
Abstract
Abstract
Epigenetic reprogramming and somatic cell nuclear transfer (SCNT) cloning efficiency were recently enhanced using histone deacetylase inhibitors (HDACis). In this study, we investigated the time effect of CI994, an HDACi, on the blastocyst formation rate, acetylation levels of H3K9 and H4K12, DNA methylation levels of anti-5-methylcytosine (5mC), and some mRNA expression of pluripotency-related genes in pig SCNT embryos. Treatment with 10 μM CI994 for 24 hours significantly improved the blastocyst formation rate of SCNT embryos in comparison with the untreated group (p < 0.05). Moreover, average fluorescence intensities of H3K9 and H4K12 in CI994-treated embryos were remarkably increased at the pseudo-pronuclear stage, but not at the blastocyst stage. The intensity of POU5F1 was higher in CI994-treated blastocysts than in control blastocysts, whereas that of 5mC did not differ between the two groups. The percentage of apoptotic cells in blastocysts was significantly higher in the untreated group than in the CI994-treated group. mRNA levels of POU5F1 and SOX2 were significantly increased in the CI994-treated group. These observations suggest that optimum exposure (10 μM for 24 hours) to CI994 after activation elevates the level of histone acetylation and subsequently improves the in vitro development of pig SCNT embryos.
Introduction
S
Although cloned pigs have been successfully produced, the survival rate to birth for cloned embryos remains unsatisfactorily low. This low efficiency is assumed to be due to incomplete or incorrect reprogramming, such as abnormal histone modifications and DNA methylation (Cao et al., 2017; Ding et al., 2008; Kang et al., 2001). Hence, repair of aberrant nuclear reprogramming may increase the rate of development into healthy cloned offspring.
A variety of small chemical compounds are used to aid nuclear reprogramming and improve the efficiency of nuclear transfer (NT), such as the DNA methylation inhibitors, 5-aza-2′-deoxycytidine and RG108; the histone lysine methyltransferase inhibitor, BIX-01294; and the histone deacetylase inhibitors (HDACis), valproic acid (Kang et al., 2013), sodium butyrate (Xiong et al., 2015), CUDC-101 (Jin et al., 2014), m-carboxycinnamic acid bishydroxamide (Song et al., 2014), suberoylanilide hydroxamic acid (Whitworth et al., 2015), PCI-24781 (Jin et al., 2016), PXD101 (Qiu et al., 2017), trichostatin A (Cao et al., 2017), LAQ824 (Jin et al., 2017a), scriptaid (Zhang et al., 2016), and M344 (Jin et al., 2017b).
CI994 (tacedinaline, N-acetyldinaline [4-(acetylamino)-N-(2-amino-phenyl) benzamide]), a class I HDACi, is used as a cytostatic anticancer agent (Monneret, 2005; Pauer et al., 2004). CI994 has been applied in many settings; however, the effect of CI994 treatment on the developmental capacity of cloned pig embryos has not been reported. Hence, we hypothesized that CI994 treatment could improve the nuclear reprogramming of cloned pig embryos by increasing histone acetylation levels.
To validate this assumption, we explored the effect of CI994 on the developmental competence of cloned porcine embryos and to investigate its effect on acetylation levels of histone H3 lysine 9 (H3K9ac) and histone H4 at lysine 4 (H4K12ac), the global DNA methylation level of 5-methylcytosine (5mC), and the protein level of POU5F1 using immunofluorescence. We also compared apoptosis and expression of the pluripotency-related genes, POU5F1, NANOG, and SOX2, between CI994-treated and untreated SCNT blastocysts.
Materials and Methods
Animals
This study was carried out in strict accordance with the Guide for the Care and Use of Laboratory Animals of Yanbian University. Animal procedures were conducted following the protocol approved by the Animal Care and Welfare Committee of Yanbian University.
Chemicals
CI994 was purchased from Selleck Chemicals (Houston, TX). Other chemicals and reagents used in this study were purchased from Sigma-Aldrich Chemical Company (St. Louis, MO) unless otherwise stated.
Production of pig SCNT embryos and embryo transfer
Hybrid pig ovaries were collected at a local slaughterhouse and transported to the laboratory at 31°C–35°C. Cumulus–oocyte complexes (COCs) surrounded by a minimum of three cumulus cells were selected and cultured in maturation medium (NCSU-37). Following 22 hours of maturation with hormones, the COCs were transferred in hormone-free medium for another 18–24 hours. The detailed information was previously published (Jin et al., 2017c).
Ear fibroblasts were isolated from the tissue of an adult pig. Tissues were cut into pieces and cultured at 38°C in 5% CO2 in Dulbecco's modified Eagle's medium (DMEM; HyClone). The detailed procedures were previously published (Kang et al., 2017).
SCNT procedure was performed as previously described (Yin et al., 2002). In brief, matured oocytes were enucleated by aspirating the first polar body. A single donor cell was placed into the perivitelline space of an enucleated oocyte and electrically fused using two direct pulses of 150 V/mm for 50 μs in fusion solution (0.28 M mannitol supplemented with 0.1 mM MgSO4).
Fused couplets were activated by two direct pulses of 100 V/mm for 20 μs in activation solution (0.28 M mannitol containing 0.1 mM MgSO4 and 0.05 mM CaCl2). Then, eggs were cultured in NCSU-37 medium under 5% CO2, 5% O2, and 90% N2 at 38°C for 7 days. To count the total number of cells in blastocysts, day 7 SCNT blastocysts were washed in phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde in PBS, and stained with 20 μg/mL propidium iodide (PI). The number of nuclei was counted under an epifluorescence microscope (Nikon, Tokyo, Japan) equipped with a digital camera.
Embryo transfer (ET) was conducted during autumn and winter. For ET, ∼150 SCNT embryos per recipient were kept in manipulation medium, transported in a portable incubator, and loaded into a sterilized straw before transfer. CI994-treated (10 μM for 24 hours) or nontreated pig SCNT embryos at the one-cell stage after fusion or at the two- to four-cell stage after a day of culture were transferred into the oviducts of naturally cycling gilts on the first day of standing estrus. Cloned fetuses were recovered 21 days after the transfer was performed.
Immunofluorescence
Untreated and CI994-treated cloned embryos were collected at the pseudo-pronuclear and blastocyst stages. The immunofluorescence protocol was adapted from a previous study (Liang et al., 2015). Briefly, fixed and permeabilized embryos were blocked in 2% bovine serum albumin prepared in PBS. This was followed by incubation in blocking solution together with primary antibodies at 4°C overnight. The post-translational modifications were investigated on H3K9 and H4K12. The primary antibodies were rabbit polyclonal antibodies against H3K9 and H4K12 (diluted 1:200, 07-352 and 07-595; Upstate Biotechnology, NY) or a rabbit polyclonal antibody against amino acids 1–134 mapping at the N-terminus of Oct-3/4 of human origin (diluted 1:200, sc-9081; Santa Cruz Biotechnology, CA).
After washing with PBS, embryos were incubated with a goat anti-rabbit fluorescein isothiocyanate (FITC)-conjugated secondary antibody (diluted 1:200, F0382; Sigma, MO) for 1 hour at 38°C. For staining of global methylation, permeabilized embryos were treated with 4 N HCl for 30 minutes at room temperature and neutralized for 10 minutes with 100 mM Tris-HCl buffer. After washing three times, samples were incubated with primary antibodies, a mouse monoclonal antibody against 5mC (diluted 1:200, mAb, NA81; Calbiochem, Damstadt, Germany), and secondary antibodies, an FITC-conjugated goat anti-mouse secondary antibody (diluted 1:200, F2012; Sigma).
DNA was counterstained with 20 μg/mL PI for 10 minutes. After a final wash in PBS, embryos were mounted on glass slides in a drop of 100% glycerol, compressed gently with a cover slip, and scanned using an epifluorescence microscope (Nikon). Images were captured and quantified using Nikon NIS element software.
Apoptosis assays
TUNEL procedures have been previously published (Jin et al., 2016). Fixed and permeabilized SCNT embryos were labeled with a fluorescein-conjugated terminal deoxynucleotidyl transferase 2′-deoxyuridine 5′-triphosphate nick end labeling (TUNEL) assay kit (In Situ Cell Death Detection Kit, Roche Diagnostics Deutschland GmbH, Mannheim, Germany) in the dark for 1 hour at 38°C. Cloned blastocysts were stained with 25 μg/mL Hoechst 33342 to determine the total cell number. Samples were mounted on glass slides with a drop of antifade mounting medium and analyzed using an epifluorescence microscope (Nikon) equipped with a digital camera.
Quantitative real-time polymerase chain reaction
To investigate the mRNA expression levels of pluripotency-related genes (POU5F1, NANOG, and SOX2), blastocysts derived from CI994-treated (10 μM for 24 hours) and untreated porcine NT embryos were collected. Quantitative real-time polymerase chain reaction (PCR) was performed according to a previous report (Zhu et al., 2017). Total RNA was extracted from at least 35 blastocysts per condition using a Dynabeads mRNA DIRECT Kit (Life Technologies AS, Oslo, Norway) and the RNA concentration was determined using a UV-vis spectrophotometer (UV-2450; Shimadzu, Tokyo, Japan). Complementary DNA was immediately synthesized from extracted RNA using the SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA). Real-time PCR amplification was conducted with a QuantiTect SYBR Green PCR kit (TaKaRa, Kusatsu, Japan) on an Mx3005P real-time PCR system (Agilent Genomics).
Each real-time PCR mixture contained 1 μL of cDNA (1400 ng/μL), 10 μL of SYBR, 0.5 μL of ROX, 7.5 μL of nuclease-free water, and 0.5 μL of appropriate forward and reverse primers (Table 1) in a total volume of 20 μL. The expression of each target gene was quantified relative to that of the internal reaction control gene [glyceraldehyde 3-phosphate dehydrogenase (GAPDH)] using the equation R = 2-[ΔCt sample-ΔCt control]. For ease of comparison, the average mRNA transcript level of each gene in the control group was set to 1.
Statistical analysis
Each experiment was repeated at least three times. Data are expressed as mean values ± standard deviations. Data were analyzed using a univariate analysis of variance (ANOVA), followed by Duncan's multiple range test using SPSS 16.0 (SPSS, Inc., Chicago, IL) statistical software. The average fluorescence intensity of each individual nucleus was quantified using Nikon NIS element software. Gene expression and SCNT blastocyst formation rates were compared using Student's t-test. p < 0.05 was regarded as statistically significant.
Results
Effect of CI994 treatment on the in vitro and in vivo developmental competence of pig SCNT embryos
After activation, reconstructed embryos were treated with different concentrations (0, 1, 10, and 100 μM) of CI994 for 24 hours and their in vitro developmental competence was examined. The blastocyst formation rate of embryos treated with 10 μM CI994 was significantly higher than those of embryos treated with 1 and 100 μM CI994 and nontreated embryos (10 μM, 23.3% ± 5.9%; 1 μM, 12.2% ± 3.9%; 100 μM, 13.0% ± 5.9%; and nontreated, 10.6% ± 1.2%; Table 2). However, CI994 treatment had no effect on the cleavage rate or total cell number per blastocyst (Fig. 1B, D).

Effect of treatment with CI994 on the development of pig SCNT embryos to the blastocyst stage.
Values with different superscripts in the same column are significantly different (p < 0.05).
Based on these results, 10 μM was chosen as the optimal concentration of CI994. Next, the effects of treatment with 10 μM CI994 for different durations (0, 12, 24, and 48 hours) on development of porcine SCNT embryos were analyzed. The proportion of reconstructed embryos that developed to the blastocyst stage was significantly higher in the group treated with CI994 for 24 hours than in the control group (21.9% ± 6.4% vs. 11.4% ± 0.5%; Table 3). In addition, the percentage of embryos that developed to the blastocyst stage did not significantly differ among groups treated with CI994 for 12, 24, and 48 hours. However, CI994 treatment did not affect the cleavage rate and mean number of cells per blastocyst. Hence, these data suggest that treatment with 10 μM CI994 for 24 hours can remarkably improve the in vitro developmental capacity of cloned pig embryos (Fig. 1A, C).
Values with different superscripts in the same column are significantly different (p < 0.05).
To investigate the effect of CI994 treatment on the in vivo developmental capacity, NT embryos were transferred into surrogates. An average of 156 untreated embryos were transferred into three surrogate mothers (145, 154, and 169 embryos to individual recipients) and two fetuses were harvested. Conversely, averages of 145 CI994-treated cloned embryos were transferred into three surrogates (138, 151, and 146 embryos into individual recipients); one recipient became pregnant, and three fetuses were obtained.
Effect of CI994 treatment on levels of H3K9ac and H4K12ac in cloned pig embryos
Immunofluorescence signals for H3K9ac and H4K12ac were significantly increased in the CI994-treated group at the pseudo-pronuclear stage (Fig. 2). In addition, the intensity of H3K9ac labeling was twofold lower than that of H4K12ac in control embryos at the pseudo-pronuclear stage. Additionally, the level of H4K12ac was significantly higher than that of H3K9ac in CI994-treated porcine SCNT embryos at the pseudo-pronuclear stage. However, acetylation levels did not clearly differ between the CI994-treated and control groups at the blastocyst stage (Fig. 3).
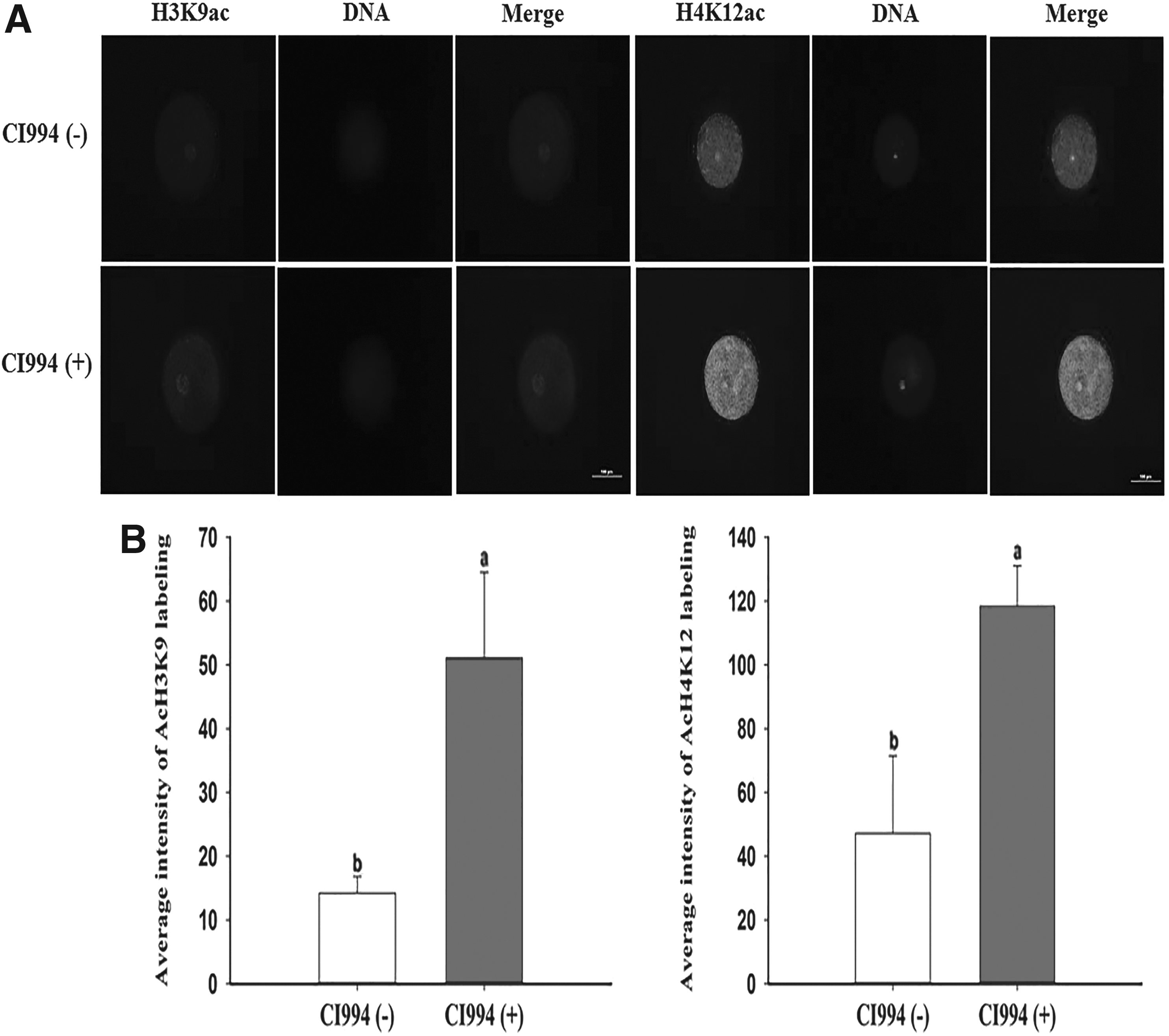
Evaluation of H3K9ac and H4K12ac levels in pseudo-pronuclear-stage embryos.
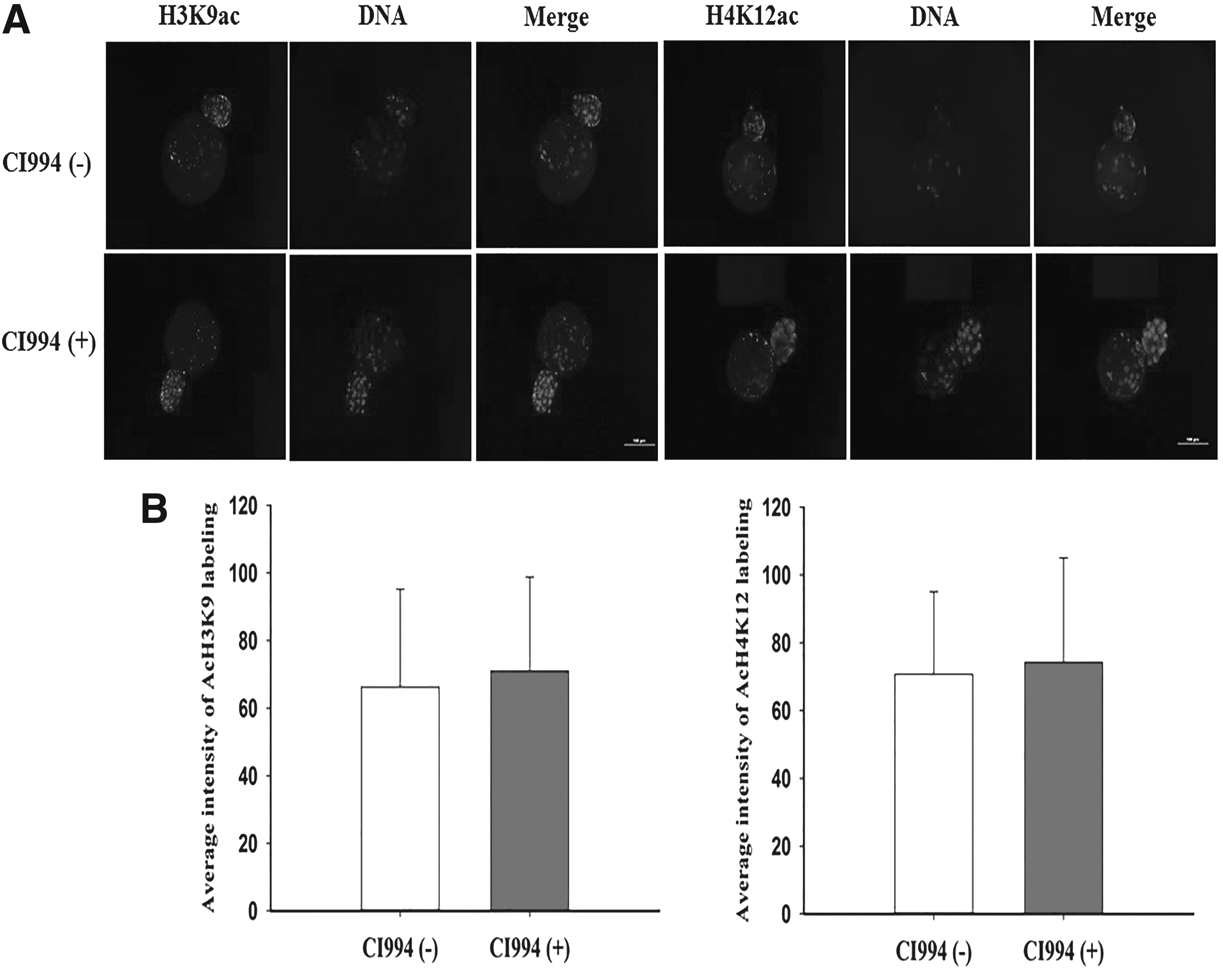
Evaluation of H3K9ac and H4K12ac levels in blastocyst-stage embryos.
Effect of CI994 treatment on 5mC and POU5F1 expression in SCNT blastocysts
We examined the intensity of 5mC and POU5F1 labeling in day 7 pig NT blastocysts. The immunofluorescence staining intensity of 5mC was similar in CI994-treated and untreated embryos. However, treatment with 10 μM CI994 for 24 hours remarkably increased the intensity of POU5F1 staining in cloned embryos compared with untreated embryos (Fig. 4).
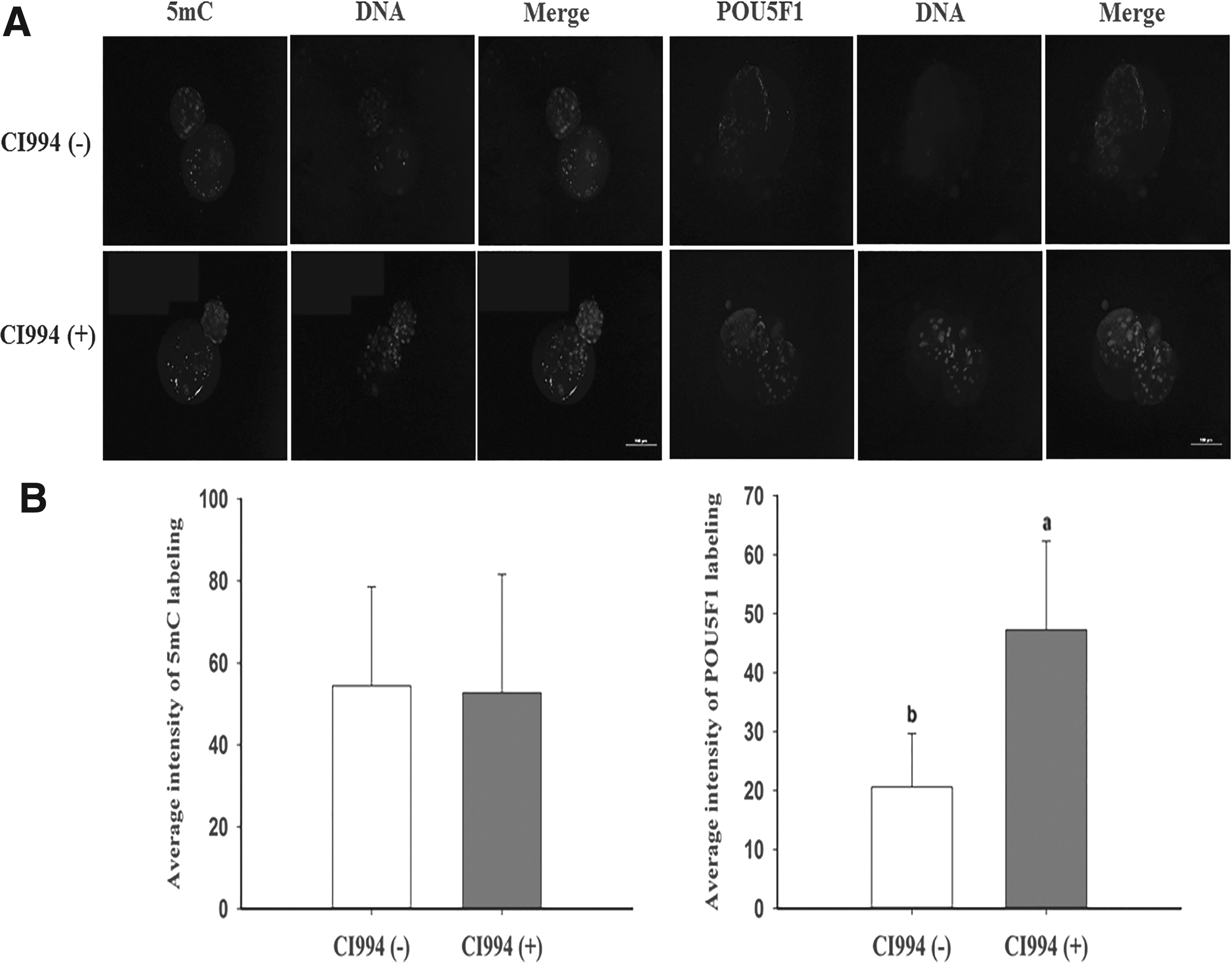
Immunofluorescence staining of 5mC and POU5F1 in porcine SCNT embryos at the blastocyst stage.
Effect of CI994 treatment on apoptosis in cloned blastocysts
To explore the effect of CI994 treatment on the pig SCNT blastocyst quality, we examined the number of apoptotic cells by the TUNEL assay. The percentage of apoptotic cells in cloned blastocysts was significantly higher in the control group than in the CI994-treated group (Fig. 5).
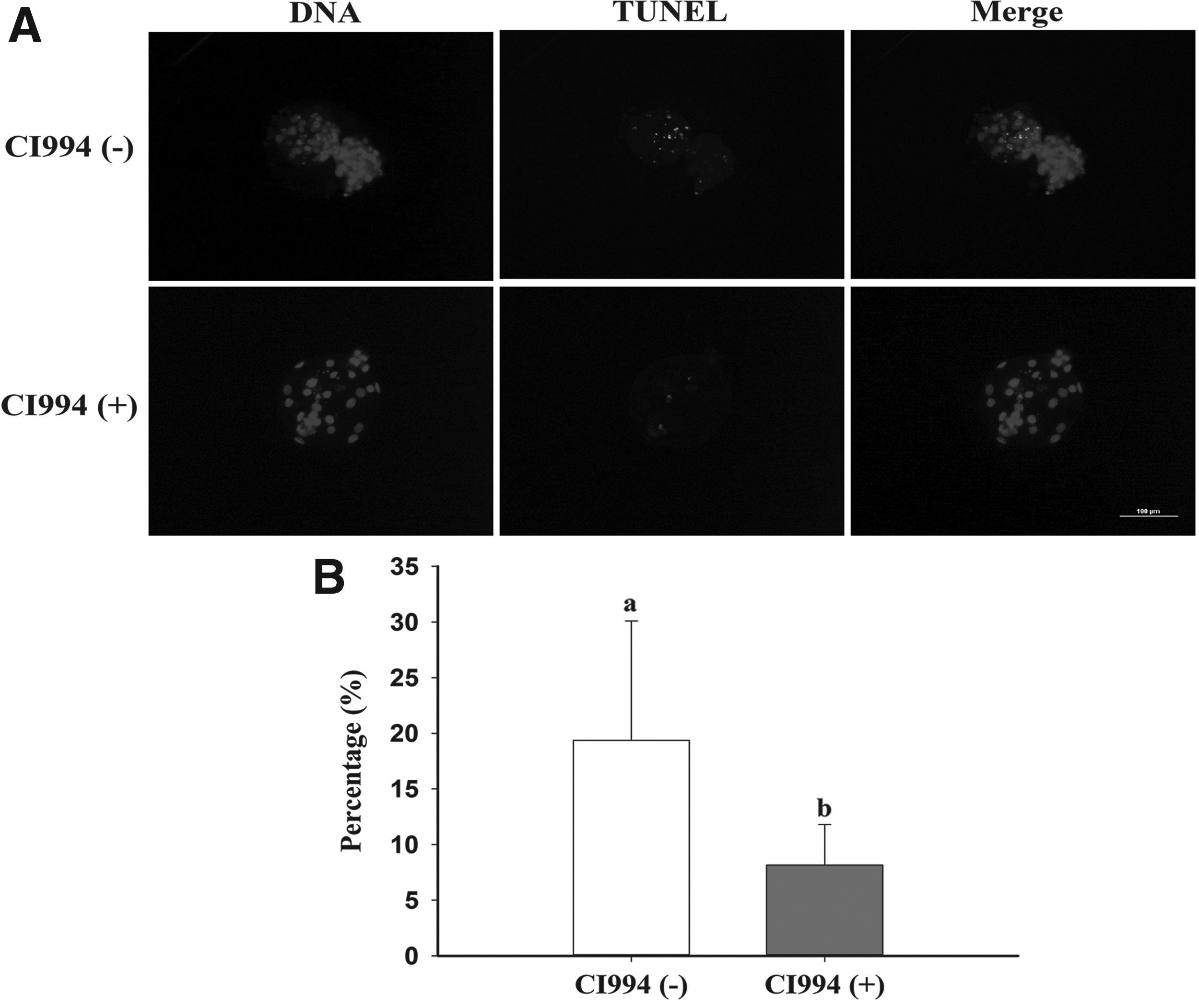
Representative images of apoptotic cells in porcine SCNT blastocysts.
Gene expression levels in pig SCNT blastocysts
To evaluate the quality of blastocysts, we measured POU5F1, NANOG, and SOX2 mRNA expression in CI994-treated SCNT blastocysts (Fig. 6). mRNA expression levels of POU5F1 and SOX2 were much higher in the CI994-treated group than in the control group. However, NANOG mRNA expression did not significantly differ between the CI994-treated and untreated groups.
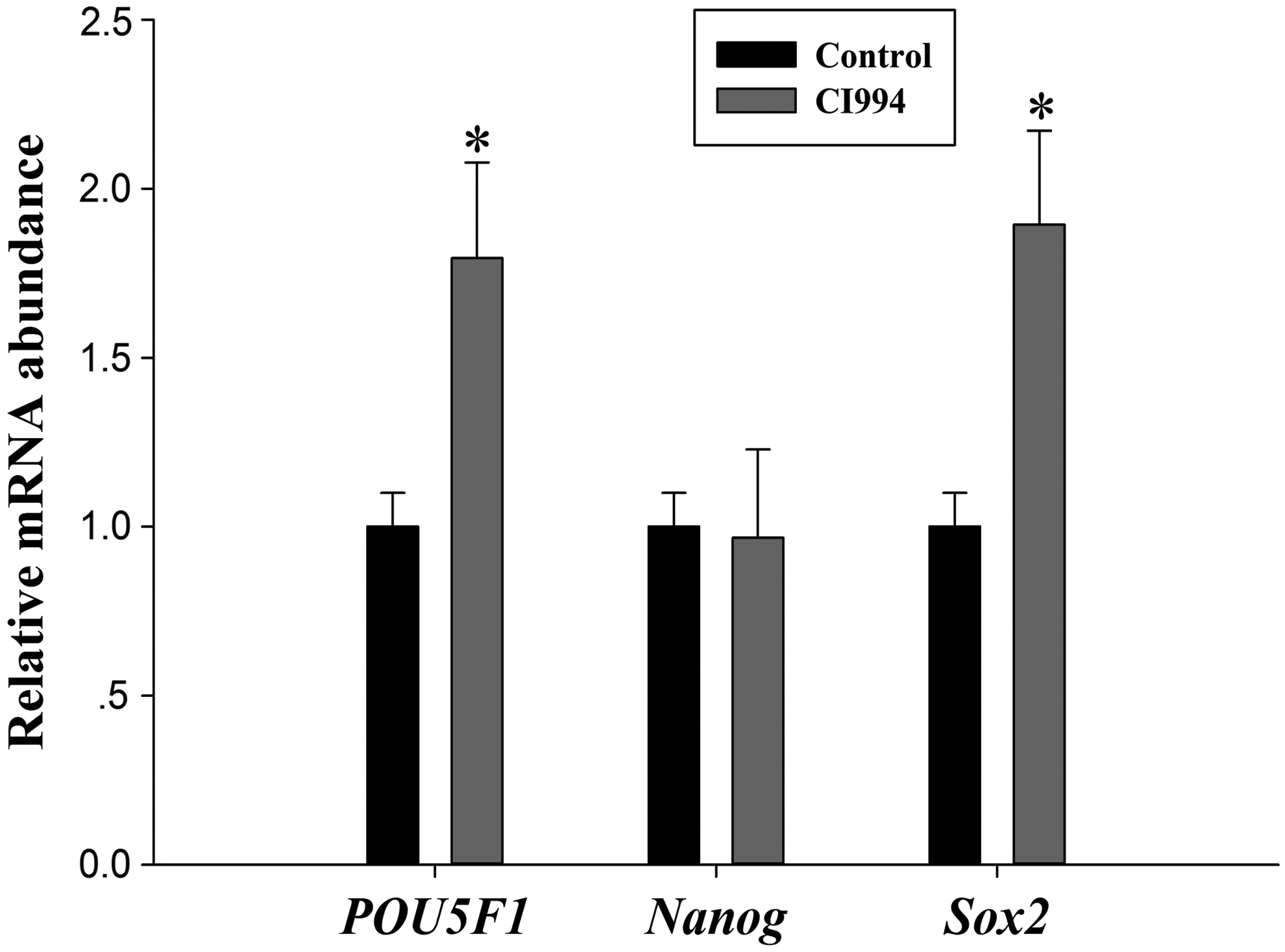
mRNA expression levels (mean ± SEM) of POU5F1, NANOG, and SOX2 in porcine SCNT blastocysts after 7 days of in vitro culture. Embryos were untreated or treated with CI994; *indicates p < 0.05 compared with the control group.
Discussion
Recently, several lines of investigation revealed abnormal DNA methylation, histone modifications, and gene expression in reconstructed embryos (Jin et al., 2017b; Liang et al., 2015). Several studies suggested that HDACis improve the efficiency of SCNT (d; Jin et al., 2017c; Sangalli et al., 2014); however, the precise molecular mechanism by which they act has not been fully elucidated.
Eighteen mammalian HDACs fall into four classes on the basis of their homology to yeast proteins (Verdone et al., 2006). Generally, hyperacetylation induced by inhibition of HDACs loosens the association between nucleosomes and DNA or linker histones and relaxes the chromatin structure (Lee et al., 1993). CI994 is a potent member of the benzamide class of HDACis, which inhibits HDAC1 with an IC50 of 0.57 μM and causes G1 cell cycle arrest (Methot et al., 2008). Recently, Seki et al. (2016) demonstrated that CI994 can reduce the total time of fibrillation, atrial fibrosis, and intra-atrial adipocyte and immune cell infiltration in sustained atrial fibrillation, without significant effects on cardiac function.
The ability of CI994 to enhance nuclear reprogramming and improve the efficiency of SCNT has not been reported. Our results showed that treatment with 10 μM CI994 for 24 hours remarkably improved the in vitro developmental potential of cloned pig embryos. This improvement in the development of reconstructed embryos by CI994 treatment may be owing to histone hyperacetylation. In this study, CI994-treated SCNT embryos were transferred into three surrogates and three fetuses were harvested. In accordance with other studies, HDACi treatment did not affect the in vivo development of cloned embryos (c, d; Jin et al., 2017b).
Histone acetylation is a key epigenetic modification that modulates chromatin structure and function, improves nuclear reprogramming, and plays a critical role in the development of cloned embryos. H3K9ac predicts the pluripotency and reprogramming capacity (Hezroni et al., 2011), and its level decreases with embryonic stem cell differentiation (Krejci et al., 2009). A study by van der Heijden et al. (2006) showed that H4K12ac is important for the paternal influence on early gene expression in zygotes. Cumulatively, the increased H3K9ac and H4K12ac levels could have positive effects on the developmental competence of porcine SCNT embryos.
DNA methylation reprogramming is a key factor in mammalian embryogenesis. Accruing evidence shows that following NT, aberrant DNA methylation contributes to the abnormal expression of genes critical for development and to subsequent failed embryogenesis (Dean et al., 2001; Tian et al., 2009; Zhang et al., 2016). Therefore, global genomic methylation levels in cloned embryos were investigated by immunostaining for 5mC. DNA methylation levels in 10 μM CI994-treated blastocysts were similar to those in control blastocysts (Fig. 4), supporting previous studies (Cao et al., 2017; Jin et al., 2017a). However, this observation is inconsistent with several recent findings that scriptaid treatment decreases the intensity of 5mC labeling in bovine and porcine SCNT embryos (Liang et al., 2015; Zhang et al., 2014).
Apoptosis, an important criterion for evaluation of blastocyst quality, occurs frequently during early embryonic development and has a marked impact on embryo development (Haouzi and Hamamah, 2009). In the current study, CI994 treatment significantly inhibited apoptosis in cloned blastocysts. This result implies that CI994 improves the quality of cloned pig embryos by reducing cell death, which is consistent with previous studies (Jin et al., 2016; Liang et al., 2015).
POU5F1 (also known as Oct3/4), a core transcription factor, plays an important role in determining the early steps of embryogenesis and differentiation (Rossant and Tam, 2009). POU5F1 is expressed in all cells of porcine blastocyst-stage embryos before they reach the uterus, but its expression is downregulated in both trophectoderm and primitive endoderm upon arrival in the uterus (Keefer et al., 2007). POU5F1 is therefore considered an ideal marker for evaluating nuclear reprogramming in cloned pig embryos. The mRNA and protein levels of POU5F1 were significantly higher in CI994-treated embryos than in control embryos.
Similar to previous studies, expression of POU5F1 was increased by HDACi treatment (Liang et al., 2015; Sangalli et al., 2014; Song et al., 2014; Xiong et al., 2015). NANOG is an NK-2 class homeobox transcription factor that is expressed throughout pluripotent cells in the ICM (Chambers et al., 2003; Mitsui et al., 2003). NANOG expression did not differ between CI994-treated and untreated embryos. This observation is inconsistent with previous studies (Jin et al., 2017a; Xu et al., 2013). SOX2, a member of the SoxB1 transcription factor family, plays an essential role in different types of multipotent stem cells (Adachi et al., 2013) and induction of pluripotency (Takahashi and Yamanaka, 2006). In this study, mRNA expression of SOX2 was enhanced by CI994 treatment at the blastocyst stage. This observation is in line with previous studies (Jin et al., 2017a; Su et al., 2011).
Conclusion
In summary, the present study indicates that CI994 treatment (10 μM for 24 hours) is sufficient to improve the in vitro early developmental efficiency of porcine SCNT embryos. Improvement in the blastocyst formation rate could be related to enhanced H3K9ac and H4K12ac, decreased cell death in cloned blastocysts, and upregulated mRNA expression of pluripotency genes. We also examined the in vivo development of CI994-treated SCNT embryos and obtained three fetuses. Further studies are required to further elucidate the mechanism by which CI994 improves cloning efficiency.
Footnotes
Acknowledgments
This study was supported by the Science and Technology Development Projects of Jilin Province of China (grant number: 20160520057JH), the 13th 5-Year Plan Science and Technology Research Project of the Education Department of Jilin Province of China (grant number: 2016-257), and the Institute for Basic Science (grant number: IBS-R021-D1-2016-a06). The authors would like to thank Hanji and Longxing pig farms for providing surrogate pig recipients.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.