
Abstract
Abstract
Dental pulp stem cells (DPSCs), as one type of mesenchymal stem cells (MSCs), have the capability of self-renewal and multipotency to differentiate into several cell lineages, including osteogenesis, odontoblasts, chondrogenesis, neurogenesis, and adipogenesis. It has found that tumor necrosis factor-α (TNF-α) can promote osteogenic differentiation of human DPSCs in our previous studies. Other experimentation revealed that signal transducer and activator of transcription 3 (STAT3) underwent a rapid activation both in osteogenesis and inflammation microenvironment of MSCs in vitro. MicroRNAs (miRNAs or miRs) have been proved in previous studies to regulate MSCs differentiation in vitro. In this study, we identified miR-21 as a key miRNA contributed the functional axis of odontoblast differentiation induced by STAT3. It is observed that the expression of miR-21 and STAT3 increased gradually in low concentration (1–10 ng/mL) of TNF-α, while they were suppressed in high concentration (50–100 ng/mL). The upregulation of miR-21 may facilitate the odontoblast differentiation of DPSCs coordinating with STAT3. SiSTAT3 or treated by the inhibitor of STAT3, cucurbitacin I (Cuc I), significantly increased primary miR-21 expression along with decreased mature miR-21 expression. Meanwhile, the inhibition of miR-21 (anti-miR-21) decreased the activation of STAT3 as well as suppressed the marker proteins of odontoblast differentiation. The results revealed a new function of miR-21, suggesting that miR-21/STAT3 signal may act as a modulator within a complex network of factors to regulate odontoblast differentiation of human DPSCs. It may provide a novel therapeutic strategy to regulate the odontoblast differentiation of DPSCs.
Introduction
D
The environment of receptor regions is generally uncertain, including inflammation microenvironments and stress state (Yang et al., 2012). Tumor necrosis factor α (TNF-α), a major proinflammatory cytokine, can lead to bone loss in numerous inflammatory diseases (Zhang et al., 2015). Similar to periodontal diseases, TNF-α is a key proinflammatory cytokine produced upon periodontal inflammatory response, which actively participates in osteoclastogenesis (Page, 1991). Our previous studies have shown that TNF-α stimulates osteogenic differentiation of DPSCs positively in low concentration (10 ng/mL) and negatively in higher concentration (50, 100 ng/mL) (Feng et al., 2013, 2015).
The proliferation and differentiation of DPSCs are well controlled by a designed genetic program. MicroRNAs (miRNAs or miRs) are small (17–25 nucleotides) and noncoding RNAs that participate in the regulation of gene expression, involving in a wide spectrum of biological processes (Wang et al., 2013), including cell proliferation and apoptosis, energy metabolism, major signal pathways, tissue morphogenesis, and tumors (Kumarswamy et al., 2011). In mammals, the active miRNAs regulate the gene expression by binding to complementary sequences in the 3-untranslated region (3′ UTR) of target mRNAs and then leading to mRNA translational repression, degradation, or other changing (Bae et al., 2012; Thomas et al., 2010). Abnormal expression of miRs associates with various developmental disorders and diseases (Moriyama et al., 2009).
Recent research found that expression of miRNA-21 (miR-21), which can reduce Smad7 levels to maintain BMP9/Smad signal activation, was upregulated during the osteogenic differentiation of murine multilineage cells (Song et al., 2015). MiR-21 is upregulated in asthma, a chronic inflammatory disease affecting airways (Elbehidy et al., 2016). Previous researches have also shown that the miR-21 binds with several transcriptional factors, such as transducer and activator of transcription 3 (STAT3), AP1, p53, and so on, influence the expression of downstream target genes (van der Fits et al., 2011).
STAT3, a member of the STAT protein family, has been known as an important regulator in tumor cells and also plays a critical role in inflammation and tumorigenesis by regulating cell metabolism and differentiation (Kathiria et al., 2012). STAT3 protein exists in an inactive form in the cytoplasm and can be activated by associated kinases and phosphorylated at diverse phosphorylation sites (Timofeeva et al., 2012). Recent studies show that STAT3 is involved in the regulatory network of embryonic stem cells (ESCs) and participate in LIF and BMP signaling pathways, which play important roles in self-renewal, reprogramming, and pluripotency in ESCs (Chen et al., 2008). Besides that, the phosphorylation of STAT3 in the mesenchymal stem cells (MSCs) upregulates the relevant genes of osteogenic differentiation (Nicolaidou et al., 2012).
It also has been proved that inflammatory factors can lead to the activation of STAT3 significantly and upregulate the expression of miR-21 (Zhang et al., 2016). Furthermore, miR-21 can bind with STAT3, and there is a positive feedback loop between STAT3 and miR-21 (Huang et al., 2016). Dentin, the same as bone, is a mineralizing tissue, which includes the expression of mineralization-related genes, such as alkaline phosphatase (ALP), dentin sialophosphoprotein (DSPP), and dentin matrix acidic phosphoprotein 1 (DMP1), in its development, and the osteogenic and odontoblast differentiation both lead to ALP-positive cell staining and Alizarin red staining positively (Chaudhary et al., 2016; Chen et al., 2015). Nevertheless, the role of miR-21/STAT3 signal in odontoblast differentiation has not been reported.
In this study, we analyzed the regulation of miR-21/STAT3 signal on odontoblast differentiation of DPSCs in inflammation microenvironment constructed by TNF-α. Our findings may provide a novel view to treat bone or tooth defect.
Materials and Methods
Isolation, culture, and chemical treatment of DPSCs
Extracted normal human impacted third molars completely were collected from patients (18–28 years of age, n = 15) with informed consent of patients who were undergoing treatment at the Dental Department of the Affiliated Hospital of Nantong University, which were approved by the Ethics Committee of the Affiliated Hospital of Nantong University. The Ethics Committee's approval number is 2016–077. All subjects had neither carious lesions nor any other oral infection.
The pulp tissues were separated from the crown and root completely and then immersed into the following digestive solution: 3 mg/mL type I collagenase, 4 mg/mL dispase in 4 mL phosphate-buffered saline (PBS), and 100 U/mL penicillin with 100 μg/mL streptomycin for 1 hour at 37°C. Single-cell suspensions were obtained by passing the digested tissues through a 70-μm cell strainer (BD Falcon). Cell suspensions of dental pulp were put into 25 cm2 culture dishes and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in 5% CO2 atmosphere. The medium was replaced every 3 days. Cells were passaged at the ratio of 1:3, when they reached 85%–90% confluence.
Cells from the third passage were used for all experiments. The inflammation microenvironment was produced with TNF-α (Sigma) and formulated the final concentration (1–100 ng/mL).
Cucurbitacin I (Cuc I) (Indofin, Hillsborough, NJ) was dissolved in dimethyl sulfoxide (DMSO) at a concentration of 50 mM and diluted in culture medium to a final concentration of 30 μM.
MTT assay
DPSCs were seeded on 96-well plates at a cell density of 5 9 103 cells/well. At 24, 48, and 72 hours after the TNF-a treatment, the proliferation/survival of the cells was evaluated using the methylthiazolyldiphenyl-tetrazolium bromide (MTT) test. In brief, 20 mL MTT (5 g/L; Beyotime, Shanghai, China) was added to each well, and the cells were incubated for 4 hours at 37°C. The precipitate was extracted with DMSO (Sigma), and the optical density was measured at the wavelength of 490 nm.
Alizarin red S and ALP staining
DPSCs were fixed with 4% paraformaldehyde (PFA) for 1 hour and washed with PBS. Cells were then stained with 40 mM Alizarin red S (pH = 4.2) for 10 minutes under conditions of gentle agitation. Absorbance of the extracted Alizarin red S (Sigma) stain was measured at 570 nm. DPSCs were subjected to the ALP (SenBeiJia, Nanjing, China) using the ALP Assay Kit according to the manufacturer's instructions.
Western blot
Cells were lysed in buffer consisting of 50 mM TRIS, 150 mM NaCl, 2% sodium dodecyl sulfate (SDS), and a protease inhibitor mixture. After centrifugation at 12,000 rpm for 12 minutes, protein concentrations were determined using the Bradford assay (Bio-Rad). The resulting supernatant (50 μg of protein) was subjected to SDS-polyacrylamide gel electrophoresis (PAGE). The separated proteins were transferred onto PVDF membranes at 350 mA for 2.5 hours in a blotting apparatus (Bio-Rad, CA). Membranes were blocked with 5% nonfat milk and incubated with primary antibodies (1:400) at 4°C overnight and subsequently with anti-rabbit horseradish peroxidase-conjugated secondary antibodies (1:1000) for 2 hours at room temperature.
Concomitantly, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was run as a reference protein. The following primary antibodies were used: GAPDH (anti-mouse; Santa Cruz), DMP1 (anti-mouse; Sigma), DSPP (anti-mouse; Sigma), STAT3 (anti-mouse; Santa Cruz), p-STAT3-Ser727 (anti-mouse; Cell Signaling), and β-tubulin (anti-mouse; Cell Signaling).
Quantitative reverse transcriptase PCR
Expression levels of pri-miR-21 were assayed in total RNA samples isolated from 1 to 10 × 106 cells using the RNeasy Mini kit (Qiagen, Hilden, Germany). RNA (1 μg) was treated with RQ1 DNase I (Promega, Madison, WI) and converted into complementary DNA using IScript reverse transcriptase (Bio-Rad, Hemel Hempstead, United Kingdom), oligo(dT)12–18, and random hexamer primers (Bio-Rad). Real-time PCR was performed using the IQ5 instrument and SYBR Green Supermix (Bio-Rad), using the following cycle parameters: 5 minutes at 95°C, followed by 45 cycles of 15 seconds at 97°C and 30 seconds at 60°C. Primer sequences for pri-miR-21 and RPS11 have been described previously (Loffler et al., 2007; van der Fits et al., 2011; Vermeer et al., 2008). Data were analyzed using IQ5 software using the ΔΔCt method (Bio-Rad), and expressed relative to RPS11 expression levels. Specificity of the PCR products was confirmed by melting curve analysis.
For determining mature miR-21 expression levels, total RNA was isolated from 1 to 10 × 106 cells using the mirVana RNA Isolation Kit (Ambion, Austin, TX). Total RNA (150 ng) was used for reverse transcriptase and subsequent preamplification reaction using Megaplex Primer pool A, according to the manufacturer's protocol (Applied Biosystems, Foster City, CA). The miRNA real-time PCR was performed for miR-21, U6 using the TaqMan miRNA assays and Universal PCR Master Mix (Applied Biosystems) on an IQ5 instrument (Bio-Rad), according to the manufacturer's protocol (Applied Biosystems). The cycle parameters were as follows: 10 minutes at 95°C, followed by 40 cycles of 15 seconds at 95°C and 60 seconds at 60°C. MiR-21 expression was analyzed using the ΔΔCt method in the IQ5 software (Bio-Rad), and expressed relative to U6 expression levels.
Immunofluorescent staining
DPSCs were fixed with 4% PFA for 1 hour, washed with PBS containing 0.1% Triton X-100 (PBST), and blocked for 30 minutes in PBST supplemented with 10% FBS. Cells were then incubated with one of the following primary antibodies (1:100) in the same solution overnight at 4°C: p-STAT3 (anti-mouse; Sigma). Cells were then washed and incubated in secondary antibodies for 2 hours at room temperature. Nuclei were stained with DAPI (4′6′-diamidino-2-phenylindole dihydrochloride) (1:800; Santa Cruz). The cells were examined using a Leica fluorescence microscope (Germany).
Nuclear and cytoplasmic extraction
After treated with PBS, DPSCs seeded in plates were washed with ice-cold PBS, followed by suspension in 100 μL of hypotonic buffer (10 mM HEPES, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM DTT, 1 μL of 10% NP-40, 1 mM PMSF, and a protease inhibitor cocktail). The DPSCs placed on the ice were allowed to swell for 10 minutes. The DPSCs suspension was subsequently vibrated on a vortex for 10 seconds and then centrifuged at 14,000 g for 2 minutes at 4°C. The resulting supernatant was regarded as the cytosolic extract. The remaining pellet containing the nucleus was washed with PBS and then resuspended in 50 μL of ice-cold nuclear extraction buffer [pH 7.9] (20 mM HEPES, 400 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 0.5 mM PMSF, and a protease inhibitor cocktail).
Similarly, the suspension was vibrated on a vortex for 10 seconds and incubated on ice for 1 hour with intermittent vortexing. At last the suspension treated was centrifuged at 14,000 g for 10 minutes and the resulting supernatant was regarded as the nuclear fraction. The protein samples harvested in the experiment were stored at −80°C before use.
SiRNAs and transfection
SiRNA transfection was carried out using a commercially available kit (GENECHEM). For siRNA inhibition studies, DPSCs were washed with the siRNA transfection medium and then incubated (at 37°C and 5% CO2) for 12 hours with transfection medium containing the transfection reagent and either STAT3 siRNA (50 nM) or control siRNA (50 nM), according to the manufacturer's instructions. After transfection, the cells were harvested at 72 hours for RNA or protein extraction.
Chromatin immunoprecipitation
ChIP was performed using the EZ-ChIP kit (Upstate, Temecula, CA), according to the manufacturer's instruction. In short, DPSCs were grown overnight in medium deprived of cytokines. Cells were incubated with or without TNF-α (10 ng/mL) for 30 minutes at 37°C, immediately fixed in formaldehyde, and chromatin was sonicated using the Bioruptor (Diagenode, Liege, Belgium). Immunoprecipitation was performed using anti-STAT3 antibodies (Santa Cruz Biotechnology, Santa Cruz, CA), or with control antibodies supplied in the ChIP kit (anti-acetyl-Histone H3, or normal rabbit IgG). Coimmunoprecipitated DNA was amplified using primers for the GAPDH promoter, as supplied in the ChIP kit, or primers specific for the upstream region of the miR-21 gene (Loffler et al., 2007).
Real-time PCR was performed using the IQ5 instrument and SYBR Green Supermix (Bio-Rad), using the following cycle parameters: 6 minutes at 95°C, followed by 45 cycles of 15 seconds at 96°C, 30 seconds at 57°C, and 30 seconds at 72°C. Data were analyzed using IQ5 software using the ΔCt method (Bio-Rad). Specificity of the PCR products was confirmed by melting curve analysis.
Transfection experiments
DPSCs were transferred to medium without penicillin and streptomycin. Next day, cells were transfected by Lipofectamine 2000 (Invitrogen), with 100 nM fluorescently labeled oligonucleotides (BLOCK-iT, Invitrogen) combined with 40 nM anti-miR-21 miRNA inhibitor (no. AM10206; Ambion) or negative control (negative control no.1: AM17010; Ambion). At 7 days after transfection, DPSCs were harvested.
Statistical analysis
The data were analyzed and are expressed as the mean ± standard deviation. Differences between groups were tested for statistical significance using ANOVA. Student's t-test was used to determine significance with SPSS 17.0 software. A value for p < 0.05 was considered as statistically significant. Each of the experiments was repeated for three times.
Results
TNF-α influenced the odontoblast differentiation of DPSCs
TNF-α promoted the expression of DMP1 and DSPP in low concentration (1, 10 ng/mL), but the odontoblast differentiation potential of DPSCs was significantly inhibited in high concentration of 50 and 100 ng/mL (Fig. 1A). Alizarin red S staining and ALP staining also proved the same results (Fig. 1B, C). The best concentration to odontoblast was considered to be 10 ng/mL TNF-α, and so, 10 ng/mL TNF-α was chosen in the next experiments. The survival of DPSCs did not change significantly in different concentration of TNF-α (Fig. 1D). These data proved that suitable inflammation microenvironment had the positive influence during the odontoblast differentiation of DPSCs, and TNF-α did not influence the survival of DPSCs.

TNF-α influenced the odontoblast differentiation of DPSCs.
STAT3 activation and translocated into nucleus during odontoblast differentiation of DPSCs stimulated by TNF-α
The increasing expression of STAT3 in the nucleus and p-STAT3 can be observed in low concentration of TNF-α group (1, 10 ng/mL), while the contrary expression of STAT3 was shown in the cytoplasm (Fig. 2A). The immunocytochemistry of STAT3 in the nucleus of stimulation group (TNF-α, 10 ng/mL) had higher intensity compared with that in the control group (Fig. 2B). These data showed that STAT3 was activated and translocated into nucleus during odontoblast differentiation of DPSCs stimulated by TNF-α.

STAT3 was activated and translocated into nucleus during odontoblast differentiation of DPSCs stimulated by TNF-α.
TNF-α influenced the expression of miR-21 during odontoblast differentiation of DPSCs and an interaction existed in STAT3 and miR-21
Quantitative reverse transcriptase PCR (RT-PCR) was used to analyze RNA of DPSCs stimulated by TNF-α with different concentration for 7 days, and the results showed that the expression of the miR-21 increased with the same tendency as DMP1, DSPP, and STAT3 in the nucleus (Fig. 3A). Furthermore, the expression of the miR-21 in DPSCs stimulated by TNF-α (10 ng/mL) has increased following the time of culture (Fig. 3B).
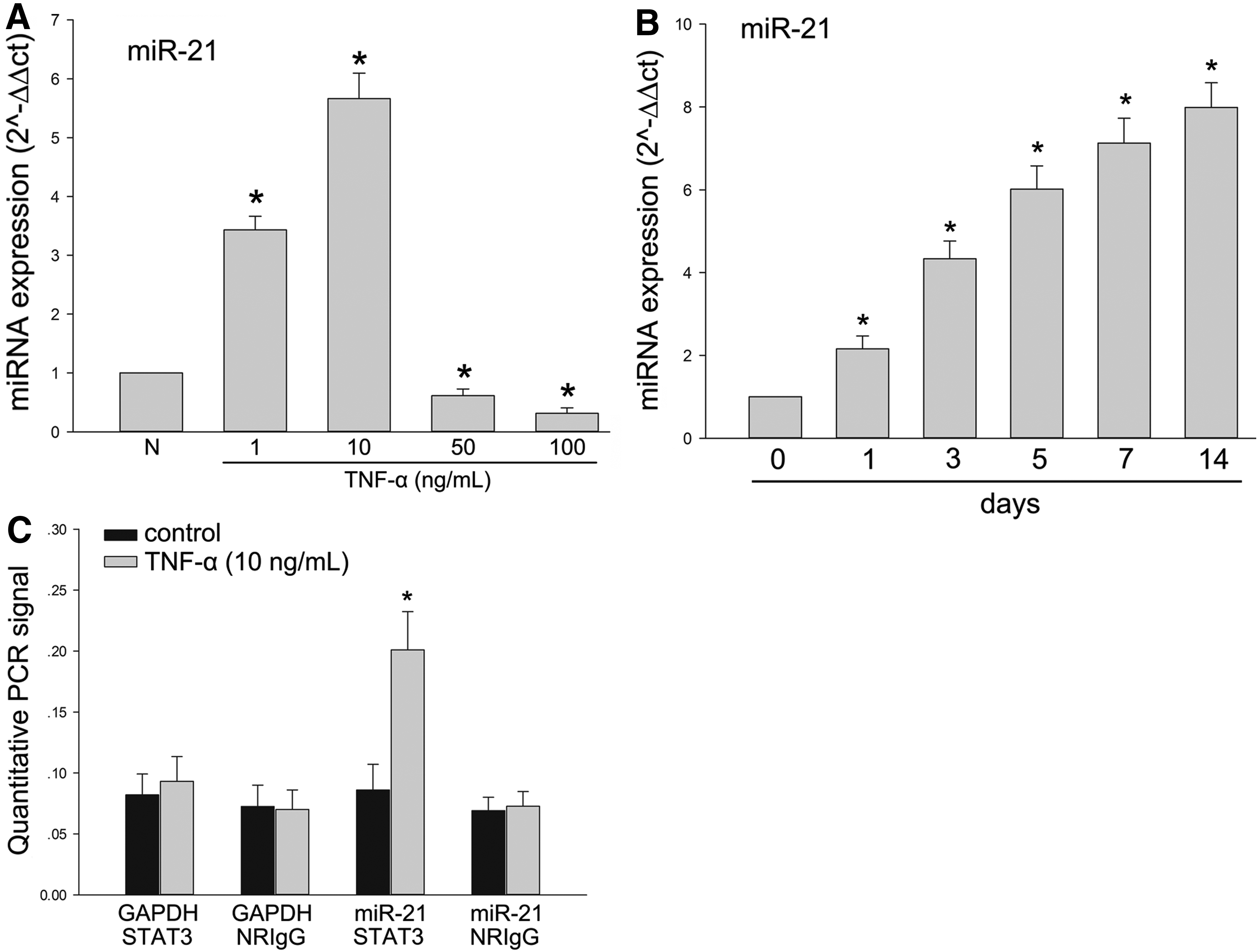
TNF-α influenced the expression of miR-21 during odontoblast differentiation of DPSCs and an interaction existed in STAT3 and miR-21.
DPSCs were treated with TNF-α (10 ng/mL) for 7 days to activate STAT3 and analyzed by chromatin immunoprecipitation (ChIP). Chromatin that was precipitated with STAT3 antibodies was significantly enriched for the miR-21 promoter sequence when compared with chromatin precipitated with normal rabbit IgG (Fig. 3C). In contrast, without stimulation of TNF-α, the changing has not been detected (Fig. 3C). The GAPDH promoter was observed in DPSCs cultured with or without TNF-α (10 ng/mL) as a control, and there was no precipitation demonstrated with the ChIP (Fig. 3C).
A positive feedback loop between STAT3 and miR-21
To prove STAT3 is involved in the expression of miR-21 directly during odontoblast differentiation, the expression of STAT3 was interfered by siRNA and cucurbitacin I (Cuc I), an inhibitor of the STAT3 signaling. Western blot was performed to prove the silencing efficiency of siRNA for STAT3 (Fig. 4A). Furthermore, RT-PCR analysis of the expression of primary miR-21 (pri-miR-21) has significantly increased in siSTAT3 and Cuc I group compared with control group, while the reverse results were detected about the expression of mature miR-21 (Fig. 4B, C). Western blot showed that the expressions of DMP1 and DSPP had a significant decreasing when transfected with siSTAT3 or treated by Cuc I (Fig. 4D).

A positive feedback loop between STAT3 and miR-21.
MiR-21 expression was knocked down in DPSCs by transfecting with anti-miR-21 oligonucleotides and analyzed the odontoblast differentiation and the activation of STAT3. Comparing with DPSCs transfected with fluorescently labeled oligonucleotides only as a control, transfection of anti-miR-21 led to the decreasing activation of STAT3 and expression of DMP1 and DSPP (Fig. 4E). The DPSCs were cultured with TNF-α (10 ng/mL) for 7 days (Fig. 4B–E).
Discussion
Our experiments demonstrated that the miR-21, as a positive role, was involved in odontoblast differentiation of DPSCs treated by TNF-α. Furthermore, there is a positive reciprocal loop between miR-21 and STAT3. Stimulated by TNF-α of the most appropriate concentration (10 ng/mL), the increasing odontoblast differentiation along with the same changing of the maker proteins, such as DMP1 and DSPP, has been found. SiSTAT3 or treated by cucurbitacin I (Cuc I) significantly increased pri-miR-21 expression (Fig. 4B) and decreased miR-21 expression (Fig. 4C). Meanwhile, the inhibition of miR-21 (anti-miR-21) decreased the activation of STAT3 as well as the maker proteins of odontoblast differentiation (Fig. 4E). The interaction happened in the inflammation microenvironment between miR-21 and STAT3 influenced the odontoblast differentiation of DPSCs.
As a type of tool cell of tissue engineering and regenerative medicine, DPSCs were applied widely in various fields (Chen et al., 2016), including many oral diseases such as pulpitis, periodontitis, and healing process of bone fracture. As a kind of common factor of treatment, DPSCs also influence the pulp regeneration (Zhu et al., 2012). TNF-α, LPS (lipopolysaccharide, LPS), and family of IL (interleukin, IL) were observed normally in above diseases and played a vital role about resorption and reconstruction of bone and dentin. Previous studies prove that DPSCs have the ability to differentiate to specific cell lineages in inflammation microenvironment (Feng et al., 2015; Xing et al., 2015). Furthermore, obtaining DPSCs is relatively easier and can be used frequently.
TNF-α has been reported to trigger interactions between invading monocytes and vascular endothelial cells, which subsequently induce endothelial apoptosis in the circulation. In this model, the human umbilical vein endothelial cells have been impaired by stimulation of TNF-α. TNF-α treatment resulted in additional time and dose-dependent decreases in cell viability in treated cells compared with that of control cells. High-concentration (12.5–200 ng/mL) TNF-α reduced cell viability (Zhou et al., 2017). A high TNF-α concentration (20–40 ng/mL) also induced apoptosis in endothelial progenitor cells more obviously (Du et al., 2014). In acute lung injury model, inhibiting the expression of TNF-α can alleviate vascular leakage and tissue damage of the lung (Weifeng et al., 2016). In nucleus pulposus inflammatory environment, a high TNF-α concentration (20–40 ng/mL) produced greater effects on proliferation and senescence than a low TNF-α concentration (10 ng/mL) (Li et al., 2017).
In our experiment, we established the model of inflammation microenvironment with TNF-α, and different concentrations (0, 1, 10, 50, 100 ng/mL) were used individually. The results were similar with previous studies that the low concentrations (10 ng/mL) could promote osteogenic/odontoblast differentiation of DPSCs, and the reverse results were shown in higher concentrations (100 ng/mL) (Fig. 1) (Feng et al., 2013).
STAT3 is a latent transcription factor that mediates extracellular signals such as cytokines and growth factors via interaction with polypeptide receptors at the cell surface (Levy and Darnell, 2002). After STAT3 protein becomes activated, the activated protein (p-STAT3) translocates to the nucleus and binds to sequence-specific DNA elements for transcription of target genes (Akira et al., 1994). It has been reported that MSCs, which infected with recombinant adenovirus vector Ad-let-7a and cocultured with rat pulmonary artery smooth muscle cells, restrained the activation of STAT3 and increased its downstream bone morphogenetic protein receptor 2 (BMPR2) expression (Cheng et al., 2017).
STAT3, acts as a key regulator, participated in many physiological and pathological processes, including inflammation (Fu et al., 2016). STAT3 can be phosphorylated by stimulation of inflammatory factor, IL-6, and initiates the downstream biological function after translocating into the nucleus as showed in Figure 2. It has been proved that STAT3 played a vital role in process of odontoblast differentiation of DPSCs (Vandomme et al., 2014). There is still a long way to go to investigate the application of STAT3-related signaling in human therapy, which will be the focus of our future research.
Meanwhile, microRNAs, such as miR-21, were involved in almost all biological process in which inflammatory has also been contained (Garo and Murugaiyan, 2016). Previous experiments indicated that miR-21 had a strong connection with osteogenic differentiation of MSCs (Yang et al., 2013). Earlier studies had found that osteoblastic bone formation occurred via the upregulation of expression of miR-21 in MSCs with the enhanced expression of BMP9 (Song et al., 2015). Furthermore, the interaction between miR-21 and STAT3 has also been reported, which revealed a positive feedback loop that contributed to atrial fibrosis (Huang et al., 2016), while fibrosis would be inhibited by suppressing STAT3/miR-21 (Zhou et al., 2015). The fibrosis is regarded as the beginning of ossification commonly (Martinaud et al., 2015).
Correspondingly, our results also exhibited the same positive feedback loop that contributed the odontoblast differentiation stimulated by TNF-α (10 ng/mL) (Fig. 4). In addition, the interaction of miR-21/STAT3 has been discovered in cancer models widely. The activation of STAT3, stimulated by IL-21 in Sézary cells, resulted in upregulation of miR-21 expression (van der Fits et al., 2011). Although the multiple findings about the miR-21/STAT3 interactions in various disease models have been reported, further studies should be taken to pay more attention to the application of miR-engineering for the treatment of human diseases.
In summary, it is demonstrated that STAT3 was activated by stimulation of TNF-α and regulated miR-21 mutually in DPSCs. Afterward, the miR-21/STAT3 signal positively led to the increasing expression of downstream proteins of odontoblast differentiation (Fig. 5). The more the findings of miRs and proteins about tissue repairing, the more the methods we can use in tissue engineering and regenerative medicine.
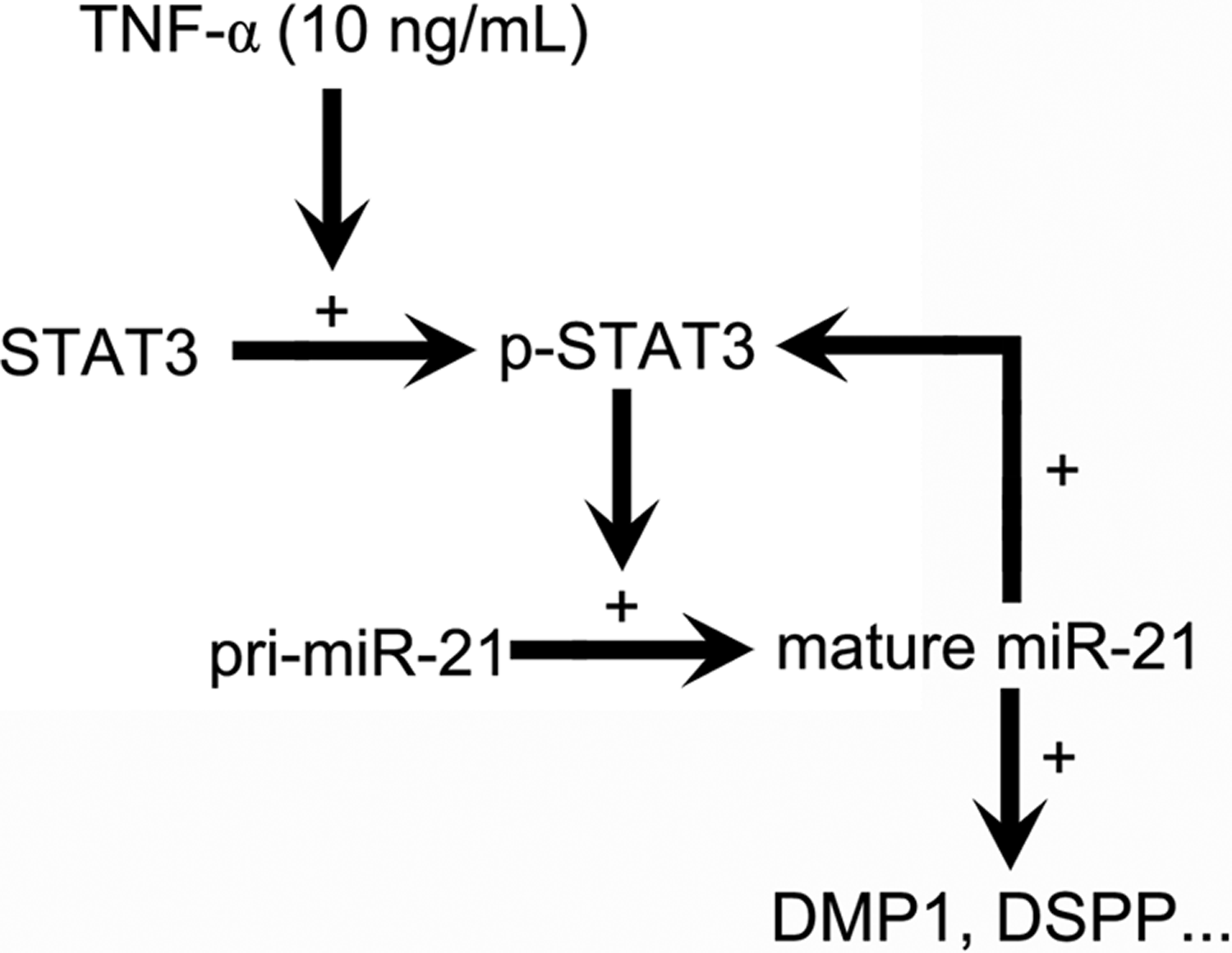
Schematic diagram summarizes our results. A positive feedback loop between p-STAT3 and mature miR-21 mediates the odontoblast differentiation of DPSCs. +, promoting activity, or expression of downstream molecule.
Footnotes
Acknowledgments
This work was supported by Natural Science Foundation of China Grant (nos. 81500809, 81501076), Jiangsu Natural Science Foundation (BK2011385), Nantong City Science and Technology Projects Funds (MS22016049), “Top Six Types of Talents” Financial Assistance of Jiangsu Province Grant (No. 2013-WSW-048), Graduate Student Innovation of Science and Technology Projects in Jiangsu Province in Nantong University ![]() and Nantong Natural Science Foundation (no. BK2014038), Nantong Health Bureau Youth Foundation of China (WQ2015016).
and Nantong Natural Science Foundation (no. BK2014038), Nantong Health Bureau Youth Foundation of China (WQ2015016).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.