
Abstract
Abstract
Buffalo (Bubalus bubalis) is a major source of milk, meat, and draught power in many developing countries in Asia. Animal cloning holds a lot of potential for fast multiplication of elite buffaloes and conservation of their valuable germplasm. Although the progress of buffalo cloning has been slow in comparison to cattle or pig, several breakthroughs were reported in buffalo cloning such as the production of cloned calves from somatic cells isolated from over one-decade old frozen–thawed semen or from urine-derived cells. Since the initiation of buffalo cloning, several approaches have been tried to refine nuclear transfer protocols. This has resulted in increasing the blastocyst production rate and improving their quality leading to an increase in live birth rate. In this review, we discuss current developments in buffalo cloning, its challenges, and the future roadmap.
Buffalo: An Animal with Great Potential But Limited Success in Application of Reproductive Technologies
D
After dairy cattle, buffalo is the second most important livestock in the world in terms of milk production (FAOSTAT, 2015). In recent years, buffalo husbandry has expanded widely in the Mediterranean, Latin America, and Central/Northern Europe, where several herds have been introduced (Warriach et al., 2015). India is the largest milk producer in the world, with production of about 155 million metric tons during 2015–2016, out of which nearly 55% were contributed by buffalo (Annual report 2016–2017 of Department of Animal Husbandry, Dairying & Fisheries, Government of India, http://dahd.nic.in).
Reproductive technologies such as superovulation and embryo transfer technology, Ovum Pick-Up and in vitro fertilization, semen sexing, stem cells, and transgenesis have not been developed in buffalo to the extent as they have been developed and successful in other farm animal species such as cattle and pig (Drost 2007; Singh et al., 2009). Animal cloning has emerged as the technique for multiplication and conservation of many animal species and a tool for transgenic animal production. We had initiated cloning in India with the aim of multiplying elite male and female buffaloes for dissemination of quality germplasm in the country to mitigate the ever growing demand for milk and meat (Singla et al., 2015). In addition to us, other laboratories in China, Thailand, and Italy have also reported buffalo cloning (Lagutina et al., 2011; Shi et al., 2007; Srirattana et al., 2014; Sun et al., 2015; Tasripoo et al., 2014).
Over the last decade, several buffalo clones have been produced in the world (Selokar et al., 2014; Shi et al., 2007; Singla et al., 2015; Tasripoo et al., 2014). However, the most notable was the birth of calf of cloned embryos produced using somatic cells isolated from over decade old frozen–thawed semen (Selokar et al., 2014). This report has paved a new path for the restoration of highly precious progeny-tested bulls, which may have died many years before, but the semen of which is available in semen banks. It has also provided a possible approach for restoration of endangered species if their DNA is available in biological resource banks.
Birth of a cloned calf produced from urine-derived somatic cells, which is the first report across species (Madheshiya et al., 2015), was another important breakthrough in the field of buffalo cloning. It has demonstrated the feasibility of using biological fluids, which are easily obtainable in a noninvasive manner, as a source of somatic cells for somatic cell nuclear transfer (SCNT). In this article, we have discussed the developments and challenges of buffalo cloning and the possible roadmap for its future applications.
Methods of Buffalo Cloning
The method of producing cloned embryos appears to be quite simple and straightforward. In principle, the donor genome is fused with the cytoplasm of an enucleated ooplasm followed by activation of the recombined ooplasm to stimulate embryonic development to the blastocyst stage at which the embryos can be either transferred to suitably synchronized recipients or cryopreserved for future use (Wilmut et al., 1997). Two different versions of SCNT have been used for the production of cloned embryos in buffalo (Shah et al., 2008; Shi et al., 2007). The first is the traditional micromanipulation-based method of SCNT (Shi et al., 2007), which was used for the production of the first cloned mammal “Dolly”. This method is well established and has been extensively used in most cloning laboratories. Till date, 23 animal species, including buffalo, have been successfully cloned using this method (reviewed by Keefer 2015).
An alternative method, called Handmade cloning (HMC), which was developed as a simplified alternative to micromanipulation-based SCNT (Vajta et al., 2001), has been successfully used for production of cloned offspring in many farm animal species such as cattle, pig, sheep, goat, and buffalo (Du et al., 2007; Singla et al., 2015; Vajta et al., 2005; Zhang et al., 2013). A major advantage of HMC over the micromanipulation-based method is that there is no requirement for micromanipulators and its tools making instruments for enucleation and fusion and of highly skilled workforce to operate those instruments (Vajta 2007). This significantly reduces the cost of establishing a cloning laboratory due to which such laboratories can be set up in developing countries where availability of funds and technical expertise are major constraints.
In addition to multiple advantages of HMC, one practical problem is that two oocytes are used for the reconstruction of a single embryo, which increases the requirements of oocytes to generate cloned embryos, and use of two recipient oocytes to generate single cloned embryo may become responsible for higher mitochondrial heteroplasmy (Vajta 2007). We adopted this simplified method of SCNT developed by Vajta et al. (2001) and incorporated several modifications in the basic procedures resulting in efficient enucleation, fusion, and activation, leading to high blastocyst development rate (Selokar et al., 2012a; Shah et al., 2008).
Until now, more than 10 live healthy buffalo clones have been produced using this method by us (Singla et al., 2015), whereas only two healthy clones were reported using micromanipulation-based SCNT (Shi et al., 2007; Tasripoo et al., 2014). The technical differences between micromanipulation-based SCNT and HMC for the production of cloned embryos are illustrated in Figure 1. Published reports suggest that with higher blastocyst production rate, HMC can be used as an alternative method of SCNT.
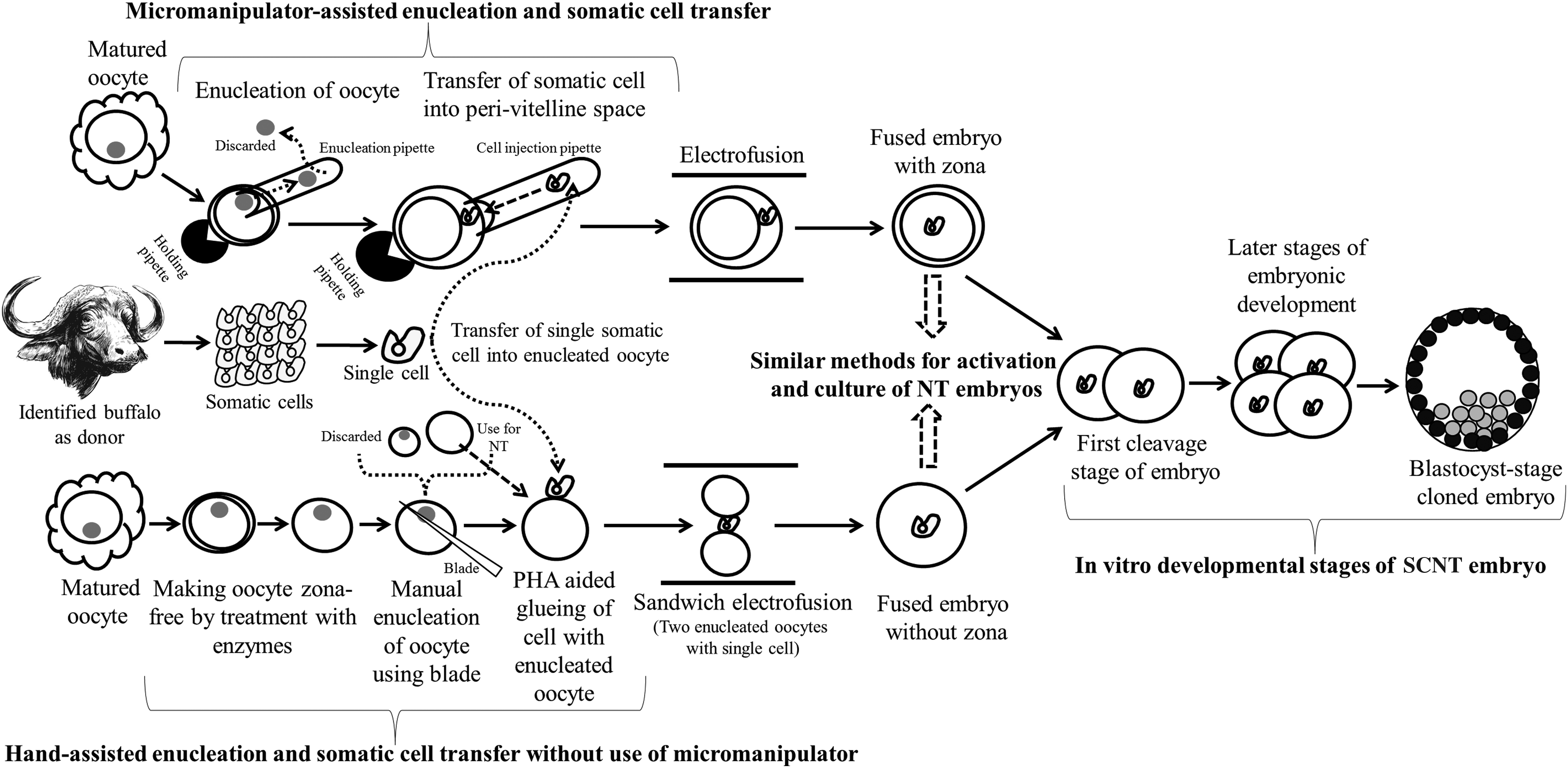
Schematic presentation of methods used for producing cloned embryos. In principle, a donor nucleus is transferred to one or two enucleated oocytes to generate a cloned embryo. The upper part, which illustrates micromanipulation-assisted traditional cloning, and the lower part, which describes micromanipulation-free Handmade cloning, have been used for cloning in buffalo.
History of Buffalo Cloning
The first live birth of cloned buffalo was reported in 2007 by Shi et al. (2007) in Chinese Swamp buffalo by micromanipulation-based SCNT using fetal fibroblasts and granulosa cells as the source of donor nuclei. In this study, three live calves were born, two out of which died at a very early age and one survived. This study has opened a new avenue for application of SCNT as a tool for making multiple copies of elite buffaloes at a fast pace for overall genetic improvement. The next breakthrough in buffalo cloning was made by us at National Dairy Research Institute, India in 2009 where we successfully produced a live calf of Murrah, a Riverine breed of buffalo by producing embryos by HMC using fetal fibroblasts as donor cells (Shah et al., 2009).
Following this, a number of cloned calves were produced using different types of donor cells of Murrah buffalo (Singla et al., 2015). In 2014, Tasripoo et al. (2014) reported the birth of a cloned calf in Thailand, using ear skin fibroblasts of an adult Swamp buffalo as donor cells. Recently, Central Institute for Research on Buffaloes, India has also produced a cloned calf of a semen-donating bull of Murrah (www.icar.org.in/en/node/10066). The timeline and the current status of buffalo cloning have been presented in Figures 2 and 3, respectively. Several buffalo clones were produced over the period of 10 years although with very low efficiency (Table 1). Therefore, more attempts and further refinements are needed with an aim to improve the overall efficiency of cloning.
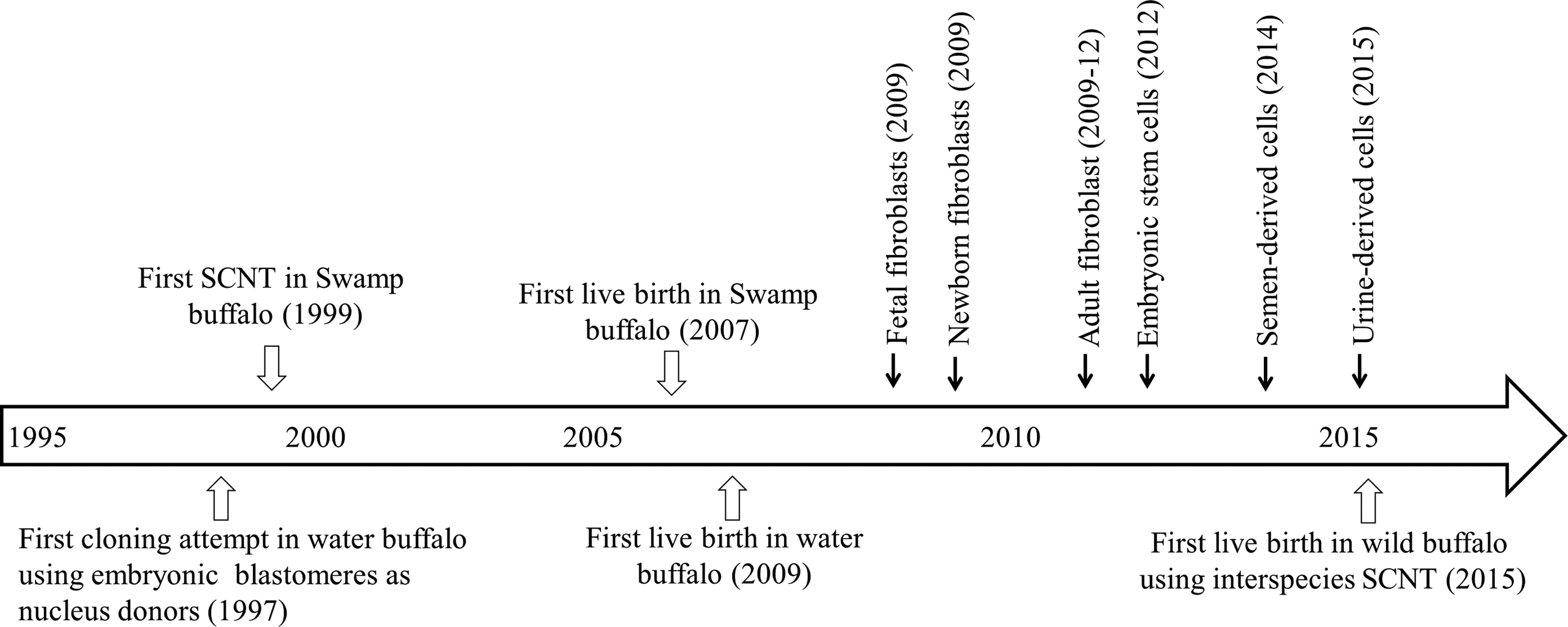
Major milestones in development of cloning technology in buffalo.

Comparison between micromanipulation-assisted TC and HMC.
More births were reported by Indian Council of Agriculture Research, (1) male cloned calf born from somatic cells isolated from cryopreserved semen of a progeny-tested bull (www.icar.org.in/en/node/7889 accessed on 23 March 2017), (2) wild female buffalo (www.icar.org.in/en/node/8465 accessed on 23 March 2017), and (3) male cloned calf born from somatic cells isolated from tail-derived cells of a superior bull (www.icar.org.in/en/node/10066 accessed on 23 March 2017). All these calves are surviving and growing normally (our personal observation), and these calves were not included in above table as they are not reported in any research publication.
Improving the Developmental Competence of Cloned Embryos
Several approaches have been used to improve the cloning efficiency in buffalo. These include (1) use of somatic cells from diverse sources as donor cells (Jyotsana et al., 2016; Madheshiya et al., 2015; Mohapatra et al., 2015a; Parnpai et al., 1999; Selokar et al., 2014; Shah et al., 2009; Shi et al., 2007), (2) use of recipient oocytes of good quality (Lu et al., 2011; Mohapatra et al., 2015b; Panda et al., 2011), (3) improvements in culture conditions (Saini et al., 2015; Shah et al., 2008), (4) proper synchrony of the cell cycle stage of donor cells (Saikhun et al., 2004; Selokar et al., 2012b; Shi et al., 2007), (5) optimization of conditions for electrofusion of somatic cell with oocyte (Selokar et al., 2012a), and (6) use of epigenetic drugs to modify the epigenetic marks (Luo et al., 2013; Panda et al., 2012; Saini et al., 2016, 2017; Srirattana et al., 2014; Sun et al., 2015).
These attempts have resulted in increase in blastocyst production rates to levels comparable to those obtained in other species; however, a number of clones produced were much lower in comparison to other farm animal species. Possible reasons for less cloned animal births in buffalo may be as follows: (1) the focus of most of studies has been on in vitro rather than in vivo developmental competence of cloned embryos, (2) a very few laboratories are engaged in buffalo cloning, (3) lack of strategic planning and investment in countries where buffalo is a prime livestock, and (4) intrinsic reproduction problems of buffalo may be responsible for lower conception rate. On basis of our previous experiments, the factors which influence the in vitro development of HMC cloned embryos in buffalo are presented in Figure 4. In the following section, we will discuss the approaches used to improve the developmental competence of cloned buffalo embryos in detail.

Critical factors which affect the in vitro development of cloned embryos produced by HMC.
Recipient oocyte
The oocyte is a unique and highly complex cell, which reprograms the donor nucleus during SCNT, and is one of the major factors that affect the outcome of cloning. In vitro maturation of oocytes is a crucial step for enabling them to become capable of reprogramming the donor cell for converting it from differentiated to totipotent state. Following the report of Campbell et al. (1993) in which MII phase was observed to be the most appropriate phase of oocytes for the production of SCNT embryos from differentiated cells in mammals, MII oocytes have been used in almost all cloning experiments across species.
In buffalo, only a few investigations have been carried out to explore the role of the oocyte in influencing the developmental competence of SCNT embryos. Lu et al. (2011) reported that when oocytes matured in vitro for 28, 29, 30, 31, or 32 hours were activated, more oocytes matured in vitro for 30 hours developed into blastocysts and that activated at 3 hours after electrofusion gave higher blastocyst rate than activation at 1 hour suggesting that the recipient oocyte age and the interval from fusion to activation affect the developmental competence of buffalo SCNT embryos.
The loss of 20%–50% cytoplasmic volume during enucleation by protrusion cone-guided bisection of oocytes is a major problem in HMC, which compromises the developmental competence of embryos (Vajta et al., 2001). This can be compensated by use of two demi-cytoplasts instead of one for production of reconstructed embryos. Panda et al. (2011) reported that instead of using two demi-cytoplasts for generation of reconstructed embryos, increasing the cytoplasmic volume further by use of three demi-cytoplasts was not beneficial, while reducing the cytoplasmic volume by use of a single demi-cytoplast seriously compromised the developmental competence.
In addition to these findings, Brilliant Cresyl Blue (BCB) staining can be used to select competent oocytes for SCNT. We have found that BCB+ oocytes supported higher in vitro developmental competence, and the resulting embryos were of better quality than those produced from BCB- oocytes (Mohapatra et al., 2015b). A few investigators have examined the ability of buffalo oocytes to reprogram somatic cells of other animals (Lu et al., 2005; Selokar et al., 2011). However, the developmental competence of interspecies embryos was found to be poor in these investigations; therefore, buffalo oocytes would not be an ideal choice for interspecies SCNT.
Donor cell type
Ideally, all the cells of an individual possess an identical genome, and therefore, any cell of the individual can be reprogrammed following SCNT. In the very first report in buffalo, embryonic blastomeres were used as nuclear donors to produce cloned embryos by micromanipulation-based cloning; however, the reconstructed embryos failed to achieve development to the blastocyst stage (Singla et al., 1997). Later, fetal fibroblasts were successfully reprogrammed, and blastocyst stage cloned embryos were produced (Parnpai et al., 1999).
Many somatic cell types, including embryonic stem cells (George et al., 2011; Panda et al., 2011), fibroblast cells from fetal, newborn, and adult animal (Saikhun et al., 2004; Shah et al., 2009; Srirattana et al., 2010), mammary gland cells (Golla et al., 2012; Selokar et al., 2016), cumulus and granulosa cells (Pandey et al., 2010; Parnpai et al., 2001; Shah et al., 2008; Shi et al., 2007; Srirattana et al., 2010), blood cells (Jyotsana et al., 2016), trophectoderm cells (Mohapatra et al., 2015a), semen-derived somatic cells (Selokar et al., 2014), and urine-derived somatic cells (Madheshiya et al., 2015), have been successfully used as nucleus donors in buffalo and yielded blastocyst production rate ranging from 10% to 20% in micromanipulation-based cloning and 35%–45% in HMC. The efficiency of various cell types is presented in Table 1. However, it is still unclear which cell type is the most efficient nuclear donor for buffalo cloning and needs further investigations.
Cell cycle synchronization
It has been demonstrated in bovine SCNT that synchronization of donor cells in the G0/G1 stage of cell cycle offers better survival of cloned embryos after transfer to surrogate mothers (Gibbons et al., 2002); however, cloned animals have also been produced from donor cells in different stages of cell cycle (Bordignon and Smith, 2006; Tani et al., 2001). Therefore, it is still debatable which cell cycle stage may result in the best cloning efficiency across species.
In buffalo, various cell cycle synchronization methods such as total confluence, serum starvation, or chemical treatments have been used to synchronize cells in the G0/G1 stage of the cell cycle (Saikhun et al., 2004; Selokar et al., 2012b; Shi et al., 2007). These investigations suggest that cell synchronized at G0/G1 stage of cell cycle resulted in high blastocyst production rates, and at present, donor cells are cultured to total confluence or serum starved before being used for SCNT (Saini et al., 2017; Tasripoo et al., 2014).
Electrofusion of somatic cell and oocyte
Efficient electrofusion is an important factor, which contributes to efficient generation of cloned embryos. Electrofusion of donor cell and enucleated oocyte is used extensively because of lower toxicity, good repeatability, and higher efficiency than of other methods of fusion such as use of Sendai virus (Zimmermann and Zimmermann, 1996). We have earlier examined different electrofusion parameters such as AC alignment voltage, DC pulses, fusion methods, and orientation and position of somatic cell demi-cytoplasts during electrofusion for production of reconstructed buffalo cloned embryos for HMC (Selokar et al., 2012a). We found that the optimum parameters for production of buffalo embryos by HMC include the use of a single step fusion method, in which alignment and fusion are carried out by subjecting the enucleated oocyte demi-cytoplasts to 4 V AC followed by a single pulse of 3.36 kV/cm DC for 4 μsec.
These conditions have been used for production of cloned embryos by HMC in all the studies at our laboratory. In micromanipulation-based SCNT, the first polar body with metaphase-II plate is removed using specialized aspiration pipette followed by transfer of a single cell into the perivitelline space of the enucleated recipient oocyte with a micropipette under an inverted microscope equipped with a micromanipulator, followed by electrofusion using different pulses and conditions for fusion of transferred somatic cell with recipient oocyte (Parnpai et al., 2001; Saikhun et al., 2004; Shi et al., 2007).
Culture medium and culture system
The system and media used for in vitro culture of cloned embryos are major factors affecting their in vitro development. Several different types of culture media and systems have been used for the in vitro culture of cloned embryos, and most of them have supported development of embryos to varying degrees. Zona-enclosed cloned embryos derived from micromanipulation-based cloning are generally cultured in 30–100 μL droplets having feeder layer of adherent cells (Kitiyanant et al., 2001; Parnpai et al., 1999; Shi et al., 2007), whereas embryos produced by HMC showed compromised developmental competence in this culture system (Shah et al., 2008; Vajta et al., 2003, 2005). The zona-free embryos produced in HMC require attentions such as culture of individual embryos and minimal shaking during culture to prevent aggregation of embryos. Vajta et al. (2000) developed a special culture system called well-of-the-wells (WOW) for the culture of zona-free embryos.
This system has given good results in many species and is currently extensively used for the culture of embryos produced by HMC. Conversely, Shah et al. (2008) reported that a flat surface culture system, in which 15–20 zona-free embryos are cultured in 400 μL of medium in four-well dish, was more efficient than WOW for in vitro development of buffalo embryos produced by HMC. This discrepancy may be due to species specificity and differences in culture media and conditions used among various studies. Later, we found that micro-droplet culture system was also an efficient system and resulted in blastocyst rate ranging from 30% to 40%, which is comparable to flat surface (Kaith et al., 2015).
In this system, 4–6 microdrops of 20 μL each were made in the well corners of four-well dish and overlaid with 300–400 μL sterile mineral oil. Individual zona-free embryos were placed in each droplet and handled carefully to avoid shaking off embryos or merging of droplets (Kaith et al., 2015). However, further investigations are needed to improve the culture systems for more efficient production of cloned buffalo embryos.
Culture medium not only provides nutrients for embryonic development in vitro but also influences in vivo development. Requirement of medium depends upon stage of development of embryo and type of embryo (IVF or SCNT) and species. In case of buffalo SCNT embryos, TCM-199 or SOF-based medium with microdrop culture surface has been extensively used in micromanipulation-based cloning (Kitiyanant et al., 2001; Parnpai et al., 1999; Shi et al., 2007), whereas RVCL medium with flat surface system has been found to be the best for culture of embryos produced by HMC (George et al., 2011; Saini et al., 2017; Selokar et al., 2012a; Shah et al., 2008). The efficacy of several other commercially available media needs to be examined to further improve the cloning efficiency.
Epigenetic reprogramming
Reprogramming refers to the process through which the differentiated donor cell is converted to the totipotent state (Eckardt and McLaughin, 2004). It is generally believed that low cloning efficiency is due to incomplete or aberrant nuclear reprogramming of the donor cell by the recipient oocyte. Reprogramming of the genome is a highly complex process involving timely activation and deactivation of a large number of genes leading to proper expression of proteins. Previous investigations in both Swamp and Riverine buffaloes suggested that cloned embryos are hyper DNA methylated, and histone acetylation, methylation status, and gene expression pattern in the cloned embryo are different from that of their IVF counterparts (Jyotsana et al., 2016; Luo et al., 2013; Mohapatra et al., 2015b; Saini et al., 2016, 2017; Srirattana et al., 2014; Sun et al., 2015; Suteevun et al., 2006a, 2006b).
To achieve proper reprogramming, the epigenetic status of cloned embryo needs to be similar to that of IVF embryos. Suteevun et al. (2006a) reported that the relative levels of DNA methylation/nuclei of swamp cloned embryos were decreased from the 2-cell stage to 8-cell stage and then began to increase at the morula stage and reached the highest levels at the blastocyst stage, whereas the relative levels of global DNA methylation increased from 2-cell stage to blastocyst stage.
In addition, they observed the lowest levels of histone acetylation level at the morula stage among all preimplantation stages. In case of riverine HMC embryos, there are no data on epigenetic changes during developmental stages (2-cell stage to blastocyst); however, analysis at blastocyst stage indicated that HMC blastocysts are epigenetically different compared with IVF counterpart (Jyotsna et al., 2016; Mohapatra et al., 2015b; Saini et al., 2017). Abnormal epigenetic status of DNA and histone may contribute to the developmental failure of buffalo SCNT embryos. Therefore, different approaches have been used to correct the reprogramming process by modifying the epigenetic status of somatic cells or embryos by treatments with epigenetic modifiers, suppression of epigenetic regulation genes, exposure to extracts of oocytes, and combinations of above approaches.
Although the application of these treatments has led to considerable improvements in blastocyst rate and quality (Sadeesh et al., 2017; Saini et al., 2016, 2017; Srirattana et al., 2014; Sun et al., 2015), its applicability to increase the live birth rate is yet to be established due to nonavailability of sufficient data on the in vivo developmental competence of these cloned embryos. The effects of epigenetic agents which modify the epigenetic status on global transcriptome or proteomic and on gene-specific methylation reprogramming in cloned embryos need to be studied.
Perspectives and Prospective
Both micromanipulation-based SCNT and HMC can be used for the production of cloned buffaloes and have many applications such as multiplication and conservation of elite germplasm, restoration of proven bulls even after death, conservation of wild ungulates, and production of transgenic animals. Despite extensive research leading to optimization of cloning procedures and several breakthroughs, the most important factor limiting the application of this technology on a wide scale is the abysmally low live birth rate.
Application of several approaches that were used to improve the epigenetic status of cloned embryos led to significant improvements in blastocyst rate and quality. However, to what extent they influence the live birth rate is still unknown due to nonavailability of sufficient data. The focus of future research work needs to be on determining the conception and live birth rate following transfer of cloned embryos to recipients in large numbers. In addition, research efforts should be aimed at understanding the reprogramming process to identify the abnormalities introduced in cloned embryos due to aberrant reprogramming. In parallel, more support is needed from governments or private funders in Asian countries to promote buffalo cloning for its utilization to mitigate the ever growing demand for food (milk or meat) or drugs (through transgenesis).
The major challenges of buffalo cloning in developing countries, where buffalo dominates, are as follows: (1) preference of cattle/pig over buffalo milk and meat due to traditional practices, (2) high cost of technological inputs, (3) nonavailability of skilled scientists, technicians, and field workers in sufficient numbers, and (4) absence of coordination among research scientists, animal husbandry departments, and industries for technology transfer. By addressing these issues, it could hasten the improvement and application of this technology especially in emerging countries like India and China.
Footnotes
Acknowledgments
Buffalo cloning work in India was supported by Indian Council of Agricultural Research, Delhi, and Department of Biotechnology, Delhi, and Chhattisgarh Wildlife Department, Raipur, and Science and Engineering Research Board, Department of Science and Technology, Delhi. Authors highly acknowledged continuous supports from members of cloning teams working in two Indian institutions, ICAR-National Dairy Research Institute, Karnal and ICAR-Central Institute for Research on Buffaloes, Hisar.
Author Disclosure Statement
The authors declare there are no conflicts of interest.