramming cassette composed of a CAG promoter driving expression of Oct4, Sox2, Klf4, and c-Myc linked by 2A self-cleaving peptides (Sommer et al., 2009). The BGU-iPSC colonies exhibited typical ESC morphology and expressed the pluripotent cell markers Oct4, SSEA4, and TRA-1-60 (Fig. 1A). The pluripotency of the cells was further demonstrated by in vitro and in vivo differentiation into the three germ layers in EBs and teratomas (Fig. 1B, C, respectively).
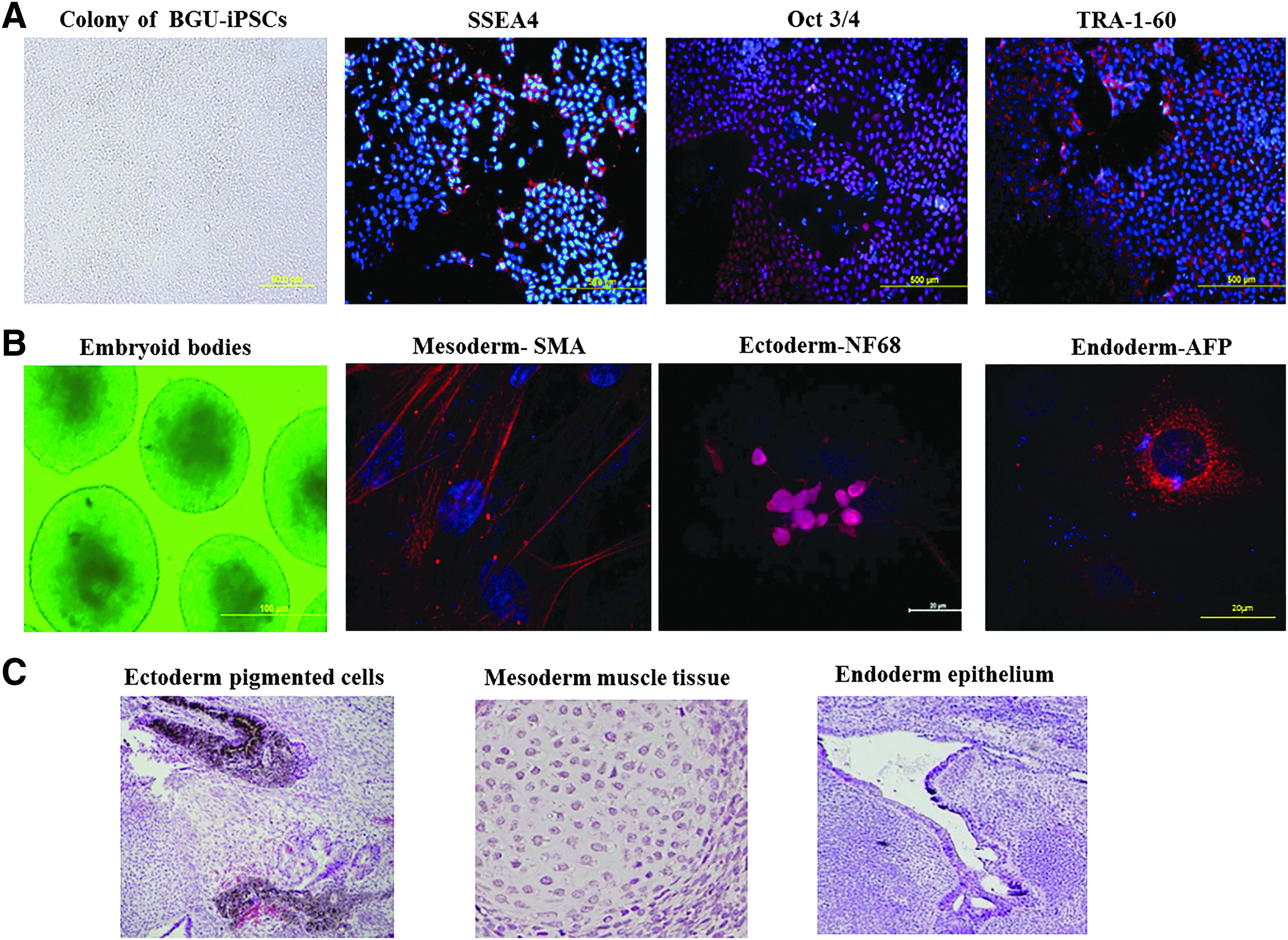
Characterization of BGU-iPSCs.
We investigated whether BGU-iPSCs express MV receptors and are permissive for infection with MV. We found that 25% of BGU-iPSCs were strongly CD150 positive (Fig. 2A). Although the persistent infection doubled the percentage of CD150 positive cells (52%), the cells expressed lower amounts of CD150 per cell. All cells, including CD150 negative cells, remained MV (GFP) positive 21 days postinfection, suggesting that the virus can enter these cells independent of the CD150 receptor (Young and Rall, 2009).

BGU-iPSCs express the MV receptors. BGU-iPSCs were infected with GFP-MV (2 × 106/mL PFU).
Wild-type virus is required for CD150 infection; therefore, it is likely that only the CD150 positive cells will be infected. In addition, FACS analysis showed that 98.4% of the BGU-iPSCs expressed the pluripotent marker SSEA4 and 98.6% the MV receptor CD46 (Fig. 2B).
iPSCs remain pluripotent after MV persistent infection
We infected two human iPSC lines created in our laboratory: BGU-iPSCs and EMF-iPSCs with GFP-MV. FACS analysis indicated that the pluripotent marker SSEA4 was expressed on the cell surface before and after infection (Fig. 3A, B, respectively). The expression of the pluripotent markers NANOG, POU5, and SOX2 was demonstrated by qPCR analysis (Fig. 3C). Although the pluripotent markers tested were upregulated in MV infected BGU-iPSCs but not in infected EMF-iPSCs, both cell lines remained pluripotent following infection. Using fluorescence microscopy, we could determine the presence of GFP positive cells overtime. As seen in Figure 3D, at 20 days postinfection, most of the BGU-iPSCs were infected and expressed as GFP-MV. EMF-iPSC infected cells showed a similar pattern of infection (not shown).
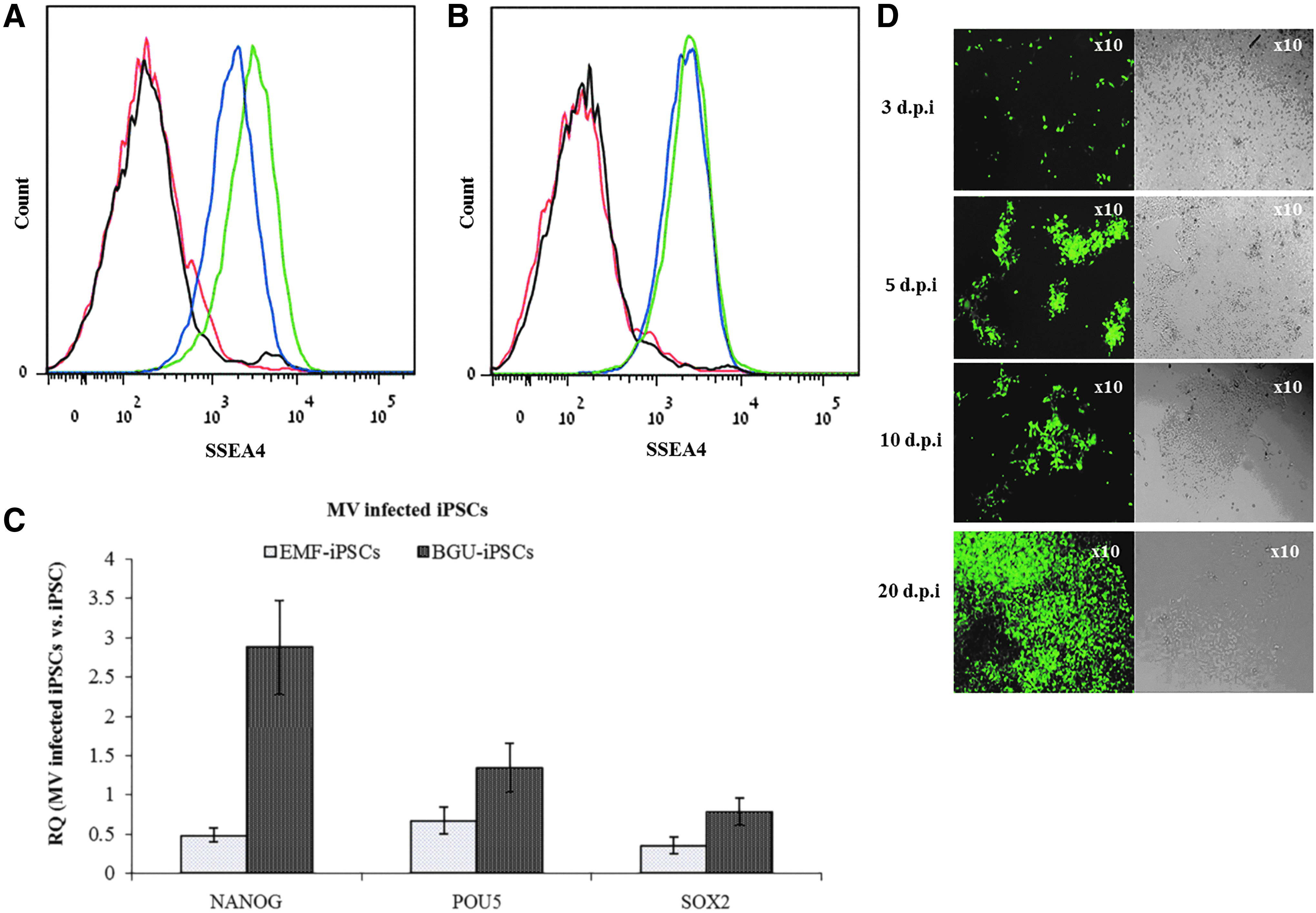
Expression of pluripotent markers in noninfected and MV infected cells. FACS analysis of the pluripotent marker SSEA4 on
Infection with GFP-MV did not interfere with spontaneous differentiation in vitro
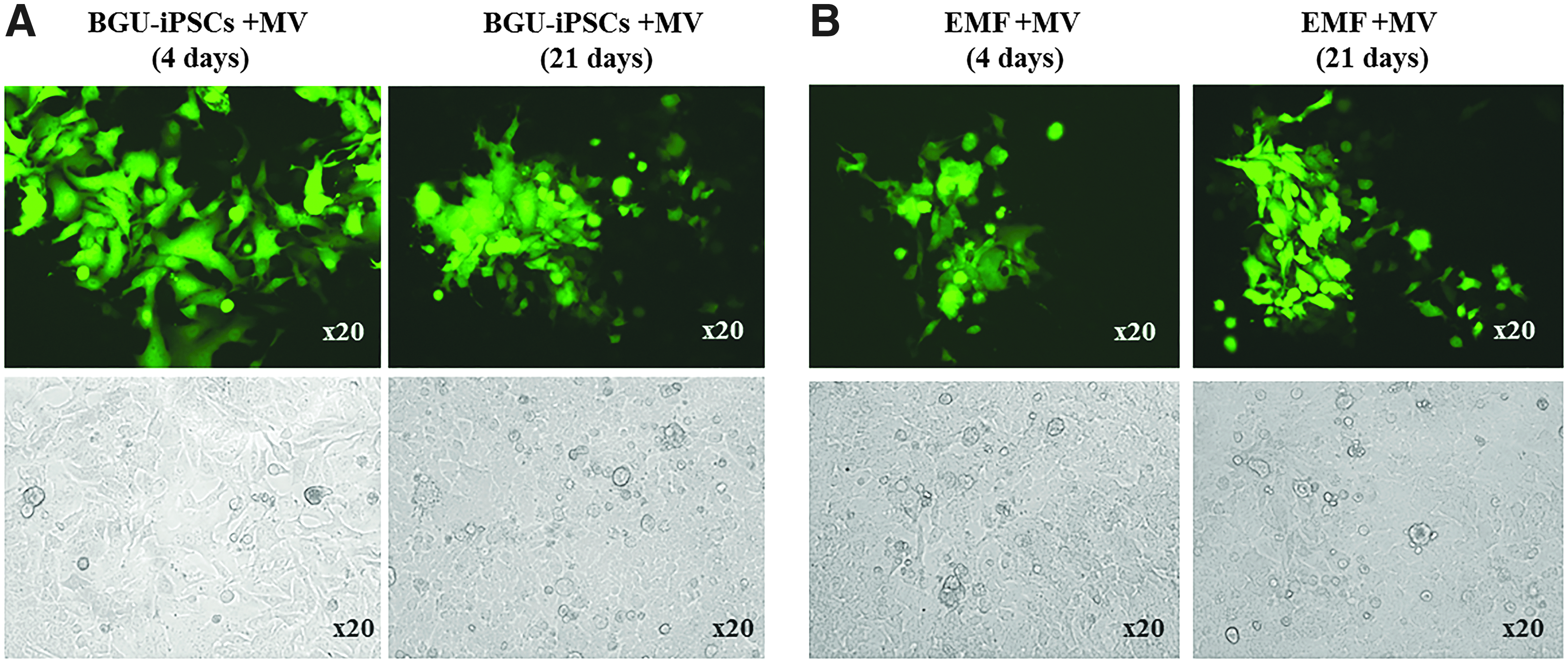
It was an open question whether following infection with MV, iPSCs will be able to form EBs and differentiate in vitro into embryonic germ layers. As seen in Figure 4, MV infected BGU-iPSCs and EMF-iPSCs, similarly to noninfected cells, spontaneously differentiated to form EBs; spontaneously differentiated to form mesoderm (stained for SMA) and ectoderm (stained for NF68); endoderm markers were not tested. The differentiated cells remained positive for the GFP-MV.

Infection with GFP-MV did not interfere with spontaneous differentiation. Fluorescent GFP-MV persistently infected MV-BGU-iPSCs and MV-EMF-iPSCs spontaneously differentiated to form EBs for 4 days. Bright field, GFP-MV fluorescence microscopy (magnification 10 × ), 21 days after plating of EBs on gelatin, the cells spontaneously differentiated into mesoderm and were stained for SMA (red), and ectoderm, shown by staining for NF68 (also a neuronal marker) (red). Nuclear DAPI stain (blue). Following differentiation, all cells remained GFP-MV positive. Pictures were taken by confocal microscopy. EBs, embryoid bodies.
Infection of VERO-SLAM cells with GFP-MV derived from cell-free supernatants of persistently infected iPSCs
We showed that MV particles derived from differentiated persistent MV infected BGU-iPSCs or MV infected EMF-iPSCs were infective. VERO-SLAM cells were incubated with supernatant collected at day 0 and at day 21 of differentiation. GFP (GFP-MV fluorescence) of viable cells was followed for 2 weeks, and pictures were taken by confocal microscopy (Fig. 5A, B). These results confirm that MV derived from the persistently infected cells is capable of infecting naive cells.
MV derived from cell-free supernatants of differentiated MV-BGU-iPSCs or MV-EMF-iPSCs is infective. VERO-SLAM monolayers were infected with MV-GFP derived from cell-free supernatants of MV-BGU-iPSCs
Persistent MV-BGU-iPSC expresses high amounts of miR-124
Since MV persistently infect neuronal cells, and miR-124 is a differentiation marker of neuronal cells, we assessed whether miR-124 is expressed in noninfected and/or in MV persistently infected iPSCs. We showed that BGU-iPSCs express miR-124. In comparison, the levels of miR-124 were increased threefold in persistently infected cells (Fig. 6).
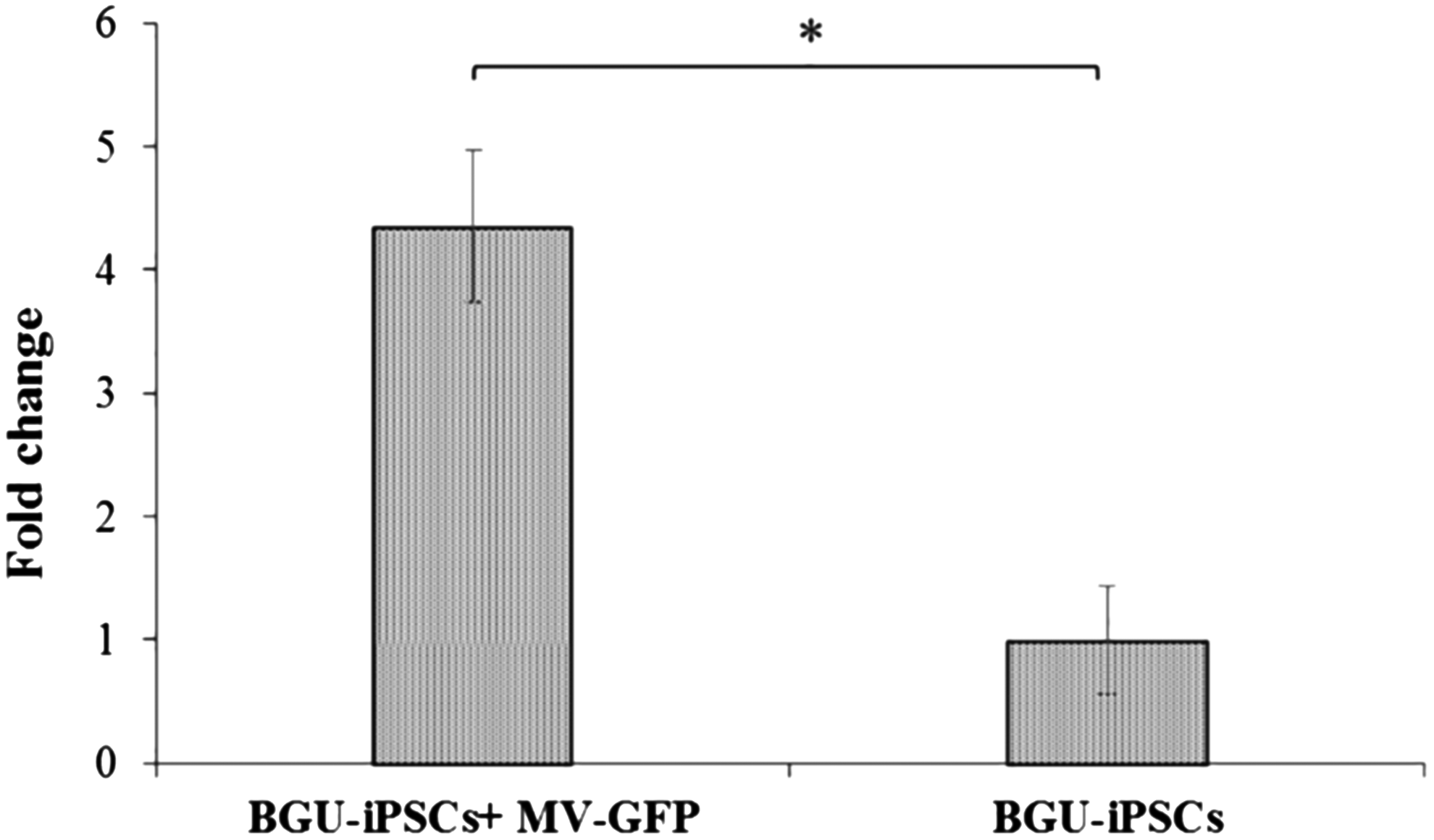
Expression of miR-124 in BGU-iPSCs increased following persistent infection with GFP-MV. Twenty d.p.i., cDNA libraries were prepared, and miRNA-124 relative expression was determined by qPCR and expressed as the fold change relative to U6. Expression in the uninfected BGU-iPSC line was used as the reference. Three independent cDNA libraries were tested thrice; averages with SEs are indicated. Average Cq ± STDEV is presented. T-tests were performed, and p < 0.05 significance between samples is shown (*). SE, standard error; STDEV, standard deviation.
Discussion
In this study we showed that human iPSCs express the MV receptors CD46 and CD150 and can be persistently infected by MV.
The MV infected iPSCs keep their pluripotent stem cells characteristics as follows: they express pluripotent cell markers when seeded on Matrigel, they form EBs in vitro when seeded on ultralow attachment plates, and differentiate into embryonic germ layers when seeded on gelatin. In addition, viral particles derived from these cells were able to infect naive VERO-SLAM cells. Both cell lines remained pluripotent after MV persistent infection although the pluripotent markers tested were upregulated in MV infected BGU-iPSCs but did not vary in MV infected EMF-iPSCs. These results probably reflect the difference in the iPSCs' cell of origin.
The reported that functions of CD46 are related to its role as a cofactor for complement factor I and as a costimulatory factor for T cells which induces the differentiation of CD4+ into T-regulatory 1 cells (Astier et al., 2000; Kemper et al., 2003). To our knowledge, this is the first time that the expression of CD46 on pluripotent stem cells has been documented. CD46 is now considered as a “multitasker molecule” and is recognized for its roles in linking innate and adaptive immune responses (Liszewski and Atkinson, 2015). Furthermore, we found that pluripotent stem cells BGU-iPSCs express CD150, even before differentiation.
Only a small percentage of uninfected cells were strongly positive (25%). Interestingly, on one hand the percentage of CD150 positive, persistently infected cells doubled, compared to noninfected cells but on the other, each cell expressed lower amounts of CD150 (Fig. 2). Since the number of CD150 positive cells increases in infected cells but the cells CD150 relative amounts decreases, it is possible that this effect is a result of differentiation of MV-BGU-iPSC toward mesoderm (Fig. 4). It has been reported that the expression of CD150 on peripheral blood monocytes could be induced by mitogen and cytokine stimulation and also by MV particles (Browning et al., 2004; Kruse et al., 2001; Ono et al., 2001; Veillette et al., 2007; Yurchenko et al., 2010). It was also suggested that after infection of cells, and after surface contact with MV envelope proteins.
CD150 is downregulated from the cell surface of activated PBL and cell lines (Erlenhoefer et al., 2001). Welstead et al. demonstrated that expression of the hemagglutinin (H) protein of MV was sufficient for CD150 downregulation but not of CD46 (Schneider-Schaulies et al., 2001; Welstead et al., 2004). In these articles it is proposed that the H protein interacts either with SLAM on the cell surface or within the endoplasmic reticulum (ER) downregulating SLAM expression. It is possible that these mechanisms take place also in our system.
MV infected BGU-iPSCs and EMF-iPSCs are able to differentiate in vitro into EBs and release infective MV particles. This suggests that MV, a virus, does not integrate into the host-cell genome and, in principle, does not interfere with the transcriptional-differentiation program of the pluripotent stem cells, as determined by the differentiation markers tested.
Interestingly, and supporting our observation, it has been recently reported that newborn somatic cells were reprogrammed into iPSCs using the reprogramming vector cocktail with the OCT4-expressing MV vector and SOX2-, KLF4-, and cMYC-expressing lentiviral vectors (Driscoll et al., 2015). Furthermore, the results presented in this study support the idea that MV can be utilized to persistently infect iPSCs with the potential to introduce genes of interest without affecting the differentiation program of the cells. Thus, MV has an advantage over the congenital human cytomegalovirus (CMV) model reported by Berger et al. (2015) in which CMV infected only neural precursors but not undifferentiated hESCs.
Most RNA viruses, including MV, do not encode microRNAs (Trobaugh and Klimstra, 2017). Individual microRNAs may regulate several 100 genes, and it is estimated that more than 30% of animal genes may be subject to microRNA control (Lewis et al., 2005). In this study we found that BGU-iPSCs express miR-124, one of the most conserved and abundantly expressed neuronal-specific miRNAs (Lagos-Quintana et al., 2002). The sequence of mature miR-124 is completely conserved from worms to humans (Lewis et al., 2005). MiR-124 is expressed in neurons, but not astrocytes, and the levels of miR-124 increases over time in the developing neural cell (Smirnova et al., 2005).
Recently we have demonstrated that in MV persistently infected human neuroblastoma cells, high expression of miR-124 correlates with reduced cell division (Naaman et al., 2017). In this work we showed that expression of miR-124 in MV persistently infected BGU-iPSCs was upregulated compared to noninfected cells implying that either more cells became miR-124 positive cells and/or that MV infection led to upregulation of miR-124 expression. In conclusion: wild-type MV can persistently infect hiPSCs that can differentiate in vitro and produce infective particles. We suggest that MV persistently infected cells may provide viral particles for long-term immunity, as well as in rare cases, MV that induce CNS diseases.
Footnotes
Acknowledgments
The authors acknowledge support from NIH R03 AI094164-01A1, the Richard H. Holzer Foundation, and the American Associates of the Ben-Gurion University (AABGU) the Philadelphia Chapter.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.