
Abstract
Abstract
Biobanks of cryopreserved gametes and embryos of domestic animals have been utilized to spread desired genotypes and to conserve the animal germplasm of endangered breeds. In principle, somatic cells can be used for the same purposes, and for reviving of animals, the somatic cells must be suitable for animal cloning techniques, such as somatic cell nuclear transfer. In the present study, we derived and cryopreserved somatic cells from three breeds of riverine and swamp-like type buffaloes and established a somatic cell bank. In total, 350 cryovials of 14 different individual animals (25 cryovials per animal) were cryopreserved and informative data such as breed value, origin, and others were documented. Immunostaining of the established cells against vimentin and cytokeratin suggested a commitment to the fibroblast lineage. In addition, microsatellite analysis was performed and documented for unambiguous parentage verification of clones in the future. Subsequently, the cryopreserved cells were tested for their suitability as nuclear donors (n = 7) using handmade cloning, and the reconstructed embryos were cultured in vitro. The cleavage rates (95.99% ± 2.17% vs. 82.18% ± 2.50%) and blastocyst rates (37.73% ± 1.54% vs. 24.31% ± 1.78%) were higher (p < 0.05) for riverine buffalo cells than that of swamp-like buffalo cells, whereas the total cell numbers of blastocysts (258.16 ± 36.25 vs. 198.16 ± 36.25, respectively) were similar. In conclusion, we demonstrated the feasibility of biobanking of buffalo somatic cells, and that the cryopreserved cells can be used to produce cloned embryos. This study encourages the development of somatic cell biobanks of domestic livestock, including endangered breeds of buffalo, to preserve valuable genotypes for future revitalization by animal cloning techniques.
Introduction
B
Somatic cells hold various advantages compared to gametes and embryos, (a) tissue biopsies can be easily obtained from animals of both sexes, (b) tissue biopsies can be obtained in remote areas, and (c) somatic cells can be cultured from tissues samples of old or even from dead animals, shortly after death (Holt and Pickard, 1999; Leon-Quinto et al., 2009). Somatic cells of domesticated animals have been preserved in the biobanks, and few examples of such banks are available in Vietnam (Groeneveld et al., 2008) and Turkey (Arat et al., 2011). It is also proved that frozen cells of biobanks could be used to produce the cloned animals (Arat et al., 2011). However, the biobanking of somatic cells of domestic animals is currently not popular and not many such biobanks are available (Groeneveld et al., 2016).
India holds one of the world's most diverse population of livestock species and possesses more than 100 breeds of livestock (cattle, buffalo, goat, sheep, and pig), which have significantly been contributing to the economy by producing milk, meat, and fibers (Handbook of Animal Husbandry (4th ed.), 2013. ICAR, Delhi, India). In India, 49 semen production banks of domestic animals are currently active with an aim to propagate the germplasm of best animals through artificial insemination and also support the conservation of germplasm (Annual report of Department of Animal Husbandry, Dairying & Fisheries, Gov. of India published in 2016–2017).
At present, more than 1 million semen doses of 311 breeding males of 44 breeds of livestock (cattle, buffalo, sheep, goat, camel, yak, and equine) were cryopreserved as backup materials for reintroduction of lost germplasm in the future. (Annual report of National Bureau of Animal Genetic Resources, India published in 2015–2016). In addition to gametes or embryos, somatic cells can also support the conservation of animal genetic resources as they carry complete genome and proteome and can be used to reintroduce the lost germplasm using animal cloning techniques. Such biobanks can also provide biomaterials for research studies, such as whole-genome sequencing approaches, and save the time of researchers, which they devoted for the culturing of somatic cells in the laboratory.
Many Indian breeds of livestock become endangered due to decreasing demand in the commercial markets. We suggest the preservation of somatic cells of best indigenous and endangered breeds of livestock. Considering this in mind, we cryopreserved somatic cells of 14 buffaloes from three different breeds, and the established cells were characterized and assessed for the suitability of the cryopreserved cells as nuclear donors in animal cloning and their competence for blastocyst development.
Material and Methods
Animal experiments were approved by the Institute Animal Ethics Committee, ICAR-Central Institute for Research on Buffaloes, Hisar, India. Buffaloes were selected for the collection of tissue biopsies by considering their pedigree records, breed characteristics, and production performance (milk production for females and standard breeding scale for bulls). In case of field buffaloes, the written consensus from farmers has been signed before the collection of tissue biopsies from their animals. The culture of somatic cells, oocytes, and embryos were performed at 38.5°C in a 5% CO2 incubator. Chemicals and culture media were obtained from Sigma Chemical Company (St. Louis, MO) and disposable plastic wares from Nunc (Roskilde, Denmark), unless otherwise noted.
Isolation and culture of somatic cells
Skin tissue biopsies, from the underneath part of the tail, just above anal region (12 buffaloes), and from the ear pinna (2 buffaloes) were collected using a biopsy punch to establish cultures of somatic cells (Supplementary Fig. S1 Supplementary Data are available online at www.liebertpub.com/cell shows a site of biopsy collections from tail).
In India, swamp-like buffaloes are available in the north eastern part of the country, which is ∼2000 km away from our laboratory location; therefore, tissue biopsies of swamp-like buffaloes were airlifted and transported to the laboratory within 24 hours of the collection. Collected biopsies were transported in Dulbecco's phosphate-buffered saline fortified with Ca2+ and Mg2+ salts (DPBS+) to the laboratory and washed four to five times with DPBS+. The biopsies were chopped into small pieces (about 1 mm in size per piece) using a sterile surgical blade and washed three times with the Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS).
The tissue explants were seeded in 15 μL drops of DMEM supplemented with 20% FBS, 1% (v/v) nonessential amino acids, 1 × vitamin mix (Himedia, VA001, Mumbai, India), and 1 × antibiotic solution (GIBCO, 15240–062, Grand Island, NY), hereafter called culture medium, in 25 cm2 culture flasks at 38.5°C in a 5% CO2 incubator (three to five explants in one flask). After 10–12 hours of seeding, 3 mL of fresh culture medium was added and kept for culturing. Cell outgrowths were appeared after 7–10 days of the explant seeding. The growth of cells was observed until they reach 70%–80% confluence, after that, they were trypsinized and subcultured.
Characterization of cells
The established cells were characterized using immunofluorescence staining with antibodies against vimentin (marker of fibroblasts), cytokeratin-18 (marker of epithelial cells), and tubulin (expressed in all cells). In brief, early passages (4 or 5) cells were cultured until 70%–80% confluence in the four-well culture dishes. Cells were fixed using 4% formaldehyde solution at 37°C for 1 hour. Fixed cells were permeabilized by 0.5% Triton-X for 30 minutes, blocked with 3% bovine serum albumin in DPBS, followed by overnight incubation with primary mouse antibodies (antivimentin 1:1500, V6630, Sigma, and anticytokeratin 18, 1:1500, MAB1611, Millipore) at 4°C, and then with the secondary antibody (goat anti-mouse IgG) conjugated with fluorescein isothiocyanate at 37°C for 1 h.
Positive controls used for testing the staining protocol were labeled with mouse antitubulin (1:1500, T8328, Sigma), whereas negative controls consisted of the omission of primary antibody. Nuclei were stained by Hoechst-33342 (10 μg/mL) for 10 minutes. Fluorescence signals were captured using an epifluorescence microscope (Nikon Ti-E eclipse, Tokyo, Japan), and images were merged using Adobe Photoshop CS 8.0.1 software (Adobe Systems, Inc., San Jose, CA).
Cryopreservation and cataloging of cells
After characterization, cells (passages 5–10) were cryopreserved using a slow freezing method as previously described (Selokar et al., 2014). In brief, 100% confluent cells were trypsinized and washed with DPSB+. Cell pellets were dissolved in 1.0 mL freezing medium consisting of 10% DMSO, 20% FBS, and 70% DMEM and transferred into 1.5 mL cryogenic vials. All vials were labeled with a unique animal ID, animal sex, date of freezing, and passage number. Cells in the freezing medium were subjected to slow freezing, overnight at a rate of 1.0°C/min to −80°C in freezing containers (Mr. Frosty™ freezer, Cat # 5100-0001; Thermo Scientific Pvt. Ltd. Whitefield, Bangalore, India). In one vial, 105 cells were cryopreserved.
After overnight storage, cryovials were quickly transferred into liquid nitrogen for the long-term storage. Three to five cryovials were cryopreserved per freezing day, and 25 cryovials per animal were preserved. After 1-week of storage in liquid nitrogen, one cryovial from an individual freezing day was quickly thawed in the water bath at 37°C and washed once with the culture medium. Thawed cells were cultured in 25 cm2 culture flasks in a 5% CO2 incubator. Sixty to seventy percent attachment rate and proliferation of attached cells were criteria for long-term storage, whereas, poorly frozen vials were discarded. Attachment rate was checked after 24 hours of culture.
Microsatellite analysis
DNAs from culture cells were isolated using a method described in a commercially available kit (PureLink® Genomic DNA Mini Kit, Catalog no.: K18200; Thermo Scientific Pvt. Ltd. Whitefield). Concentration and purity of isolated DNAs were checked using a spectrophotometer (PICOPET01, Picodrop Ltd. Hinxton, United Kingdom). One microgram DNA per animal was dispatched to CALF Laboratory, National Dairy Development Board (NDDB), Anand, India for analysis of microsatellite alleles. The CALF laboratory of NDDB has been commercially performing analysis of microsatellite alleles to confirm parentage of cattle and buffalo in India. We have used their service for generation of size of 13 microsatellite alleles.
Production of cloned embryos
Cryopreserved cells were thawed as mentioned above and cultured in culture medium in a CO2 incubator for one day before being used as donors in somatic cell nuclear transfer (SCNT). Handmade cloning method of SCNT was performed as previously described (Selokar et al., 2011). In brief, riverine buffalo ovaries were collected from Delhi slaughterhouse and transported to laboratory within 6–8 hours. Due to nonavailability of swamp-like buffalo ovaries, swamp-like buffalo cells were fused with the enucleated recipient oocytes of riverine buffaloes for production of cloned embryos. Oocytes were manually aspirated using 18G needles and subjected to in vitro maturation for 21 hours. Cumulus cells of matured oocytes were stripped off using hyaluronidase (0.5 mg/mL), and zona pellucida was digested using pronase (2.0 mg/mL) treatments.
Protrusion cone-guided manual bisection of zona-free oocytes was performed using a microblade in tissue culture medium 199 (TCM medium) supplemented with cytochalasin (2.5 μg/mL), followed by confirmation of enucleation using Hoechst 33342 staining. Enucleated cytoplasts were selected as recipient cytoplasts for the generation of cloned embryos. Enucleated cytoplasts were dipped in phytohemagglutinin (0.5 mg/mL) solution for 5–10 seconds and gently dragged over a one fibroblast for attachment. BTX Electrocell Manipulator 2001(BTX, San Diego, CA) machine was used to fuse cytoplast–fibroblast pairs with cytoplasts (sandwich electrofusion as shown in Fig. 1a). For fusion, 4 V AC pulse was used to align cytoplast-donor cell pairs with cytoplasts at vertical position, followed by the single pulse of 160 V of DC for cell fusion.
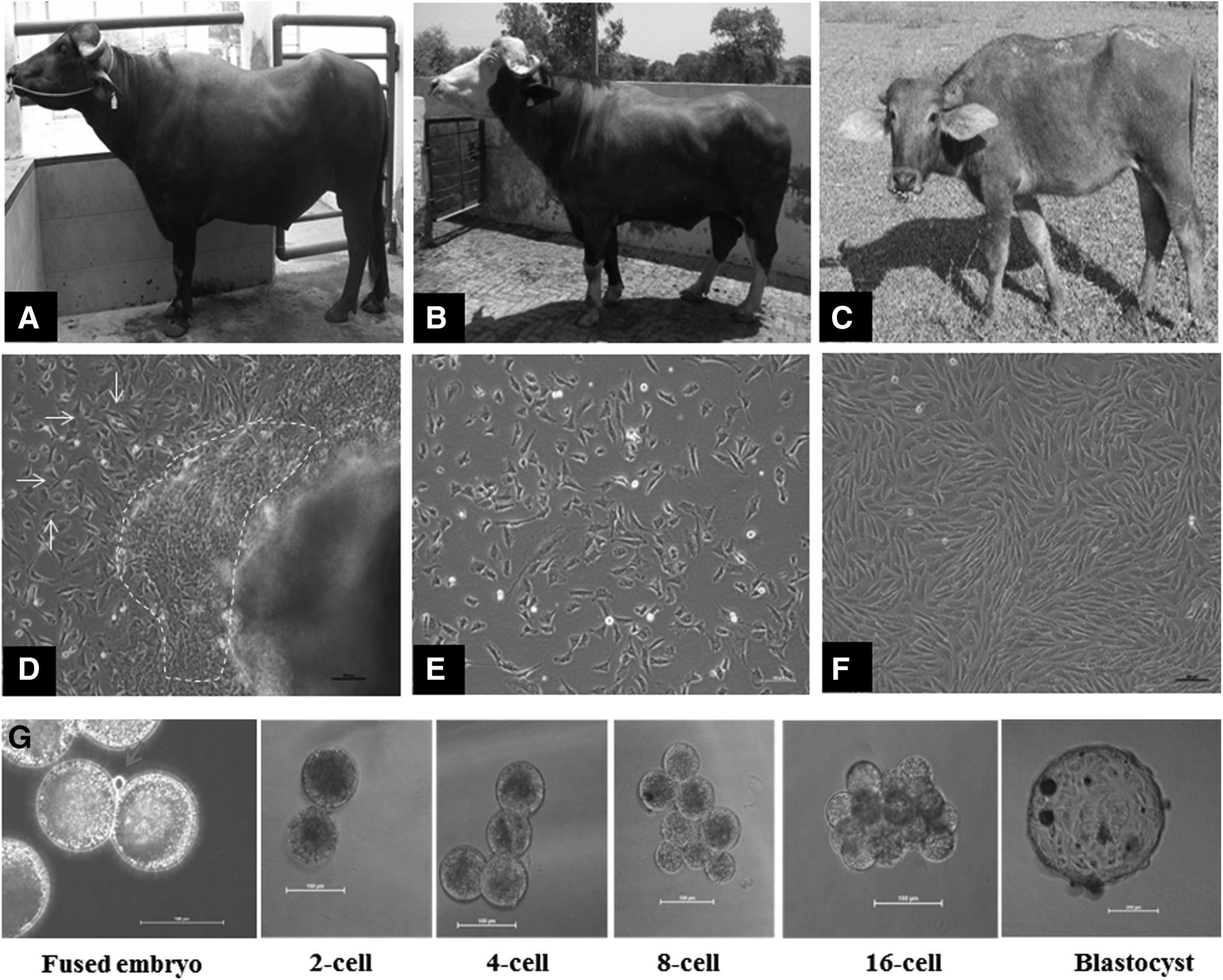
Buffalo biopsy donors, derivation of primary buffalo somatic cells, and developmental stages of buffalo cloned embryos: The buffalo breeds selected for isolation of somatic cells—
Successfully fused embryos were incubated in the TCM medium supplemented with 20% FBS for 6 hours in a CO2 incubator. After 6 hours, embryos were activated by incubating in 2 μM of calcium ionophore (A23187, Sigma) for 5 minutes, followed by 4-hour incubation in 2 mM 6-dimethylamino purine in a CO2 incubator. Activated embryos were cultured in 400 μL of Research Vitro Cleave medium (K-RVCL-50, Cook®, Brisbane, Qld, Australia) supplemented with 1% fatty acid-free bovine albumin in four-well dishes (15–20 embryos per well of four-well dishes) for 8 days into a CO2 incubator at 38.5°C. The cleavage and blastocyst rates were recorded on day 8 of in vitro culture. We used six blastocysts each of riverine and swamp-like buffaloes, and were stained with H-33342 DNA stain to determine total cell number per blastocyst.
Statistical analyses
All statistical analyses were performed using the SAS/STAT® software (SAS Version 9.3; SAS Institute, Inc., Cary, NC, 2011). The cleavage and blastocyst rates were compared using the Glimmix procedure (PROC GLIMMIX) of SAS. The total cell number was analyzed using general linear model (PROC GLM) of SAS. Differences were defined as p < 0.05 (p-values less than 0.05 were considered significant).
Results and Discussion
In this study, we isolated and cryopreserved primary fibroblasts from 14 riverine and swamp-like buffaloes of three breeds for the initiation of a somatic cell bank (Table 1). Preserved cells belonged to the fibroblast cell type, which was determined by the expression of cytoskeleton marker, namely vimentin. The microsatellite alleles (n = 13) of all animal cells were analyzed and documented, which can be used to determine the unambiguous parentage of born clones in the future. For the handmade SCNT experiments, frozen cells were thawed and cultured. The cloned blastocyst production rates of 34%–42% for riverine buffalo cells and of 21%–26% for swamp-like buffalo cells were achieved. We demonstrated the feasibility of biobanking of buffalo somatic cells.
BuMuCell, BuNRCell, and BuSwCell represent the somatic cells of Murrah, Nili-Ravi, and Swamp-like breeds of buffalo, respectively.
Indicated that all animal cells belonged to fibroblast type, which was characterized using a positive expression of fibroblast marker (namely vimentin) and absence of epithelial cell marker (namely cytokeratin-18), and profile of 13 microsatellite alleles of all animal cell is available for parentage of clones in future.
Indicated that these cells derived from the ear tissue biopsies, while other animal cells derived from tail tissue biopsies.
Indicate an age of animal informed by their owners.
SCNT, somatic cell nuclear transfer.
Biopsies of skin tissue have been commonly used to establish the somatic cell cultures of domestic animals. Collection of skin tissue biopsies is simple, rapid, and relatively painless. In domestic animals, at the time of identification, ear tags can be collected without any extra efforts, which can be used to establish cultures of somatic cells (Groeneveld et al., 2008). In this study, we collected skin biopsies from the underneath part of the tail, just above the anal region, which has the least exposure to sunlight, since constant sunlight exposure has been reported to cause mutations of cellular DNA (Hoffmann-Dörr et al., 2005; Schuch and Menck, 2010).
We assume that cells derived from tissues that are least exposed to sunlight may be efficient in the genomic reprogramming during SCNT, compared with cells derived from sunlight-exposed ear-tags or skin tissues. The impact of exposure to sunlight on the suitability of skin fibroblasts for reprogramming needs further investigations, which are beyond the scope of this work.
In culture, outgrowths of the two types of cells, fibroblast-like and epithelial-like, were observed after 7–10 days of explant seeding. The fibroblast-like cells speedily proliferated, while the epithelial-like cells slowly grew close to the vicinity of explants (Fig. 1D). We are interested in the culturing of fibroblast cells because the fibroblast cell type is robust and quickly proliferates under basic culture conditions, while epithelial cells require specific nutrient factors and matrix supports, which may increase the cost of cell maintenance in vitro (Freshney, 2010).
Cultures of primary explants were treated with 0.25% trypsin-EDTA solution for 2–3 minutes, which resulted in early dissociation of fibroblast cells as they were loosely attached to the surface of culture dishes in comparison to epithelial cells, which helps in purification of fibroblast cells (Freshney, 2010). The purity of fibroblast cells was confirmed by positive expression of vimentin, a fibroblast cell-type marker, and no expression of cytokeratin-18, an epithelial cell-type marker (Fig. 2).
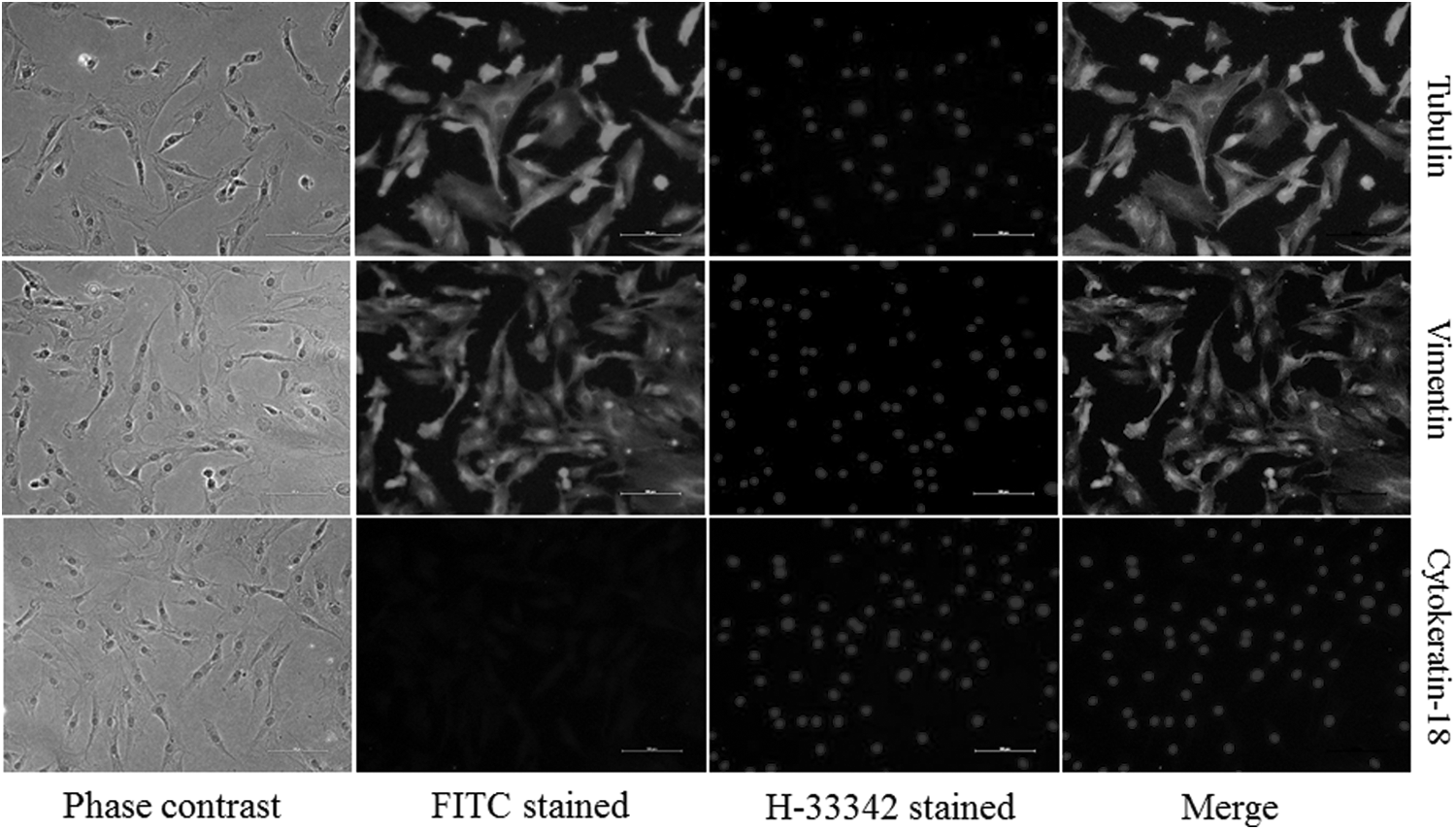
Exemplary immunofluorescence staining of buffalo somatic cells. Phase contrast, FITC labeled and H-33342 labeled images of cells stained with cytoskeleton markers, such as tubulin, vimentin, cytokeratin-18. Positive signals for vimentin and no signals for cytokeratin-18 indicate that cultured cells belonged to fibroblast cell type. Tubulin staining was used as positive staining controls. Scale bar represents 100 μm. FITC, fluorescein isothiocyanate.
Cross-contamination of cell lines can happen when a large number of cell cultures are simultaneously handled and propagated (Freshney, 2010). Single somatic cells are used as donors to produce cloned animals, and the parentage of clones has to be confirmed by matching of genotype between donors and clones (Arat et al., 2011; Selokar et al., 2014). Hence, profiling of the genotype of all somatic cells preserved in biobanks needs to be done, preferably, before upscaling and freezing. Such genotyping database may save time and investment required to screen the produced clones if there is an unexpected genotyping mismatch between donors and clones. We analyzed 13 microsatellite alleles in all somatic cells (Supplementary Table S1), which allowed an unequivocal identification of their genotypes.
The effectiveness of somatic cell biobanks greatly depends on the use of the preserved cells to resurrect living animals using animal cloning techniques (Arat et al., 2011). In the present study, we used cells of seven animals as donors in the SCNT to determine their ability to support in vitro developmental competence of cloned embryos. The optimized handmade cloning method was used to produce cloned embryos (Selokar et al., 2011), and the developmental stages of the cloned embryos of buffaloes are presented in Figure 1G. Since the Zona pellucida is removed for the handmade cloning, during the in vitro culture, the initial blastomeres of two-cell to morulae stages are unordered, however, at day 8, apparently normal blastocyst compositions are found.
The handmade cloning method has the advantage that no specific micromanipulation equipment is required; instead a standard stereo-zoom microscope and a microblade are sufficient. We found cloned blastocyst production rates of 34%–42% for riverine buffalo cells and rates of 21%–26% for swamp-like buffalo cells (Table 2). We pooled data of cleavage and blastocyst rates of riverine and swamp-like SCNT experiments for statistical analysis. The cleavage rates (95.99% ± 2.17% vs. 82.18% ± 2.50%) and blastocyst rates (37.73% ± 1.54% vs. 24.31% ± 1.78%) were higher (p < 0.05) for riverine buffalo cells than that of swamp-like buffalo cells (Fig. 3), whereas the total cell number of blastocysts (258.16 ± 36.25 and 198.16 ± 36.25, respectively) were similar.
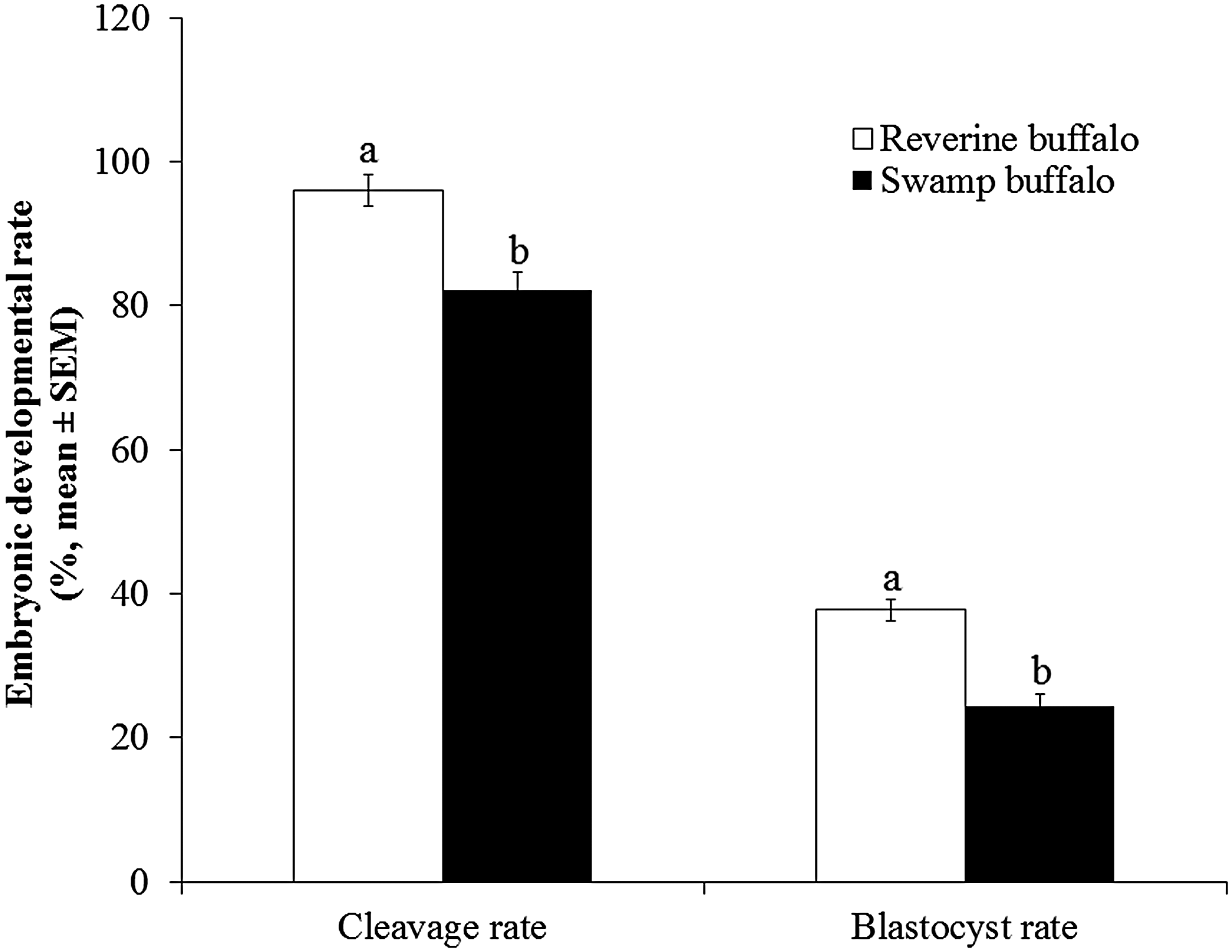
Overall cleavage and blastocyst rates of cloned embryos of riverine and swamp-like buffaloes. Means with different letters are significantly different (t-test, p < 0.05).
The cloned embryo production rates of riverine buffalo were similar to previous studies in buffalo (Madheshiya et al., 2015; Mahapatra et al., 2015; Saini et al., 2016; Selokar et al., 2011). The SCNT results of swamp-like buffalo cells indicate that riverine buffalo oocytes have capabilities to reprogram the cells of swamp-like buffalo; however, lower cleavage and blastocyst rates might be associated with incomplete nuclear reprogramming or mitochondrial incompatibility between recipient oocytes and donor cells, which requires further investigation. Overall, SCNT results indicate that cell freezing does not influence the ability of cells to be used as donors for the in vitro production of cloned embryos. Somatic cell biobanks could be used to conserve the population of endangered livestock using SCNT; however, more attempts are required to improve the efficiency of SCNT.
In conclusion, we cryopreserved somatic cells of 14 buffaloes in liquid nitrogen for long-term storage. The preserved cells belonged to fibroblast cell type. The SCNT results indicate that cell freezing does not affect in vitro production of cloned embryo. We are continuing to work on the preservation of more somatic cells of buffalo, with the same approaches and objectives as mentioned during this study. This study encourages the development of somatic cell biobanks of domestic livestock, including the endangered buffalo breeds.
Footnotes
Acknowledgments
This work was financed by the Indian Council of Agricultural Research, Delhi, India. We are thankful to the Director of ICAR-Central Institute for Research on Buffaloes to provide necessary support for this study. We are greatly thankful to buffalo owners for allowing us to collect skin biopsies from their buffaloes. We are grateful to Dr. Gopal Das, College of Veterinary Science, Guwahati, India for assisting in the collection of tissue samples from swamp-like buffaloes. We thank Dr. Wilfried Kues for helpful discussion, comments, and article drafting. We thank Juan Carlos Gutiérrez-Añez for statistical analysis of data.
Authors' Contributions
Identification of animals and design of the study: P.S.Y. and N.L.S. Culture, characterization, and cryopreservation of somatic cells of riverine buffaloes available in institute herds: N.L.S., P.S., D.K., M.S., A.S. and K.V.; Culture, characterization, and cryopreservation of somatic cells of riverine buffaloes belonged to farmers: N.L.S. and M.S.; Culture, characterization, and cryopreservation of somatic cells of swamp-like buffaloes: N.L.S. and A.K. Production of cloned embryos: N.L.S., P.S., and A.K. Reagents/materials/analysis tools contributed: P.S.Y., N.L.S., and D.K. Article writing: N.L.S.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.