
Abstract
Abstract
Cancer stem cells (CSCs) are responsible for cancer initiating, recurrence, and drug resistance. Discovery of novel biomarkers for CSCs is helpful for early diagnosis and prognosis. Lung cancer stem cells (LCSCs) were closely related to the occurrence and development of lung cancer. In our study, the important role of miR-2861 in maintaining the stemness of LCSCs was investigated. The LCSC differentiation model was established through introducing serum into the medium of H460 spheres. miR-2861 expression was significantly higher in LCSCs no matter compared to the differentiation cells or normal cells. HDAC5 expression was positively correlated with miR-2861 in LCSCs, and knockdown of miR-2861 decreased the expression of HDAC5, which implied that HDAC5 may be involved in the differentiation of LCSCs mediated by miR-2861. The role of HDAC5 in the regulation of LCSC differentiation was further verified by the inhibitory effect of LMK-235 on the phosphorylation of ERK1/2, which was recognized as the regulator of CSC differentiation. Our study provided a better understanding of miR-2861 and HDAC5 axis in maintaining the stemness of LCSCs and laid a foundation for molecular targeted therapy.
Introduction
C
Normally, CSCs are identified through surface markers, such as CD44 for colorectal CSCs and gastric CSCs (Dalerba et al., 2007; Lee et al., 2008; Takaishi et al., 2009) and CD133 for liver CSCs and ovarian CSCs (Curley et al., 2009; Ma et al., 2008). However, the specificities of these biomarkers are not high. Discovery of novel biomarkers for CSCs may be helpful for improving early diagnosis and prognosis.
Micro RNAs (miRNAs) are a class of small noncoding RNAs that control gene expression in diverse organisms (Tsuchiya et al., 2009). In some cases, miRNAs can imprecisely pair with the complementary sites and interfere with the translation of proteins. In other cases, miRNAs can perfectly pair with the complementary sites and make their targets degraded (Khvorova et al., 2003). Recently, miRNAs were reported to be aberrantly expressed in human cancers and have some relationship with the development of CSCs (Dahiya et al., 2008). For example, miR-200b was reported to be highly expressed in CD44 identified pancreatic CSCs (Bao et al., 2011). MiR-142-3p was reported to be poorly expressed in CD133 identified liver CSCs (Chai et al., 2014). These evidences implied that miRNAs may serve as a novel class of CSC biomarkers.
miRNAs are reported to regulate the proliferation and differentiation of CSCs through ERK, AKT, and NOTCH signaling pathways. The activity of NOTCH was elevated by the increased expression of miR-18a*, and activated NOTCH in turn was required for sustained ERK activation, suggesting that miR-18a*/NOTCH/ERK loop was essential for maintaining the stemness of glial CSCs (Turchi et al., 2013). miR-330 contributed to the deterioration of glial CSCs by activating ERK and PI3K/AKT signaling pathways (Yao et al., 2014). miR-18a was responsible for the regulation of PI3K/AKT and ERK pathways, which associated with the proliferation of pancreatic CSCs (Li et al., 2017). More and deeper studies need to be performed on the discovery of more unknown miRNAs and the exploration of functional mechanisms on how these miRNAs regulated the stemness and differentiation of CSCs imminently.
Wang et al. (2014) reported that miR-2861 was highly expressed in lung cancer stem cells (LCSCs). However, the signaling pathways in which miR-2861 was involved in regulating the stemness and differentiation of LCSCs remained unknown. In our study, the LCSC differentiation model would be established to explore the correlation between the expression of miR-2861 and the stemness of LCSCs. The antisense oligonucleotides of miR-2861 or other related inhibitors would be applied to identify the possible signal molecules or the downstream molecules in miR-2861 mediating pathways. Overall, we hope that our study could expand the knowledge of CSCs, especially the effect of miRNAs on the regulation of CSC differentiation.
Materials and Methods
Cell lines and culture
Human lung cancer cell lines (H460 cells and H1299 cells) were purchased from Cell Bank of the Chinese Academy of Science (Shanghai, China). Normal human mammary epithelial cell line (MCF-10A cells) was kindly provided by Suo (Suzhou Institute of Nano-Tech and Nano-Bionics, Chinese Academy of Sciences, Suzhou, China). Mesenchymal stem cells (MSCs) were kindly provided by Yang (Suzhou Institute of Nano-Tech and Nano-Bionics, Chinese Academy of Sciences, Suzhou, China).
H460 cells and H1299 cells were cultured in RPMI-1640 (Hyclone) supplemented with 10% fetal bovine serum (FBS; Hyclone). H460 spheres and H1299 spheres were cultured at a density of 2.5 × 105 cells/mL in the Dulbecco's modified Eagle's medium (DMEM)-F12 (Hyclone) supplemented with 20 ng/mL basic fibroblast growth factor (bFGF; PeproTech), 20 ng/mL epidermal growth factor (EGF; PeproTech), and 2% B27 (Gibco). Adherent cells were harvested when they were subconfluent, and tumor spheres were harvested every 6 days. All media were supplemented with 1% penicillin/streptomycin (Hyclone). Cells were maintained in a 5% CO2 humidified incubator at 37°C.
Differentiation assay of tumor spheres
Six days after serum-free medium culture, H460 spheres were cultured in PRIM 1640 medium supplemented with 10% FBS in a six-well culture plate for various durations. The H460 spheres which were cultured in serum medium for 2, 4, 6, and 8 days were collected, and the gene expression was measured using quantitative real-time polymerase chain reaction (qRT-PCR).
RNA extraction
Total RNA was extracted from each cell line using TRIzol reagent (Invitrogen) according to the method of acid guanidinium thiocyanate-phenol-chloroform extraction. RNA was examined and quantified by agarose electrophoresis and NanoDrop 2000c (Thermo). Total RNA (1 μg) from each sample was reverse transcribed into complementary DNA (cDNA) using PrimeScript™ RT reagent Kit (TaKaRa) with oligo (dt) or special stem-loop primers listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/cell).
Quantitative real-time PCR
Human small nuclear U6 and actin were used as the internal reference for the normalization of miRNAs and mRNA. The primers for qRT-PCR were designed based on NCBI database, using Primer Premier 5 and Oligo 6. qRT-PCR was performed on an ABI 7500 (Applied Biosystems) using SYBR Premix Ex Taq™ II (TaKaRa). Reactions were carried out at 95°C for 30 seconds as an initial activation followed by 40 cycles at 95°C for 5 seconds and 60°C for 30 seconds. Melting curves were generated for each qRT-PCR to verify the specificity. All the reactions were performed in triplicate, and relative expression levels of mRNA were calculated by the equation 2−ΔΔCt. The sequences of the primers used in qRT-PCR are listed in Supplementary Table S2.
Flow cytometry
The binding ability of HCBP-1 peptide to spheres or subtypes was analyzed by flow cytometry on an Accuri™ C6 by the manufacturer's recommendations (BD Biosciences). Cells were resuspended at 1 × 106 cells per 100 μL phosphate-buffered saline (PBS) containing HCBP-1 fluorescent dye-labeled peptide (Aladdin) and incubated for 30 minutes at 4°C. Cells treated with scribble peptide were used as negative control. Following extensive washing, the cells were resuspended in 400 μL of PBS for flow analysis or sorting.
Knockdown of miR-2861
A 21-nucleotide probe anti-miR-2861 (5′-CCGCCCGCCGCCAGGCCCCtt-3′) and a nucleotide probe anti-miR-ctrl (5′-UUGUACUACACAAAAGUACUG-3′) with no significant homology to any known gene sequence were purchased from GenePharma (Shanghai, China). Tumor spheres were transiently transfected with anti-miR-2861 or anti-miR-ctrl using Gene Pulser Xcell™ (Bio-Rad) and Lipofectamine 3000 (Invitrogen) sequentially according to the manufacturer's protocol. The conditions used for optimal silencing were as follows: 2 × 106 cells transfected with anti-miR-2861 or anti-miR-ctrl at a final concentration of 10 nM for 155 V about 30 mseconds and then transfected with anti-miR-2861 or anti-miR-ctrl at a final concentration of 100 nM in 7.5 μL Lipofectamine 3000. At 48 or 72 hours post-transfection, cells were harvested for total RNA and protein extraction.
LMK-235 treatment
H460 spheres were seeded in six-well plates, 6 days after culture, LMK-235 (Biotool) was dissolved in dimethyl sulfoxide (DMSO) and its final concentration was in the range of 0.5–50.0 μM, and the medium containing 0.1% DMSO was used as control. Cells were cultured for 2.5 hours in serum-free DMEM-F12 medium under the standard incubation condition, and the expression of protein was measured by Western blot.
Western blot analysis
Cells were washed twice with PBS and lysed with NP40 Cell Lysis Buffer (Invitrogen), including 1 × PMSF and 1 × phosphatase inhibitors (Biotool). The protein concentrations of the whole cell lysates were determined using a bicinchoninic acid (BCA) assay. Protein samples were boiled for 5 minutes in sodium dodecyl sulfate (SDS) protein buffer containing 1% mercaptoethanol. Proteins were separated by SDS-polyacrylamide gel electrophoresis (PAGE) and transferred to a PVDF membrane (Bio-Rad). The membrane was blocked with 5% skim milk in Tris-buffered saline (TBS) for 2 hours at room temperature and washed thrice with TBS for every 5 minutes. The membrane was incubated with primary antibody overnight at 4°C followed by incubation with the secondary peroxidase conjugated antibodies for 2 hours at room temperature.
Statistical analysis
Each experiment was performed at least thrice. Statistical calculations were determined by Student's t test using GraphPad Prism 6.0. Data were shown as mean ± standard deviation. p < 0.05 was considered statistically significant.
Results
The differentiation model of LCSCs was established
To verify the stemness of H460 spheres, the expression of NANOG in H460, H460 spheres, H1299, and H1299 spheres was measured by qRT-PCR. NANOG was highly expressed in H460 spheres and H1299 spheres compared to their parental cells (Supplementary Fig. S1), which meant that spheres had the traits of stem cells and could work as the ideal candidate for the study of CSCs.
To identify possible signal pathways and important signal molecules that may be involved in the differentiation of LCSCs, the differentiation model of LCSCs needs to be established. In our study, we attempted to set up LCSC differentiation model through introducing serum into the medium of H460 spheres. It is well known that there are many growth factors or other factors such as insulin in serum, which could induce the differentiation of cells. After H460 spheres were cultured in full serum medium for more than 2 days, the morphology of cells changed from round shape to epithelial-like shape (Fig. 1A).

The LCSC differentiation model was identified.
To validate the cell types that serum-induced H460 spheres could generate into, the qRT-PCR and flow cytometric analyses were performed. The expression levels of NANOG and OCT4 decreased gradually along with the extension of culturing time with full serum medium (Fig. 1B, C). HCBP-1 peptide was specially binding with H460 spheres, which was screened by Anxin Wang before (Wang et al., 2014). The data from flow cytometry showed that the mean green fluorescence intensity of whole populations gradually decreased with the extension of culturing time with full serum medium (Fig. 1D).
Our results indicated that the differentiation of LCSCs could be induced by serum. As an assumed marker of LCSC, the expression level of miR-2861 in H460 spheres cultured in full serum medium with prolonged durations was measured by qRT-PCR. The decreasing tendency of the miR-2861 expression in cells along with the prolonged full serum medium cultured duration was observed (Fig. 1E).
miR-2861 and HDAC5 were differentially expressed in various cells
To examine whether miR-2861 was only highly expressed in LCSCs, the expression of miR-2861 in human normal cell line MCF-10A, MSC and another non-small lung cancer line H1299 was measured by qRT-PCR. Our results indicated that miR-2861 was highly expressed in spheres no matter that their parental cells were H460 or H1299 (Fig. 2A, B), which indicated that miR-2861 was involved in maintaining the stemness of LCSCs.

miR-2861 and HDAC5 were differentially expressed in various cells.
HDAC5 was a target of miR-2861 in the process of cell differentiation and proliferation in osteoblasts (Hu et al., 2011; Xia et al., 2015) and CHO cells (Fischer et al., 2015). To explore whether HDAC5 was involved in maintaining the stemness of LCSCs, the expression level of HDAC5 was examined in MCF-10A, H460, and H460 spheres. Results presented in Figure 2C indicated that HDAC5 was highly expressed in H460 spheres; the expression level of HDAC5 in MCF-10A and H460 was normal. HDAC5 and miR-2861 had the same expression tendency during the cell differentiation, which implied that HDAC5 may be involved in the regulation of LCSC differentiation.
The expression of HDAC5 in H460 spheres was decreased after cells were treated with anti-miR-2861
To examine the correlation between HDAC5 and miR-2861, the anti-miR-2861 was applied. Anti-miR-2861 in various concentrations was introduced into the medium of H460 spheres. MiR-2861 expression was decreased after cells were treated with anti-miR-2861 for 3 days (Fig. 3A), which suggested that anti-miR-2861 interfered with the expression of miR-2861. Both the mRNA expression level and the protein expression level of HDAC5 were dramatically decreased after cells were treated with anti-miR-2861 for 3 days (Fig. 3B–D), which suggested that expression of miR-2861 and HDAC5 was positively correlated. Meanwhile, anti-miR-2861 could significantly decrease the expression of NANOG and OCT4 (Supplementary Fig. S2), which further implied that HDAC5 may be involved in maintaining the stemness of LCSCs.
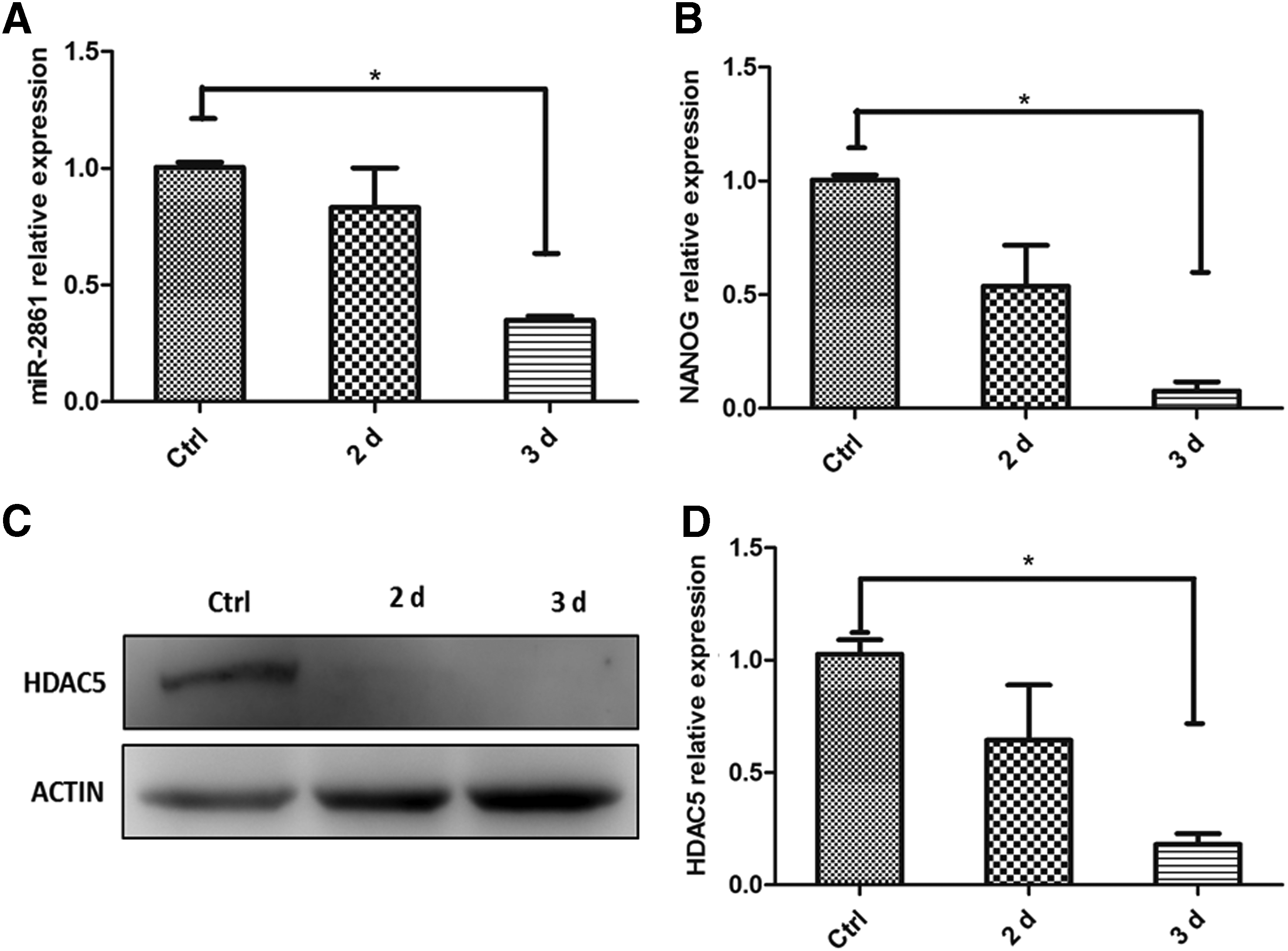
Anti-miR-2861 decreased the expression of HDAC5.
The phosphorylation level of ERK1/2 in H460 spheres was decreased after cells were treated with LMK-235
ERK1/2 participated in the differentiation of acute myeloid leukemia cells (Dai et al., 2016) and CSCs (Ahn et al., 2013; Navaraj et al., 2011; Xu et al., 2015). To explore the signaling pathways in which HDAC5 may be involved in the regulation of LCSC differentiation, the phosphorylation level of ERK1/2 was examined after cells were treated with various concentrations of HDAC5 inhibitor LMK-235. The phosphorylation level of ERK1/2 decreased gradually along with the increased concentrations of LMK-235 (Fig. 4A, B). Meanwhile, the expression of miR-2861 and NANOG decreased in LMK-235 treated cell (Supplementary Fig. S3), and the refractive index of cells became low and the diameter of cells grew slowly (Supplementary Fig. S4). These results demonstrated that LMK-235 could inactivate ERK1/2 signaling pathway in a dose-dependent manner.

LMK-235 decreased the phosphorylation level of ERK1/2.
Discussion
In our study, the relationship between miR-2861 and HDAC5 in the differentiation of LCSCs was established for the first time. MiR-2861 could maintain the stemness of LCSCs through HDAC5-ERK1/2 signal pathway. The higher expression of miR-2861 was observed in LCSCs. HDAC5 expression was decreased after treatment with anti-miR-2861 in H460 spheres. Moreover, the inhibition of HDAC5 by LMK-235 could compromise the phosphorylation level of ERK1/2 protein. Therefore, our study provided a better understanding of miR-2861 and HDAC5 axis in maintaining the stemness of LCSCs and laid a foundation for molecular targeted therapy.
Inducing the LCSC differentiation may be a new way to treat lung cancer. The establishment of LCSC differentiation model is vital in investigating the features of LCSCs. CSCs could differentiate into various cell types when treated with different factors; Zhang et al. (2013) reported that the small LCSCs could be induced by adipogenic differentiation. Azzi et al. (2011) showed that IL-15 induced the stable epithelial differentiation of renal CSCs generating a differentiated nontumorigenic cell population.
Most of CSCs are cultured in serum-free basic medium containing specific additional ingredients to keep cells in the morphology of sphere like (Misuno et al., 2013). After serum was introduced into the medium of spheres, cells would lose the sphere-like shape and attach the bottom of culture plate gradually. Through examining the expression level of NANOG, which was one of CSC markers, we believed that serum could induce the differentiation of LCSCs. The concentration of serum in the medium and the duration of full serum medium could determine the differentiation stage of spheres.
miR-2861 was highly expressed in LCSCs and distinctly expressed in various cells and in different stages of differentiation spheres, which implied that it might be a marker for LCSCs. The signaling pathways in which miR-2861 may be involved in the regulation of LCSC differentiation need to be explored further. According to our data and referring to other people's reports (Kwon et al., 2014; Weeks and Avkiran, 2015), the following molecule regulator network was proposed (Fig. 5). miR-2861 inhibited the expression of protein X (X represented one protein or a protein complex), and X could inhibit the expression of HDAC5. HDAC5 could promote the phosphorylation of ERK1/2 to regulate the differentiation of LCSCs (Varricchio et al., 2014).
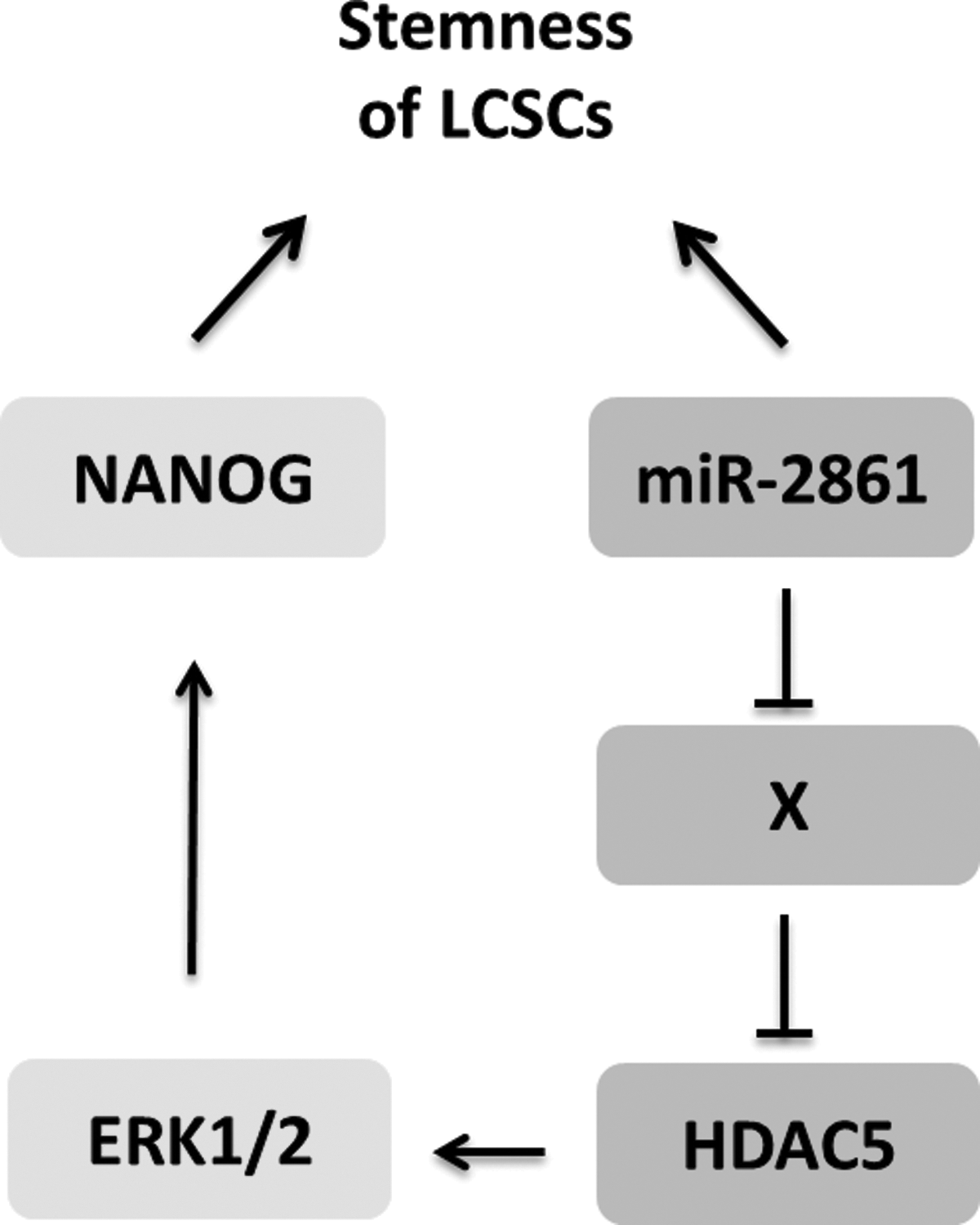
The molecule regulator network required for regulating the stemness of LCSCs.
According to previous studies, HDAC5 was the target of miR-2861 in the differentiation and proliferation of osteoblasts and CHO cells (Novo et al., 2013). However, we found that the expression of miR-2861 and HDAC5 was positively correlated in LCSCs, which implied that there were other molecules (X) existing to connect miR-2861 and HDAC5 to regulate the stemness of LCSCs. Many signal molecules could be the candidates for the X, such as vascular endothelial growth factor (VEGF). Ha et al. (2008) demonstrated that VEGF stimulated HDAC5 phosphorylation in endothelial cells, essential for vascular endothelial cell migration and angiogenesis. Li et al. (2016) believed that VEGF was related to maintain the stemness of CSCs. To summarize all those reports, VEGF possessed the traits as the candidate of the X. However, the detailed function of VEGF in the differentiation of LCSCs needs to be explored further.
HDACs are aberrantly expressed in various cancers and active in some signal pathways which regulate the differentiation of CSCs, such as Ras/Raf/MAPK (Khan and Jena, 2014; Marks et al., 2001; Rosato and Grant, 2003). In our study, HDAC5 was highly expressed in H460 spheres, and HDAC5 inhibitor decreased the phosphorylation level of ERK1/2 in LCSCs, which implied that HDAC5 was a potential target for cancer treatment and was required for the activation of ERK1/2 signal pathway. HDAC5 could promote histone acetylation and induce the relaxation of chromatin structure, leading to transcriptional activation of genes (Ozaki et al., 2006). However, it remained to be further investigated how HDAC5 functioned in regulating the phosphorylation level of ERK1/2.
In our study, the tendency of the phosphorylation level of ERK1/2 and the expression of NANOG was consistent in LCSCs, which implied that ERK1/2 may be involved in the maintenance of LCSC stemness. Kim et al. (2014) reported that activation of ERK could regulate self-renewal and differentiation of embryonic stem cells through the phosphorylation of NANOG. The article of Phiboonchaiyanan et al. (2016) showed that the activation of Cav-1 in lung cancer cells increased the phosphorylation level of ERK resulting in the increase of the NANOG expression, essential for the enhancement of self-renewal. Referring to these data, we believed that axis of miR-2861-HDAC5-ERK1/2-NANOG in maintaining the stemness of LCSCs did exist, even it may need more evidences to verify.
In conclusion, for the first time, we discovered and identified that miR-2861 could sustain the stemness of LCSCs through HDAC5-ERK1/2 signal pathway. Our findings greatly expanded the understanding of molecular mechanism of miR-2861 and HDACs in maintaining the stemness of LCSCs and provided new insights in targeted therapy.
Footnotes
Acknowledgments
This work was funded by the National Natural Science Foundation of China (No. 81372361, 81602552), Natural Science Foundation of Jiangsu province (No. BE2015647), and Science and Technology Bureau of Suzhou (No. SS201649).
Author Disclosure Statement
The authors declare there are no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.