
Abstract
Abstract
The first 20 years of somatic cell nuclear transfer can hardly be described as a success story. Controversially, many factors leading to the fiasco are not intrinsic features of the technique itself. Misunderstandings and baseless accusations alongside with unsupported fears and administrative barriers hampered cloners to overcome the initial challenging period with obvious difficulties that are common features of a radically new approach. In spite of some promising results of mostly sporadic and small-scale experiments, the future of cloning is still uncertain. On the other hand, a reincarnation, just like the idea of electric cars, may result in many benefits in various areas of science and economy. One can only hope that—in contrast to electric cars—the ongoing paralyzed phase will not last for 100 years, and breakthroughs achieved in some promising areas will provide enough evidence to intensify research and large-scale application of cloning in the next decade.
Introduction
T
The general reaction was negative—and remained more or less negative up to this day. Sharply separated good and bad science by putting cloning in the latter category; some professionals agreed with this opinion or supposed fraud; grants for cloning attempts were ceased and serious legal restrictions were implemented, including decades in prison for any attempt to make human clones.
In that tensed atmosphere only a handful of scientists realized the enormous value of the discovery: the fact that somatic cell nuclear transfer (i.e., cloning by replacing the nucleus of a mature egg with a nucleus of a cell from an adult mammal) destroyed the principal dogma of developmental biology. The reversibility of “terminal” differentiation was a revelation comparable with dethroning the Earth as the center of the universe or the theory of general relativity. Dozens of exciting questions arose immediately and hundreds of different cloning experiments could be started to answer them. Instead, most leading senior experts—after some hesitation—just shrugged: “well, if it is possible, it is possible”, and put the whole issue ad acta.
The other aspect of the discovery has been left unexploited, as well. Cloning offered great perspectives for practical purposes, including biomedical and agricultural applications. Genetic modification of somatic cells before nuclear transfer opened new ways for creation of transgenic animals with various features, including universal donors for organ xenotransplantation, models for human diseases with genetic background, and living bioreactors to manufacture inexpensive organic compounds for diagnostics and therapy. However, even without genetic modification, cloning itself provided great possibilities in agriculture to multiply rapidly genetically valuable animals.
None of these purposes has been appropriately realized during the past 20 years. The lack of support of basic research has become anecdotic: “if you want to win a grant, do not mention cloning.” Only a few research groups received support, mostly for attractive but rather difficult, sometimes utopistic purposes, including saving endangered or extinct species. Some brave venture capitals in agriculture or biomedicine were fascinated by the perspective, but the lack of immediate success disappointed them and the facilities were closed down in 2–3 years. Legal restrictions hampered the use of cloned animals in large markets, including the whole European Union (http://sciencemag.org/news/2015/09/eu-parliament-votes-ban-cloning-farm-animals). In other countries intellectual property issues were used to destroy competitor commercial ventures.
Apart from the external factors, however, cloners had some negative contribution in the lack of development, as well. We have accepted—and this way recreated—our limits, have conducted cloning experiments under compromised conditions, including instruments, logistics, and financial support, with the obvious inferior outcome. We failed to listen to each other; isolated research groups were dealing with problems that had been resolved by others years, sometime a decade earlier. We lacked the creativity; followed the original schemes without the open-mindedness and courage of the great pioneers of the end of the last century. We failed to convince the world about the importance of our research, as did our fellow scientists in genomics and stem cell research.
Finally, we gave up too early. The number of publications dealing with cloning decreased dramatically (Fig. 1). Many of us (including me) have left the field either temporarily or terminally. In Australia, at least seven cloning laboratories with considerable international reputation flourished in the late 90's—now there are two small commercial ventures, and around 30–40 cloned cattle in the whole country (Food Standards Australia New Zealand, March 2016; http://foodstandards.gov.au/consumer/foodtech/clone/Pages/default.aspx), out of the 25 million head (Fast Facts 2016-Meat and Livestock Australia; https://mla.com.au/globalassets/mla-corporate/prices–markets/documents/trends–analysis/fast-facts–maps/mla_beef-fast-facts-2016.pdf).
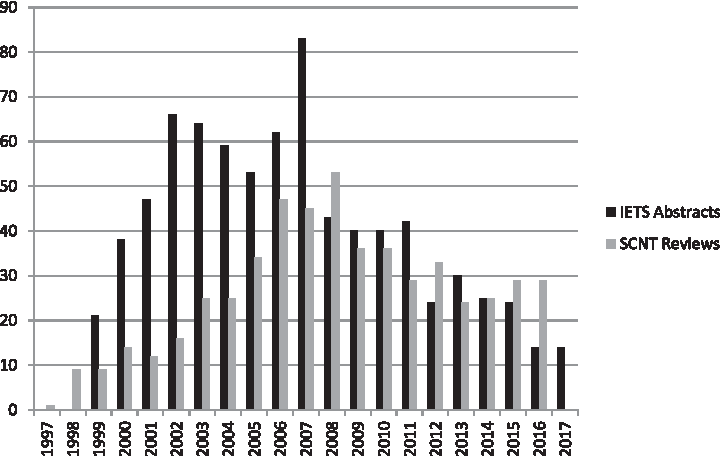
Number of publications/year dealing with somatic cell nuclear transfer. Black: Relevant abstracts on the annual meetings of the International Embryo Transfer Society (up to 2017). Grey: Reviews dealing with somatic cell nuclear transfer according to PubMed (up to 2016).
The number of (real) cloners around the world could be several hundreds, twenty years ago—now an optimistic estimation would say several dozens.
According to an anonymous expert in the field, “cloning is still a technology badly looking for home.”
Light at the End of the Tunnel
Although, in vitro developmental rates of cloned embryos have slowly but steadily increased during the past 15 years, the overall efficiency is still low. The bottleneck in scientific, and especially in commercial application is the healthy offspring per transferred embryo rate. Many scientists and most laymen regard it as an intrinsic feature of this “unnatural” intervention that cannot be changed.
We have to acknowledge that cloning stretches flexibility and adaptive ability of oocytes and the transferred genomic material to their limit. Reprogramming of the nucleus, which is, to remove all blocks from the chromosomes of a somatic cell and to make it suitable to govern development from the very beginning–and to accomplish this process in a few hours—is a task unparallel with any known processes in mammalian developmental biology. However, the fact that certain reconstructed embryos can resolve it demonstrates clearly that it is difficult, but not impossible.
Accordingly, our goal should be to provide maximum help in this process. Theoretically, there are at least four feasible ways for this support.
Finding the proper cell cycle phase of oocytes and somatic cells for optimal support of reprogramming
The success of cloning may depend on the proper synchronization of cell cycle phases of the recipient oocyte and donor somatic cell, see review by Shufaro and Reubinoff (2011). For metaphase II (M-II) oocytes, donor cells in the quiescent G0/G1 phase were suggested. Initially, it has been achieved by serum starvation (Wilmut et al., 1997), later it was replaced by growing monolayers to confluency alone (Hayes et al., 2005) or in combination with serum starvation (Ma et al., 2015). Efficient synchronization was also achieved with the kinase inhibitor roscovitine (Gibbons et al., 2002; Park et al., 2010; Selokar et al., 2012).
A comparative investigation did not find any difference regarding the in vitro development of cloned embryos reconstructed with fibroblasts after serum starvation, contact inhibition, or roscovitine treatment (Sun et al., 2008). It should also be noted that the need of synchronization of somatic cells has also been questioned, and some reports found no difference in outcomes using serum-starved or actively growing donor cells (Cibelli et al., 1998; Peura et al., 2003). In everyday practice of farm animal cloning, the most common method is to produce confluent monolayers from skin fibroblasts without additional treatments (Callesen et al., 2014; Lee and Maalouf, 2014; Liu et al., 2015).
An alternative approach for cell cycle synchronization, using telophase II (T-II) recipient cytoplasts and S/G(2) phase fibroblasts obtained with roscovitine synchronization was reported equally efficient regarding both in vivo and in vitro development (Bordignon and Smith, 2006) and is now used by several companies for commercial cattle cloning (M. Maserati, pers. comm.).
Another significant modification of the traditional sequence of events in cloning is the reverse-order nuclear transfer (Peura, 2003), where fusion and activation are performed first, followed by the removal of the chromatin of the recipient oocyte hours later. The seemingly complicated task of delayed enucleation is surprisingly easy when the zona-free approach is used (see discussed later). In sheep, reverse-order nuclear transfer improved embryo development in vitro. Unfortunately no attempt was made to observe its effect on in vivo development, so far.
Nonspecific modification of the donor genome to increase accessibility for reprogramming
Chemical agents as DNA methyltransferase or histone acetylation inhibitors, such as 5-aza-2′-deoxycytidine or trichostatin A, Scriptaid, and oxamflatin, were reported to be efficient to promote development in vitro, in some studies also in vivo (Kishigami and Wakayama, 2009; Kwon et al., 2017; Mao et al., 2015; Saini et al., 2017; Sawai et al., 2012; Wang et al., 2011a, 2011b). Surprisingly, some of these agents may have a selective pathway modifying primarily the function of developmentally important genes. Although, the mechanism is still not entirely clear even for the most popular agent, trichostatin A (Hosseini et al., 2016), recently it has been revealed that it may be more selective than supposed earlier and specific to developmentally important genes (Inoue et al., 2015).
Treatment with biological agents, including cell-free extracts of T cells or oocytes was also found (Landsverk et al., 2002; Liu et al., 2012; Rathbone et al., 2010; Yang et al., 2012). Unfortunately, convincing evidence about the increase of overall efficiency and elimination of developmental abnormalities is only available in mice, accordingly none of these treatments is routinely applied in other fields, for example domestic animal cloning. Extensive research would be needed to find the right agent or combination of agents, as well as the right dose and timing of treatment. Considering also the need of a large number of embryo transfer experiments, and years to evaluate the outcome, this approach is promising but does not offer a rapid solution.
A new direction is to induce a complex change in the somatic cell genome before transfer, to generate a sperm-like stage with protaminization of somatic cell nuclei through the heterologous expression of the protamine 1 gene (pPRM1-RFP) or analog agents. This intervention may make the genome more accessible to reprogramming by mimicking the natural way that happens after fertilization (Iuso et al., 2015; Loi et al., 2016). If this elegant theory proves its efficiency and practicality, it may have considerable impact on the future of nuclear transfer, although, even the transitional manipulation of the genome function might create additional arguments against cloning.
Targeted interventions to modify the function of developmentally important genes
Initial attempts to detect major differences between clones and their naturally conceived counterparts have failed to produce information of practical importance, due to the huge amount of seemingly incoherent alterations (Wrenzycki et al., 2006). Recently, however, due to the rapid advancement in genomics and gene editing, together with extensive reprogramming studies in laboratory mice, attention has been focused on a few important mechanisms that may contribute to the inefficiency of nuclear transfer in other mammalian species, too.
One of these mechanisms is the overexpression of the Xist gene responsible for X chromosome inactivation, causing downregulation of some developmentally important genes (Matoba et al., 2011), although the knockdown strategy only works in males (Oikawa et al., 2013). Another important alteration is related to histone H3 lysine 9 trimethylation (H3K9me3) that may hamper the reprogramming process (Inoue et al., 2010). However, there is still a massive dysregulation of gene expression with only partially known mechanism involving genes responsible for placental structure and signaling between the conceptus and endometrium (Biase et al., 2016). While targeted technologies are available to correct the first two mechanisms, placenta-related problems may require a more general approach.
To help the oocyte and the reconstructed embryo itself to make the job properly
Obviously this attempt has been used since the very beginning of cloning research, however, the resources are still far from exploited. Required improvements include:
(i) A highly efficient and reliable (background) in vitro embryo production system based on slaughterhouse-derived, in vitro-matured, and parthenogenetically activated oocytes, The use of parthenogenetic activation excludes inconsistencies related to sperm and the fertilization step, and allows monitoring the efficiency of the activation procedure. Culture to the latest possible phase of preimplantation development is preferred, to ensure that the most competent embryos are transferred. In vitro embryo production systems differ between species in requirements and achievable outcome. It is of utmost importance to select and run consistently the very best one for a given species for sensitive and intrinsically handicapped cloning embryos. For example in cattle, a consistent 50% D7 blastocyst per oocyte rate should be regarded as the minimum requirement, preferably over 90% of these blastocysts hatching in the subsequent day. This efficiency was published long ago (Holm et al., 1999; Vajta et al., 2003); still, many bovine embryo laboratories consider 35%–40% blastocyst and 60%–70% hatching rates appropriate, disregarding the fact that in such a system the quality of all embryos will be more or less suboptimal. (ii) Methods used for somatic cell isolation should be carefully selected. Although, rarely published, seemingly negligible differences in obtaining samples, time between sample collection and processing, conditions of transportation, methods used for cell separation, even the applied antibiotic–antimycotic solution may have considerable effect on the outcome. Monolayers with identical morphology may dramatically differ in the number of developmental abnormalities of clones that are often detected only in the postpartum phase. Even if maintenance, passage, and cryopreservation of somatic cell cultures seem to be a simple task, it is a good idea to get it made by an expert in cell cultures. Embryologists have different standards and approaches, and in somatic cell cloning their attention is focused on oocytes and embryos. They may disregard the special needs of the seemingly tolerant somatic cell cultures. This approach may lead to serious consequences, although contributing factors remain mostly undetected. (iii) All work related to oocytes and embryos should be thoroughly learned and practiced, simplified, and shortened. A recent publication demonstrated clearly the importance of practice: the same bovine somatic cell nuclear transfer procedure performed by the same personnel in the same laboratory has resulted in continuous and dramatic improvement through three subsequent periods (7 months each, separated from each other by3-month intervals). Apart from the 2.4-fold increase in blastocyst per reconstructed embryo rates between the first and third period, pregnancy per embryo transfer rates achieved with cloned blastocysts also have shown a stunning six-fold increase (6.7%–40%) (Gerger et al., 2017). For oocytes and embryos, the optimal in vitro environment is the dark warm incubator with maximum humidity and optimal gas atmosphere. Extensive exposition to the inconsistent and suboptimal environment of the laboratory room is harmful. Every minute, every spared small trauma counts. The anecdotic statement that “the good cloner is the fast cloner” is true, but speed and experience alone may not be enough. In this regard, radical changes of the traditional cloning technology have already proved its value, but the possibilities are still far from fully exploited (see discussed later). (iv) To provide cloned embryos a very special care after they leave the laboratory, that is, during transportation to recipients, and embryo transfer to the uterus. Measures appropriate for in vivo-derived or in vitro-fertilized embryos may be insufficient for fragile cloned ones. Optimally, recipients should be located just on the other side of the wall of the laboratory. If this is not possible, the simple loading embryos in sealed straws and shipping in purpose-designed incubators may be insufficient, leading to compromised outcomes. Although, technically demanding, the optimal method for transportation is to use the original dishes, original media, and absolutely consistent environment (gas atmosphere, temperature), with minimal mechanical trauma. The ultimate solution to minimize logistic problems will be the storage and transport of cryopreserved cloned embryos (see discussed later).
Transcervical, surgical, or laparoscopic embryo transfer and recipient management (before and after embryo transfer) should also be done at the highest level by a specialist with extensive experience in all aspects of the technique.
Alternative Approaches and Complementing Techniques
Cloning without the zona pellucida
A profound modification of the traditional, micromanipulation-based technology, zona-free cloning (Peura et al., 1998) with its subsequent version adapted to somatic cell nuclear transfer called handmade cloning (HMC) (Vajta et al., 2001, 2003). The principle of HMC is that the zona pellucida is not needed for in vitro development of preimplantation stage mammalian embryos, and its removal with enzymatic digestion does not make irreversible damage of M-II oocytes. Oriented, chemically assisted enucleation can be performed manually by a microblade or a micropipette under a stereomicroscope (Hosseini et al., 2015; Vajta et al., 2005); in skilled hands the procedure is quicker and more accurate than with the micromanipulator-based technique.
Attachment of somatic cells to cytoplasts is facilitated with phytohemagglutinin, and the subsequent fusion is more efficient without the zona pellucida (Vajta et al., 2003). The technique and the efficiency of activation is essentially the same as for traditional cloning. As precompaction-stage zona-free embryos are absorbed in the uterus (Vajta et al., 2010a), extended embryo culture to the late morula–blastocyst stage is required. During this period, the well of the well (WOW) system avoids individual blastomeres to fall apart, and also prevents aggregation of multiple embryos (Vajta et al., 2000).
HMC is often regarded as a cheap version of somatic cell nuclear transfer. This statement is correct, in that the investment required to enable a basic animal embryo laboratory to start cloning is with an order of magnitude lower by selecting HMC.
However, other differences may be even more important, including the rapid and less demanding work; equal or higher efficiencies in each step, including enucleation, fusion, activation, and in vitro development (Vajta et al., 2003). For the latter, it has been discovered that the WOW system offers additional benefits promoting embryo development possibly by concentrating autocrine and paracrine factors and stabilizing the microenvironment required for optimal development (Akagi et al., 2010; O'Neill, 2008; Sugimura et al., 2012; Vajta et al., 2010b). Both theoretical calculations and biological evidences prove that even the shape of the well modifies this effect (Feltrin et al., 2015; Matsuura, 2014).
Another benefit of the zona-free situation is the possibility to fuse the somatic cell with two enucleated oocytes (Tecirlioglu et al., 2003). The sandwich arrangement increases the fusion efficiency to 95%–100%, and improves embryo development by facilitating reprogramming and/or providing larger cytoplasmic volume to elevate the cell numbers in blastocysts that is usually low after somatic cell nuclear transfer (Akagi et al., 2014; Liu et al., 2017; Ribeiro et al., 2009). Concerns regarding heteroplasmy caused by three different possible sources of mitochondria were not justified in recent large-scale experiments resulting in the birth of hundreds of healthy offspring (Callesen et al., 2014; Liu et al., 2015).
The lack of the zona pellucida did not compromise results after cryopreservation (see discussed later) or embryo transfer. HMC has resulted in offspring in cattle, sheep, and pigs (Du et al., 2007a; Peura and Vajta, 2003; Tecirlioglu et al., 2003; Zhang et al., 2013), in water buffalo (Saha et al., 2013), and also in goats (Nasr-Esfahani et al., 2011). Foals were produced with a hybrid version using micromanipulators for enucleation followed by the zona-free approach for fusion and embryo culture (Galli et al., 2008; Lagutina et al., 2007).
A representative work in pigs based on more than 200,000 reconstructed embryos has reported around 75% pregnancy rates after laparoscopic transfers, and ∼75% of pregnancies led to birth of healthy piglets (Liu et al., 2015). Another group, using the same approach reported that transfer of more than 5000 porcine cloned embryos resulted in around 90% pregnancy rates, and 90% of them ended up with healthy piglets. The number of piglets per embryos transferred was an impressing 15% (Callesen et al., 2014). No comparable efficiency with traditional cloning in any species has been achieved so far.
Unfortunately, in spite of repeated efforts, no large-scale trials could be performed in cattle; however, pilot experiments (even if performed under compromised conditions) prove that 25%–33% healthy offspring per transfer rates are achievable (Cortez et al., 2018; Vajta et al., 2004).
Arguments against HMC include the concern that the zona-free situation makes embryos fragile. The everyday practice does not justify it, all tools and manipulations routinely used for zona-intact embryos may be used for zona-free ones, too. The only mechanically sensitive period is the precompaction phase, when embryos should be left undisturbed in the protective WOWs. Disease transmission issues are also raised, disregarding that all traditionally cloned (or biopsied) embryos miss the defense provided by an intact zona pellucida.
The most significant reason hampering the wide acceptance of HMC is probably a paradox one: it is too simple to be taken seriously. It does not require expensive equipment, and everything can be done with handheld tools. The situation is similar to that of vitrification. It was first applied for embryos more than three decades ago (Rall and Fahy, 1985), and completely fine-tuned between 1998 and 2004 for human use (Vajta and Nagy, 2006). Still, widespread application was delayed for almost a decade because embryologists failed to trust a method based on a primitive plastic tube or stick. This mental barrier hampered hundreds of thousands of infertile patients to get the most efficient available treatment, and deprived many of them to have a baby—irreversibly.
Stress for stress tolerance
If the problem with HMC is its simplicity, the widespread application of another technology with proven positive effect on cloning is hampered by the required sophisticated machine, and lack of courage to trust and test a seemingly drastic procedure. The principle of this approach is that carefully chosen and precisely controlled sublethal stress treatment may increase the adaptation ability and subsequent stress tolerance of gametes and embryos. As a consequence, improved survival rates can be achieved after mechanical, chemical, and thermal stress caused—among others—by somatic cell nuclear transfer, embryo culture, and cryopreservation with long-term positive effect on pre- and postimplantation development, births, and healthy offspring rates (Pribenszky and Vajta, 2011).
Various treatments, including osmotic stress (Lin et al., 2009), exposition to chemicals (Rahman et al., 2012; Vandaele et al., 2010), and heat shock (Isom et al., 2009) have resulted in improvements in the developmental competence in gametes and embryos of experimental and domestic species (Pribenszky et al., 2010). However, these stressors were difficult to control and could also result in adverse effects (Gharibzadeh et al., 2015; Hansen, 2007; Mullen et al., 2004).
On the other hand, high hydrostatic pressure (HHP) is controllable, it acts immediately and uniformly at each point of the sample, without penetration problems or gradient effects, and it can be applied with high accuracy, consistency, and reliability (Pribenszky and Vajta, 2011). In contrast to the extreme pressure (20–80 MPa, the latter equals with the water pressure at the deepest point of the Atlantic Ocean) it is well tolerated by the samples and does not mean any danger to the laboratory personnel. Accordingly, it is regarded as the optimal stressor, and has been used in many studies, with positive effects obtained in most of them.
Most importantly—from the point of somatic cell nuclear transfer—HHP treatment of in vitro matured porcine oocytes improved their stress tolerance (Pribenszky et al., 2008), development after vitrification (Du et al., 2008b), and somatic cell nuclear transfer (Du et al., 2008a). The latter effect may be explained by the altered RNA constitution and expression of imprinted genes during embryo development (Lin et al., 2014).
Unfortunately, no studies of the effect of HHP on somatic cell nuclear transfer in other species are available yet, due to factors mentioned in the first paragraph of this chapter. Right now, when cloning is balancing on the edge of survival, it is rather a luxury not to exploit the possibilities of this approach.
Cryopreservation
Practical application of assisted reproductive technologies in domestic animals—first of all cattle—has become feasible with the introduction of appropriate cryopreservation technologies. Widespread use of artificial insemination in cattle started after the discovery of an efficient sperm freezing technique (Polge, 1952), and the success of multiple ovulation/embryo transfer was based on the breakthrough in embryo freezing (Willadsen et al., 1978; Wilmut and Rowson, 1973). Right now, a serious setback of propagation of ovum pick-up/in vitro fertilization (OPU/IVF) technique is the inefficiency of traditional freezing for cryopreservation of in vitro-produced cattle embryos.
A decade of success and overwhelming data in human IVF (based on ∼1.5 millions babies born worldwide) were still insufficient to convince animal breeders that the right solution is the proper application of vitrification. Lack of possibility for direct transfer is often referred as the main problem, although, reports about feasible solutions have been published already (Vajta et al., 1999; Vieira et al., 2007; Zhang et al., 2015).
The low overall efficiency of somatic cell nuclear transfer generally discourages embryologists to include another potentially harmful step before embryo transfer. Accordingly, attempts and successes are relatively sparse. The first somatic cell-cloned mammal, born after embryo cryopreservation was reported in 2003 (Tecirlioglu et al., 2003). The healthy calf was produced with HMC, open pulled straw vitrification and in-straw dilution. During the subsequent 14 years, however, only a handful of publications reported similar achievements, including traditionally cloned transgenic bovine embryos (Gong et al., 2004); handmade cloned pig embryos (Du et al., 2007b); traditional cloned bovine embryos (Forell et al., 2008); traditionally cloned inter-subspecies buffalo embryo after traditional freezing (Yang et al., 2010); and handmade cloned buffalo embryos (Saha et al., 2013).
Additionally, reliable information was obtained about births from traditionally and handmade cloned sheep embryos (T.T. Peura, pers. comm.), traditionally cloned pig embryos (R.S. Prather, pers. comm.), and traditionally cloned cattle embryos (M. Maserati, pers. comm.). Two healthy offspring were also achieved with transfer of only eight vitrified handmade cloned bovine blastocysts (25%) (Polanco et al., in press). The concomitant rate achieved with fresh HMC embryos was 29%, two healthy offspring from seven transferred embryos.
Possibly some successes were not reported in full articles and are difficult to access. Still, the total number is disappointingly low, even compared with the limited quantity of cloned animals born worldwide during the past 20 years. It is worth considering that all, except for one, live births were achieved after vitrification; and the contribution of HMC in these births was considerably high, compared with the small number of laboratories using this cloning approach.
Convincing the Public
It is rarely realized by the public that—apart from the biological problems—a major hindrance cloners have to fight with is the difficulty to get the message delivered and understood both by laymen and professionals, including scientists and animal breeders, to decrease their concerns and to let us have a chance to prove the value of our approach.
The task seems to be easier now than 20 years ago, the hysteric atmosphere has more or less disappeared around cloning. People might have realized that there are far bigger and more pressing dangers for humankind to deal with, and cloners also avoided to cross the red line, respected the laws, and did not enter forbidden areas, although it is disputable whether it was really a self-restraint from everybody potentially involved, or just the inability to proceed in some cases.
In countries with no restriction to put product of clones or their descendants into the food chain, no significant civil protests arose. However, even if not expressed openly, most people still have concerns against clones and cloners, and explain their aversion with one or two arguments obtained from the media. There are appropriate answers supported by reliable scientific articles to most of these arguments, it is probably worth to collect and use them when required.
“Clones are not normal.” All human intervention in natural processes, including all medical treatments, may have unwanted effects. In the initial period of cloning, abortions, stillbirths, and malformations were often observed. However, it has been revealed soon that most (possibly all) of these unwanted outcomes were the result of the inappropriate technique, not the somatic cell nuclear transfer itself (Choi et al., 2002; Perry and Wakayama, 2002; Yanagimachi, 2002). After some controversies, it is now a wide consensus that “seemingly” healthy clones are really healthy, with no hidden developmental abnormalities (Heyman, 2005).
“Clones have shorter lifespan.” This argument is mostly based on the regretfully early death of the first sheep cloned. Retrospectively, it was mostly the result of a disease independent of cloning. The second cloned animal, a mouse, had lived longer than the average mice, and sheep clones created from the same cell line as Dolly had a lifespan identical with naturally conceived counterparts (Sinclair et al., 2016). There is no age-related attrition of learning ability or of strength, agility, and coordination in clones compared with noncloned animals (Tamashiro et al., 2000).
“Genetic defects may accumulate with cloning.” Naturally bred offspring of a clone (even of two clones) are completely normal (Heyman, 2005; Martin et al., 2004; Tamashiro et al., 2002). Bulls cloned from a top bull have the same performance regarding semen quality, in vivo and in vitro fertilizing ability, pregnancy, and calving outcome (Galli et al., 2003). Clinical parameters of offspring are normal and cloned animal products are suitable for human consumption (Heyman et al., 2007; Takahashi and Ito, 2004; Tian et al., 2005). Serial cloning of mice to six generations did not increase the number of genetic abnormalities (Wakayama et al., 2000).
“Product of cloned animals may be harmful to humans.” Extensive investigations in Japan, France, and the United States of America concluded that cloned animal products are suitable for human consumption (https://fda.gov/animalveterinary/safetyhealth/AnimalCloning/default.htm) (https://fsc.go.jp/english/evaluationreports/hy_detail_clone.pdf) (Heyman et al., 2007; Takahashi and Ito, 2004; Tian et al., 2005). No scientific data are available contradicting these investigations.
“Cloning is unnatural.” This tentative statement is difficult to fight with; it is more a philosophical than a biological issue. Analogs of somatic cell nuclear transfer do not occur in nature in mammals, but full reprogramming of somatic cells is a common phenomenon in plants, also used by humans in agriculture since millennia. Cloning can be regarded as a special way to reprogram somatic cells, similar but more efficient than other techniques to create—more or less—totipotent stem cells. In practice, cloning is just an assisted reproductive technology. Due to its special features, it is more strictly regulated than other reproductive technologies. Scientists acknowledge and accept these restrictions.
Convincing Professionals
Decision makers in legislation, academic institutions, or private sectors, including investment agencies, pharmaceutical, or animal breeding companies, may have similar private concerns, but they mostly hide them on professional discussion. Their arguments are more focused and—controversially—easier to answer. However, even the most adequate reply may still be insufficient to change their intrinsic negative opinion.
In this study, we collected statements related to cattle cloning for industrial purposes—the reason of this focus is explained in the subsequent chapter.
“Requires expensive investment, high running costs, requirement for specially qualified workforce in the laboratory.” Although, numbers at first look may be discouraging, the infrastructure, equipment, and running costs are similar to that of a cattle OPU/IVF laboratory, a technology widely used in both North and South America, and present in all developed countries. The extra investment for equipment required for traditional cloning is considerable (around USD 150,000–200,000), but the application of HMC may reduce these costs to approximately USD 5000.
It should also be considered that oocytes used for cattle cloning are derived from ovaries of slaughtered cows, therefore, the available quantity is almost unlimited and costs are negligible. Unlike artificial insemination and transfer of in vivo-derived or in vitro-produced embryos, no costs occur with male gametes, and isolation/maintenance/storage of cell lines required for nuclear transfer require modest efforts and expenses. For chemicals, only a handful of items are specifically required for cloning, and in such a small quantity that the related cost of one cloning program is less than that of a modest family dinner. The same is applicable to disposables. Apart from the salaries, the highest cost of laboratory production of cloned embryos is probably the price of petrol to transport ovaries from the slaughterhouse to the laboratory.
For personnel, the requirement for highly qualified workforce is unquestionable and unavoidable—similarly to all fields of assisted reproduction. With proper training and supervision, all capable embryologists can learn and use the technology.
“Extremely low overall efficiency.” Various numbers spreading in the press are really disappointing. However, a fair comparison with natural reproduction reveals that cloning is not as inefficient as generally supposed. Considering that one cow may produce 50–100 M-I phase oocytes per month, but only one of them can develop to offspring, cloning with the same oocytes producing (the often referred) 1% overall success rate is quite acceptable (it should be emphasized again that some laboratories report considerably higher percentages; see earlier chapters).
On the other hand, for commercial application in an established unit, the cost of embryo production is a matter of less moment than the expenses related to embryo transfer: recipients, transportation, intervention, and follow-up. Accordingly, healthy calf on the ground per embryo transfer rate is the decisive characteristic. In this study, cloning in general is rather handicapped with its 10% results achieved by most commercial applications, whereas transfer with in vivo-derived or in vitro-produced embryos may reach 50–60 or 40%–50%, respectively. This is definitely a serious setback hampering widespread use; it increases costs, and causes disproportional frustration for both professionals and clients. To improve the success rate to 20, rather 30% is an absolute requirement. Fortunately, as discussed below, there are proven and/or promising ways to achieve this goal.
“Cloning does not create improved genetics, just copies—and widespread application of cloning may destroy genetic diversity.” The first argument is intrinsically senseless—considering real-life situations. In a pure, entirely homogenous population, cloning would not help—but that is also more or less applicable for all assisted reproductive technologies. However, cattle stock is far from homogenous, even in the most developed countries, including North America, Europe, and Australia; not speaking about South America, Africa, and the whole Asian–Pacific region. A typical situation in these countries is a sharp difference between a few elite animals or farms, and the average stock. In this situation, cloning may be uniquely suitable to increase the genetic value in one generation to a top level.
Regarding the diversity, application of proper breeding schemes may perfectly eliminate the supposed danger. In fact, it would be a great achievement of cloning to become considered as a danger for genetic diversity. With the present level of application, this danger is comparable to that of overpopulating Mars by the first spaceship sent there. Quite in contrast to this fear, cloning may contribute—and has already contributed—to maintain genetic diversity by saving last survivors of seriously endangered breeds (Kim et al., 2013; Wells, 2000; Wells and Misica, 1998). This application—opposed to saving endangered or extinct species—is quite realistic and respectable, with immediate and unquestionable positive impact.
A Potential Area for Breakthrough—Cattle Breeding
Apart from obtaining the public and professional support, cloners also need to demonstrate the real value of somatic cell nuclear transfer in an area where success is predictable, requires relatively modest efforts, and the benefit is evident for both investors and the public. Biomedical application is a long and demanding procedure, and the end users may steal the show from cloners. Saving of wild species or recreation of extinct ones with somatic cell nuclear transfer is currently over the realities. Domestic animal—especially cattle—breeding, however, may meet most of the requirements listed above. Cattle have a privileged position in animal husbandry. It is the largest domestic species bred in huge numbers, with ∼1.4 billion animals worldwide. Its impact to the world economy is enormous considering the value of products, the offered workplace, and the contribution in food supply.
The reproductive cycle in cattle is long, and the 9 months pregnancy results usually only a single offspring. This slow natural reproduction justifies artificial intervention to speed up genetic advancement. Accordingly, an extensive knowledge has been accumulated in this field during the past century, and commercial application of assisted reproductive techniques is widely accepted. Embryos can be placed with a relatively simple nonsurgical, transvaginal transfer method into the uterus, and a single embryo is usually enough to establish a pregnancy leading to birth even after cloning (Cortez et al., 2018), as opposed to pigs, where surgical or laparoscopic intervention and transfer of 40–80 embryos is required. The financial advantage—a single embryo may produce a 400–1000 kg offspring with potential multiple use (beef, milk, skin) and with the required quality—is obvious.
When the first somatic cell-cloned sheep was published in Scotland, there were already pregnancies with cloned cattle in the United States (Cibelli et al., 1998), and bovine application was almost immediately considered as the most practical and feasible one. However, in spite of—first considerable, later sporadic—efforts during the past 20 years, the possibility is still unexploited, and the impact of cloning in cattle breeding right now is negligible.
The following tentative list contains the most important areas of cattle breeding, where cloning may have of considerable impact—without an in-depth analysis of details that are outside of the scope of this review.
(i) Elite and highly valuable animals: Individual cloning may multiply the amount of collectable gametes (sperm cells or oocytes) for more traditional assisted reproductive technologies (artificial insemination, embryo transfer, IVF); or just to create a kind of insurance for accident or disease for the most valuable animals of a given stock. Cloning from carcasses selected after slaughter may also help to multiply beef cattle with very special value.
(ii) Good-quality animals: Large-scale cloning may improve the genetics of a handicapped stock for further breeding, or just for end products at beef cattle. Cloning is a perfect way to produce animals with the required gender, that is, heifers for dairy, and bulls for beef herds. Some special application areas, including mass production of good-quality bulls for natural insemination of Northern herds of Australia, may also be considered.
(iii) Critically endangered breeds—as discussed above.
(iv) Cloning to save the environment: Global warming is a central issue for humankind, and enormous efforts are made to decrease industry, household, and transportation-related carbon emission. Unfortunately, much less is happening to decrease emission related to agriculture (estimated 14%–18% by various studies; more than the whole transportation factor), where pollutants related to cattle are far the most significant ones (Friel et al., 2009).
There are several ways to decrease this pollution. Some of them are difficult to realize (collection of gasses from each individual animal), others may have some food safety issues (genetic modification). The most feasible ways to decrease the harmful effect are (i) to propagate rapidly animals that are producing less polluting gasses (de Haas et al., 2017), and (ii) to improve the quality of the stock. For the latter, it is quite understandable that a single elite cow producing 15,000–20,000 kg milk per year will have less impact on global warming than 7–8 individuals with the world's average productivity (around 2200 kg/year).
Both financial and environmental points urge us to use all available methods to accelerate rapid propagation of highly productive cattle worldwide. Cloning should be seriously considered as a possibility to contribute in this process.
Conclusion
Cloning by somatic cell nuclear transfer has great potential to answer basic questions in developmental biology and gene regulation, and to provide new solutions in various commercial areas, including biomedicine and animal—especially cattle—breeding. These possibilities have remained unexploited, mostly because of external factors that discouraged or effectively stopped research and development in this field. Elimination of mental and administrative barriers, unbiased evaluation of pro's and con's, and more confidence in success would be required to let somatic cell nuclear transfer to prove its real value.
Footnotes
Author Disclosure Statement
The author is director and co-owner of a consulting company that is involved in teaching, propagation, and commercialization of open pulled straw (OPS) vitrification and handmade cloning (HMC).