
Abstract
Abstract
Accumulating evidence indicates that mesenchymal stem cells (MSCs) have been widely used in tissue engineering and regenerative medicine due to their multilineage differentiation potentials. Recent studies show that germ-like cells can also be derived from stem cells, such as human umbilical cord MSCs and human bone marrow MSCs in vitro. However, whether human adipose-derived MSCs (hAD-MSCs) can be induced into germ-like cells has never been reported. In this study, we isolated hAD-MSCs and confirmed that their characteristics were in accordance with that of MSCs established before. Germ cell lineage differentiation was performed by 10 μM retinoic acid (RA) treatment for 21 days. RA induction led to slender spindles and tadpole-like changes of cell morphology, and the expression of germ cell-specific markers (Oct4, Piwil2, Itgb1, SSEA-1, and Stra8) presented significant upregulation in the RA treatment group according to the polymerase chain reaction and immunofluorescence results. We first demonstrated that hAD-MSCs can differentiate into germ-like cells in vitro, which will provide theoretical and experimental basis for the clinical application of hAD-MSCs in the treatment for infertility.
Introduction
W
It has been reported that human adipose-derived MSCs (hAD-MSCs) possess pluripotent characteristics for indefinite proliferation and they can differentiate into multiple cell types, including osteoblasts, chondrocytes, neural cells, and cardiomyocytes (Witkowska-Zimny and Walenko, 2011). Different from other sources of stem cells, hAD-MSCs is abundant, easily harvested from adipose tissue, and can be expanded rapidly in vitro (Hauner, 2004). However, whether hAD-MSCs could differentiate into germ cells (GCs) by retinoic acid (RA) treatment still remains undetermined.
In this study, we isolated MSCs from adipose tissue and investigated the characteristics of hAD-MSCs. The main point of this study is that hAD-MSCs can be induced by RA to express germ cells markers, which can help to understand germ cell differentiation from stem cells and will bring benefits to infertility families.
Materials and Methods
Isolation and expansion of hAD-MSCs
Human adipose tissue from three healthy donors 24–32 years of age was obtained from liposuction surgery with an informed written consent. All the studies were approved by the Ethics Committees of the hospitals and the Institutional Review Board of Jining Medical University and carried out in accordance with their approved guidelines. hAD-MSCs were isolated as previously described (Hauner, 2004). Briefly, adipose tissues was washed in sterile phosphate-buffered saline (PBS) at least once and digested by 0.1% collagenase type I (Gibco) for 30 minutes at 37°C with agitation. After centrifugation (1000 g for 15 minutes), the liquid was separated and the cellular pellet appeared at the bottom of the liquid layer. Then the cell suspension was filtered through a nylon mesh with a pore size of 100 μm.
Pellets were washed three times in PBS and were resuspended in DMEM/F-12 (Hyclone) medium supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco). Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2. The medium was changed every 3–4 days. When cells reached 80%–90% confluences, they were harvested using 0.25% trypsin-EDTA (Gibco) and dispensed at a ratio of 1:3 into new culture flasks.
Evaluation of cell surface markers
hAD-MSCs were harvested at passage 3. The cells were detached by trypsinization, rinsed twice with PBS, and resuspended in 100 μL PBS at a concentration of about 1 × 105 cells. Collected cells were incubated with CD73-FITC (fluorescein isothiocyanate), CD34-PE (phycoerythrin), CD105-PerCP (peridinin chlorophyll protein), HLA-DR-FITC, CD90-PE, and CD45-PerCP monoclonal antibodies (BioLegend) for 15 minutes in the dark. After staining, the cells were washed with 1 mL PBS, resuspended in 300 μL PBS, and analyzed by flow cytometry (BD Biosciences). The mouse IgG1-isotype was used as a negative control.
Osteogenic and adipogenic differentiation
Osteogenic induction: The hAD-MSCs of passage 3 were plated in six-well plates at a density of 1 × 105 cells/cm2 and cultured with H-DMEM (Gibco) induction medium containing 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 10 nM dexamethasone (Sigma), 50 mol/L ascorbic acid (Sigma), and 10 mmol/L β-glycerophosphate (Sigma). Cells cultured in complete H-DMEM medium without induction agents were taken as control. The medium was changed every 3 days.
After 21 days, the cells were washed twice with PBS, fixed with 4% paraformaldehyde for 20 minutes and stained with Alizarin Red (Sigma) to detect the presence of calcium deposition in osteocytes. Adipogenic induction. The hAD-MSCs of passage 3 were plated in six-well plates at a density of 1 × 105 cells/cm2 and cultured with H-DMEM (Gibco) induction medium containing 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1 μmol/L dexamethasone, 0.2 mmol/L indomethacin (Sigma), 10 g/mL insulin (Sigma), and 0.5 mmol/L 3-isobutyl-1-methylxanthine (Sigma). Cells cultured in complete H-DMEM medium without induction agents were taken as control. The medium was replaced every 3 days for 14 days. At 14 days of induction, the cells were washed twice with PBS, fixed with 4% paraformaldehyde for 20 minutes, and stained with Oil Red O (Sigma) to detect the presence of neutral lipid vacuoles in adipocytes.
Induction of germ cells from hAD-MSCs
hAD-MSCs of passage 3 were cultured in H-DMEM induction medium containing 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, and 10 μM RA (Sigma). The medium was replaced every 3 days. After 21 days of RA induction, cell morphology and GC-specific markers were evaluated by immunocytochemistry and polymerase chain reaction (PCR).
Reverse transcription–PCR and quantitative RT-PCR
Expression of germ cell specific genes was detected by reverse transcription (RT)–PCR. Moreover, quantitative RT-PCR (qRT-PCR) was used to verify expression changes of GC-specific markers. Total RNA was extracted from cells using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. For RT-PCR analysis, 1 μg of total RNA was reverse transcribed into complementary DNA (cDNA) using the RevertAid™ First-Strand cDNA Synthesis Kit (Thermo Scientific, Bremen, Germany) according to the manufacturer's instructions.
The PCR mixture contained 1 μL cDNA, 0.4 μL forward primer, and reverse primer, respectively, 10 μL 2 × Taq PCR Master Mix (Tiangen) in a total volume of 20 μL with distilled water. The PCR conditions were 5 minutes at 95°C, then 45 cycles of 95°C for 30 seconds, 60°C–65°C for 30 seconds, 72°C for 30 seconds, and a final extension for 10 minutes at 72°C. The expression of GC-specific genes was detected by qRT-PCR using an Applied Biosystems 7500 Real-time PCR System and SYBR Green Master Mix (TransGen Biotech). All the primers used in this study are listed in Table 1.
Immunofluorescence staining
Coverslips were placed into a six-well plate and then hAD-MSCs were plated. After incubation and RA induction, medium was removed and the cells were washed twice with PBS and fixed with 4% paraformaldehyde for 20 minutes. Then cells were washed three times in cold PBS for 3 minutes and treated with 1% Triton X-100 for 10 minutes at room temperature, followed by incubating with 5% bovine serum albumin (BSA) in PBS for 20 minutes. Then, the cells were incubated with primary antibodies (Oct4, 1:100, Cat. No. ab1985; beta-actin, 1:100, Cat. No. 60008-1-Ig) at 4°C overnight. Then the cells were washed three times in PBS, followed by incubation with Alexa Fluor 488/FITC-conjugated goat anti-rabbit/mouse secondary antibody for 2 hours at room temperature.
Cells were then immediately rinsed four times with PBS for 5 minutes each. After slides had air dried completely, the stained cells were covered with glass slides using mounting medium, DAPI (4′, 6-diamidino-2-phenylindole) fluoromount G (SouthernBiotech). Fluorescence images were acquired with a LSM 800 confocal microscope (Zeiss) using Zeiss Confocal Software.
Statistical analysis
Data are presented as mean ± SD and results were analyzed by the student's t-test. SPSS21.0 software was used for data analysis. p < 0.05 was considered statistically significant.
Results
Isolation and characterization of hAD-MSCs
MSCs were isolated from human adipose tissue. After 3 days of primary culture, the adherent fibroblast-like cells were observed under phase-contrast microscope. The medium was changed every 3 days until the cells reached 90% confluence (about 7–8 days). Cells were passaged at a dilution of 1:3 and reached confluence again within 3–4 days. hAD-MSCs were shown as spindle-shaped cells and the morphology did not change during cell passages (Fig. 1). To determine whether the adherent cells from adipose tissue met the quantifying criteria of MSCs, we analyzed the expression of surface antigens (CD73, CD90, CD105, CD34, CD45, and HLA-DR). As shown in Figure 2, stromal cell markers (CD73, CD90, and CD105) were expressed in hAD-MSC, with a high positivity rate. However, hematopoietic cell markers (such as CD45, CD34, and HLA-DR) were not expressed. All the surface markers' expression detected in hAD-MSCs conformed to the criteria of MSCs established before (Dominici et al., 2006).

A-D: Morphology of hAD-MSCs. Primary culture of hAD-MSCs at day 3
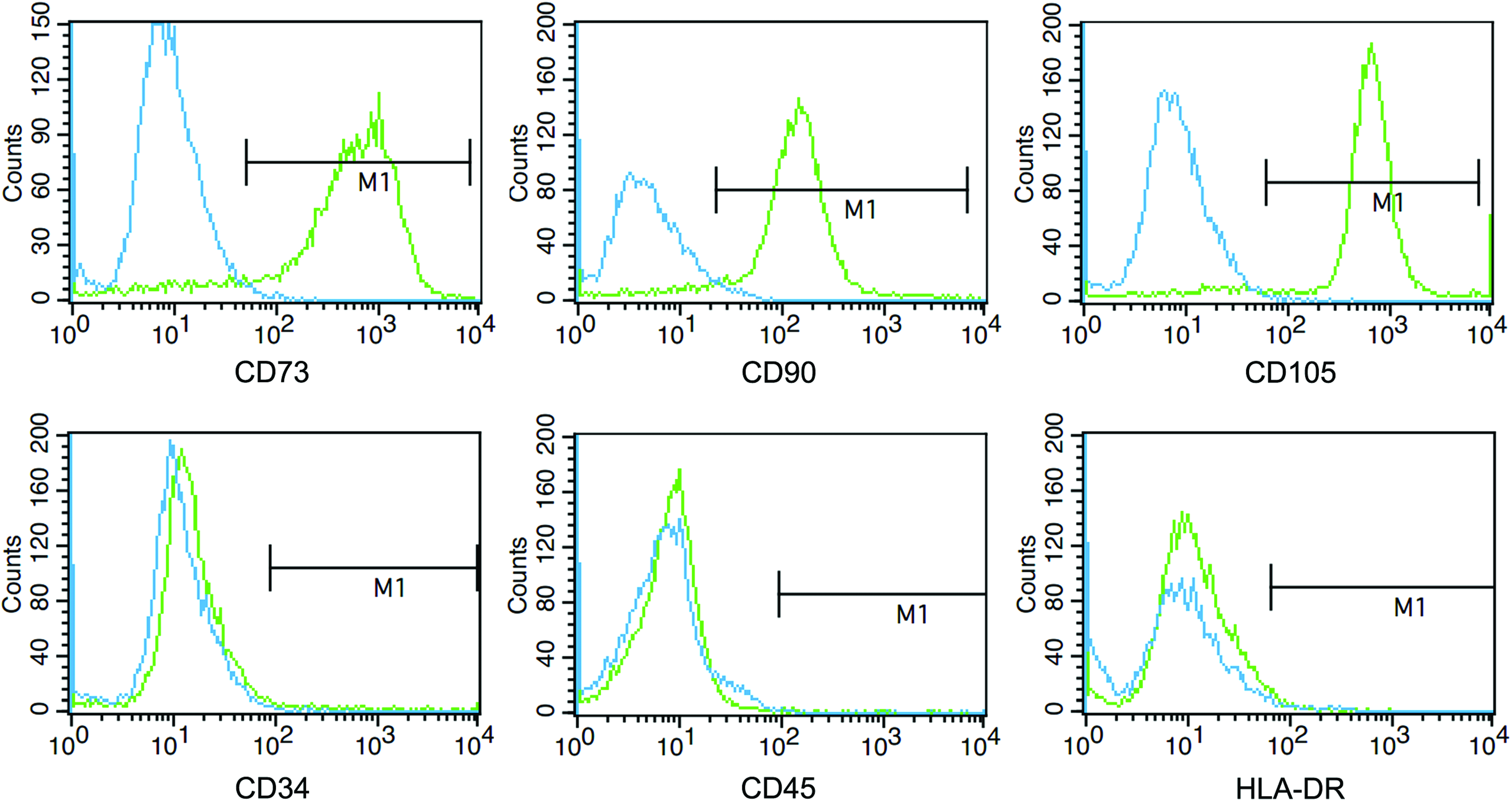
Flow cytometry analysis. Cells of passage three (P3) were collected and labeled with PE or FITC-conjugated antibodies. The cells were positive for CD73, CD90, and CD105 and negative for CD34, CD45, and HLA-DR. Color images available online at www.liebertpub.com/cell
Osteogenic and adipogenic differentiation of hAD-MSCs
We explored the potential of hAD-MSCs to differentiate into osteoblast and adipocytes using protocols as previously described (Han et al., 2014). After osteogenic induction for 21 days and adipogenic induction for 14 days, hAD-MSCs could differentiate into osteoblasts or adipocytes as identified by Alizarin Red staining or Oil Red O staining, respectively (Fig. 3), which validated the multilineage differentiation potentials of hAD-MSCs.

Osteogenic and adipogenic differentiation of hAD-MSCs. hAD-MSCs were incubated with or without osteogenic medium for 21 days and stained with Alizarin Red
Morphological changes of hAD-MSCs after RA induction
To induce hAD-MSCs into germ cells, hAD-MSCs were treated with RA and morphological changes were observed every day under a phase-contrast microscope. The morphology of hAD-MSCs under RA induction gradually changed from fibroblast to slender spindles and formed a tadpole-like shape within 7 days, which did not change significantly in the subsequent induction (Fig. 4).

Morphological changes of hAD-MSCs with and without RA induction for 7, 14, and 21 days. All the pictures were captured using the IX71 Olympus microscope under 10 × magnification. RA, retinoic acid. Arrows indicate the successfully induced germ cells.
RA-induced expression of germ cell-specific gene in hAD-MSCs
After 21 days of RA induction, RT-PCR was first performed to detect the expression of germ cell-specific genes, Oct4, Piwil2, Itgb1, SSEA-1, and Stra8. RA induction at day 21 significantly increased the expression of germ cell-specific genes compared with that at day 0 (Fig. 5A). Next, the results were further validated by quantitative real-time PCR (Fig. 5B), which together demonstrated that RA, to some degree, could induce hAD-MSC to differentiate into germ-like cells.
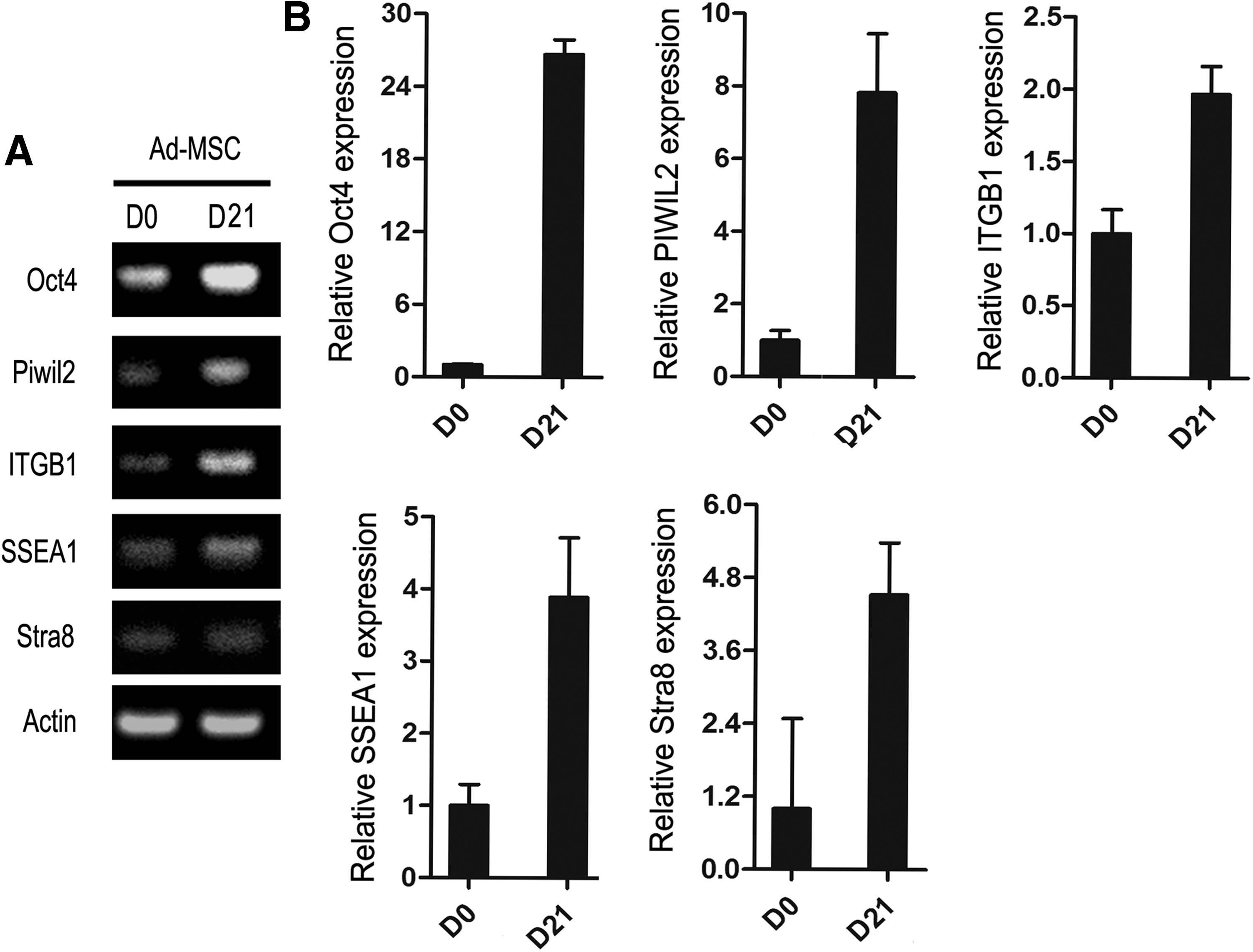
Expression of germ cell-specific genes in hAD-MSCs with and without RA treatment.
Oct4 is recognized as a marker of pluripotency and its expression could also be used to evaluate germ cell differentiation. In this study, Oct4 immunostaining at day 0 and 21 of RA induction were performed to further confirm the differentiation of hAD-MSC to germ-like cells. Beta-actin immunostaining was used to reflect the morphology changes of hAD-MSC before and after RA induction. As shown in Figure 6, more slender spindle-like cells after RA induction were observed according to beta-actin immunostaining result, and RA induction significantly induced Oct4 expression, which is in agreement with PCR results (Fig. 5).

Immunofluorescence staining of hAD-MSCs with and without RA treatment for 21 days.
Discussion
MSCs have the multidirectional differentiation capacity and self-update, which can differentiate into a variety of cell types, such as osteoblasts, chondrocytes, and adipocytes (Dominici et al., 2006; Pittenger et al., 1999). MSCs can also be induced to differentiate into germ-like cells after appropriate induction in vitro (Hua et al., 2009a; Liu et al., 2014), which makes it a promising choice for the treatment of infertility. Adipose tissue is one of the richest sources of adult stem cells, which can be easily obtained through liposuction. The isolation of MSCs from adipose tissue is much easier and has no risk for the donors without any ethical issues.
In this study, we isolated MSCs from human adipose tissue and the morphology and cell surface markers are all in accordance with that of MSCs identified before (Jiang et al., 2013; Meligy et al., 2012). Moreover, we also demonstrated the differentiation potentials of hAD-MSCs into osteocytes and adipocytes, which are also similar to the previously reported literature (Ghasemzadeh-Hasankolaei et al., 2015).
In 2006, Nayernia and his team first reported that mouse BM-MSCs can be induced into male GCs using 1 and 10 μM RA (Nayernia et al., 2006b). After that, various reports showed that MSCs derived from hBM, lung, and umbilical cord Wharton's jelly can also differentiate into germ-like cells using different induction regimens, such as RA, RA with testicular cell-conditioned medium, BMP4 and BMP8, and transforming growth factor beta 1 (TGFb1) (Drusenheimer et al., 2007; Hua et al., 2009a, 2009b) (Drusenheimer et al., 2007; Ghasemzadeh-Hasankolaei et al., 2015; Mazaheri et al., 2011; Shirazi et al., 2012).
RA, an active derivative of vitamin A, is produced in both sexes in mesonephros (Bowles et al., 2006; Bowles and Koopman 2007). RA has been reported to influence the differentiation of stem cells into GCs and stimulate mitosis and meiosis of both female and male germ cells (Chen et al., 2012; Geijsen et al., 2004; Hua et al., 2009a; Koubova et al., 2006; Nayernia et al., 2006a).
In this study, we successfully induced hAD-MSCs into germ-like cells using 10 μM RA alone for the first time. We confirmed the differentiation of hAD-MSCs into germ-like cells through observing the cell morphological changes and detecting the expression of GC-specific markers at the mRNA (Oct4, Piwil2, Itgb1, SSEA-1, and Stra8) and protein (Oct4) levels. Our results were similar to the reports mentioned above.
Oct4 belongs to the POU (Pit-Oct-Unc) transcription factor family (Drusenheimer et al., 2007). It is a marker of pluripotency that plays an important role in embryonic development. Oct4 can also be detected in both pluripotent cells and early germ cells (Allegrucci et al., 2005; Nagano, 2007). Piwil2 is a GC marker that has an important role in the self-renewal of SSC and spermatogenesis. Moreover, it is necessary for the repair of DNA damage (Lee et al., 2006; Yin et al., 2011). Itgb1 is a marker of SSCs and spermatogonia. It is closely related to metastasis and homing of PGCs and normal germline development (Lacham-Kaplan, 2004; Shinohara et al., 1999).
It has been reported that SSEA-1 is a marker of early PGCs, whose increased expression can be used to evaluate germ-like cell differentiation of MSCs (Li et al., 2014). Stra8 is considered to be a GC-specific gene, which is highly expressed at prophase of meiosis and absent in male embryonic GCs (Aflatoonian and Moore, 2006; Lacham-Kaplan, 2004; Zhou et al., 2008). In this study, Oct4, Piwil2, Itgb1, SSEA-1, and Stra8 were chosen as GC-specific markers and their expressions were detected before and after RA induction to evaluate the GCs differentiation from hAD-MSCs. Our results showed that all the selected markers displayed increased expression after RA treatment for 21 days with statistical significance, which altogether demonstrated the successful germ-like cell induction from hAD-MSCs by 10 μM RA treatment.
In summary, we first demonstrated that hAD-MSCs can differentiate into germ-like cells in vitro, which will provide theoretical and experimental basis for the clinical application of hAD-MSCs in the treatment for infertility.
Footnotes
Acknowledgments
This study was supported by the Youth Foundation of Jining Medical University (grant No. JYQ14KJ30), the Natural Science Foundation of Shandong Province (No. ZR2017LH029), the Science and Technology Program of Jining City, China (No. 2016-56-104), and the PhD start-up funds from the Affiliated Hospital of Jining Medical University (No. 2016-BS-001, 2016-BS-003).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.