
Abstract
Abstract
Wedelolactone is a multitarget natural plant compound with many pharmacological activities, including anti-inflammatory, anticancer, and antiosteoporosis. In this study, dental pulp stem cells (DPSCs) were treated with or without wedelolactone. We found that wedelolactone stimulated odontoblast differentiation and mineralization. At the molecular level, wedelolactone directly promoted the nuclear accumulation of β-catenin, and thereafter stimulated the expression of odontoblast-related marker genes containing dentin matrix protein-1 (DMP1), dentin sialophosphoprotein (DSPP), and runt-related transcription factor 2 (Runx2). Furthermore, wedelolactone upregulated the expression of IκBα and inhibited phosphonation and nuclear migration of p65. As a result, wedelolactone remarkably induced odontoblast differentiation through semaphorin 3A (Sema3A)/neuropilin-1 (NRP1) pathway-mediated β-catenin activation and nuclear factor kappa B (NF-κB) pathway inhibition. Our findings provide novel perceptions on odontogenic differentiation of DPSCs.
Introduction
D
Wedelolactone is a natural small molecular derived from Eclipta prostrata and Wedelia chinensis, which is a traditional Chinese food additive and herbal medicine to treat osteoporosis, hepatic disease, renal injuries, hyperlipidemia, viper bites and hair loss (Liu et al., 2014; Zhao et al., 2015). Wedelolactone has been reported to participate in various biological activities, such as anti-inflammatory, anticancer, and antioxidant activities (Sarveswaran et al., 2012; Syed et al., 2003; Yuan et al., 2013).
Interestingly, growing evidence shows that wedelolactone facilitated osteoblastogenesis through activation of Wnt/β-catenin signaling pathway, which led to the activation of runt-related transcription factor 2 (Runx2) and the expression of downstream genes (Liu et al., 2016). Furthermore, many scholars have found that wedelolactone could inhibit the abnormal proliferation of human renal mesangial cells via regulating the activity of several key members of nuclear factor kappa B (NF-κB) signaling pathway (Shen et al., 2017).
The canonical Wnt/β-catenin pathway plays a crucial role in multiple cellular events, for instance, cell migration, proliferation, and differentiation, especially in developmental and postdevelopmental tooth physiology (Clevers and Nusse, 2012; Liu and Millar, 2010; MacDonald et al., 2009; Nelson and Nusse, 2004). The Wnt family comprises 19 proteins divided into two main categories, namely the β-catenin-dependent canonical and the β-catenin-independent noncanonical Wnt pathways (Maeda et al., 2013). The canonical Wnt pathway transduces its signals via intracellular β-catenin. When cells are in resting state, β-catenin is rapidly degraded by the cytoplasmic complex containing adenomatous polyposis coli (APC) protein, axin, casein kinase I, and glycogen synthase kinase-3β (GSK-3β).
While GSK-3β is inhibited, β-catenin increases in the cytoplasm and translocates to the nucleus, resulting in activating the canonical Wnt/β-catenin pathway (Reya and Clevers, 2005). Canonical Wnt/β-catenin pathway might regulate renovate dentin formation and tooth development (Han et al., 2014; Hunter et al., 2015; Wang et al., 2014). Semaphorin 3A (Sema3A) and its receptor neuropilin-1 (NRP1) are expressed in DPSCs and induce odontoblast differentiation through enhancing canonical Wnt/β-catenin pathway (Song et al., 2017).
Meanwhile, NF-κB plays a crucial role in inflammation and cell survival. The NF-κB superfamily consists of p65 or RelA, p52, p50, c-Rel, and RelB, and generally exists as a homo- or heterodimer in the cytosol, bound to one of three inhibitory IκB subunits. On activation of the complex, the degradation of IκBα releases NF-κB nuclear translocation, where NF-κB binds to specific regulated sequences in the DNA and controls gene transcription (Moynagh, 2005).
Recently, in view of the role of NF-κB, inhibition may contribute to decrease inflammatory cytokines and to enhance odontoblast differentiation and collagen matrix formation in dental pulp stem cells (DPSCs) (Hozhabri et al., 2015). Thus, in the present study, we investigated whether wedelolactone could control odontoblast differentiation of DPSCs and whether the function was associated with the promotion of the Wnt/β-catenin pathway and inhibition of the NF-κB pathway.
Materials and Methods
Cell cultures
All samples were acquired from generally healthy patients (20–28 years of age) after giving the informed consents that were approved by the Ethics Committee of the Affiliated Hospital of Nantong University. Healthy pulp tissues (n = 20) were isolated from the caries-free teeth of patients without oral infection undergoing extraction of fully erupted third molars. We isolated DPSCs from healthy pulp by cleaning the tooth surface, cutting around the cementoenamel junction using sterilized dental fissure burs and then opening to reveal the pulp chamber. The pulp was digested in a solution of 3 mg/mL collagenase type I for 1 hour at 37°C. Single-cell suspensions were obtained by passing the digested tissues through a 70-μm cell strainer (BD Falcon).
Cell suspensions of dental pulp were seeded into 25 cm2 culture dishes and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS; Gibco-BRL, Life Technologies, Inc., Gaithersburg, MD), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C and 5% CO2 (Ledesma-Martinez et al., 2016). The medium was changed every 3 days. Approximately 7–10 days after seeding, the cells became nearly confluent. Cells were passaged at the ratio of 1:3 when they reached 85%–90% confluence. The cell populations were characterized by positive staining with anti-CD34, STRO-1, and c-kit, and the absence of CD45 (Feng et al., 2013). Cells from the third to fifth passages were used in all the experiments.
Odontoblast differentiation
DPSCs (2 × 104 cells/dish) were cultured in 35 mm culture dishes (Costar, Cambridge, MA) in odontogenic differentiation medium containing α-minimum essential medium (α-MEM; Invitrogen, Carlsbad, CA), 15% FBS, 10 mM β-glycerophosphate, 50 mg/mL α-ascorbic acid, 10 nM dexamethasone (Sigma-Aldrich, St Louis, MO), 0.292 mg/mL glutamine, 100 mg/mL streptomycin, and 100 U/mL penicillin G, respectively, for 0, 3, 7, 14, and 21 days, replacing the medium every 2 days. Then, the cells were collected for determination of odontoblast differentiation by evaluating mRNA expression of odontogenic markers, including DMP1 and dentin sialophosphoprotein (DSPP).
The experimental treatment groups consisted of differentiation media with TNF-α (Sigma) and/or wedelolactone (Dalian, China) or BMS-345541 (Sigma) or DKK1 (Santa Cruz). After induced for 0, 3, 7, 14, and 21 days, cells were prepared for alizarin red S staining, alkaline phosphatase (ALP) staining, and immunofluorescence. Protein and RNA were extracted for western blot analysis and real-time polymerase chain reaction (RT-PCR).
Determination of cell number
DPSCs were seeded at 0.7 × 104 cells/well into six-well plates in triplicate for each experimental condition. Wedelolactone (0.1, 1.0, 2.0, and 5.0 μg/mL; Dalian) was added to the cultures or not. DPSCs were collected at 0, 3, 7, 14, and 21 days after plating and dissociated. The total cell numbers were counted and the results expressed as mean ± SEM. Statistical significance was determined using two-way ANOVA followed by post hoc t-test with Bonferroni correction.
Alizarin red S and ALP staining
DPSCs were fixed with 4% paraformaldehyde (PFA) for 1 hour and washed with phosphate-buffered saline (PBS). Cells were then stained with 40 mM alizarin red S (pH = 4.2) for 10 minutes under conditions of gentle agitation. Absorbance of the extracted alizarin red S stain was measured at 570 nm. DPSCs were subjected to ALP staining using the ALP assay kit (Jiancheng, Nanjing, China) according to the manufacturer's instructions.
Real-time RT-PCR
Total cellular RNA was extracted from the cells and reverse transcribed using conventional protocols. PCR amplification was performed using the following primer sets: GAPDH, forward: 5′-TCCATGACAACTTTGGTATCG-3′ and reverse: 5′-TGTAGCCAAATTCGTTGTCA-3′; DMP1, forward: 5′-TGGGGATTATCCTGTGCTCT-3′ and reverse: 5′-GCTGTCACTGGGGTCTTCAT-3′; DSPP, forward: 5′-GGAGACAAGACCTCCAAGAGTA-3′ and reverse: 5′-TGCTGGGACCCTTGATTTCTA-3′; Runx2, forward: 5′-AACCCACGAATGCACTATCCA-3′ and reverse: 5′-CGGACATACCGAGGGACCTG-3′; NRP1, forward: 5′-TGAGCCCTGTGGTTTATTCC-3′ and reverse: 5′-CGTACTCCTCTGGCTTCTGG-3′; Sema3A, forward: 5′-TGTACAGGTTTGTCTTTCTTCT-3′ and reverse: 5′-TTCATCAGCTTAGTAAATCCAT-3′.
All the primer sequences were determined using established GenBank sequences. The primers were used to amplify the duplicate PCRs. Each sample was calculated from three independent experiments and GAPDH was used as an internal control.
Western blot analysis
Total extracts of cells were lysed in buffer consisting of 150 mM NaCl, 50 mM TRIS, 2% sodium dodecyl sulfate (SDS), and a protease inhibitor mixture and centrifuged at 12,000 rpm for 12 minutes at 4°C. The proteins were subjected to sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride membranes at 350 mA for 2.5 hours in a blotting apparatus (Bio-Rad, CA). After blocking in Tris-buffered saline with Tween (TBST) with 5% nonfat dry milk, the membranes were incubated overnight at 4°C with primary antibodies. Then, the membranes were reacted with corresponding horseradish peroxidase-conjugated secondary antibodies at room temperature for 2 hours. GAPDH, β-actin, Lamin B, and β-tubulin were used as the internal control for the cytoplasmic and nuclear proteins.
The following primary antibodies were used: GAPDH (anti-mouse; Santa Cruz), β-actin (anti-mouse; Santa Cruz), Lamin B (anti-mouse; Santa Cruz), β-tubulin (anti-mouse; Sigma), DMP1 (anti-rabbit; Santa Cruz), DSPP (anti-rabbit; Santa Cruz), NRP1 (anti-rabbit; Santa Cruz), Sema3A (anti-rabbit; Santa Cruz), β-catenin (anti-mouse; Cell Signaling), p65 (anti-mouse; Santa Cruz), p-p65 (anti-mouse; Santa Cruz), and IκBα (anti-rabbit; Cell Signaling).
Immunofluorescent staining
DPSCs were seeded into the 24-well plates, fixed with 4% PFA for 1 hour, rinsed with PBS containing 0.1% Triton X-100 (PBST), and blocked for 30 minutes in PBST supplemented with 10% FBS. Cells were incubated with primary antibodies against STOR-1 (1:100) (anti-mouse; Santa Cruz), β-catenin (1:100) (anti-mouse; Cell Signaling), and p65 (1:100) (anti-mouse; Santa Cruz) in the same solution overnight at 4°C. Then cells were rinsed and incubated with secondary antibodies for 2 hours at room temperature. Nuclei were stained with DAPI (4060-diamidino-2-phenylindole dihydrochloride) (1:1000; Santa Cruz). Finally, the cells were examined using a Leica fluorescence microscope (Germany).
Statistical analysis
All experiments were repeated at least in triplicate, and values presented as mean ± standard deviation. The significance of differences between the experimental groups and controls was analyzed using ANOVA. Statistical significance was evaluated by the independent-samples t-test using SPSS v19.0 software. Differences in which p was less than 0.05 were considered to be statistically significant.
Results
Morphologic characteristics and odontoblast differentiation of DPSCs
The isolated DPSCs exhibited a fibroblast-like morphology (Fig. 1A). Based on our immunofluorescent staining, DPSCs were positive for STRO-1 (Fig. 1B). To confirm the odontoblast differentiation process of DPSCs, we measured the level of odontoblastic markers. During the progression of odontoblast differentiation, protein and mRNA levels of DMP1 and DSPP were gradually upregulated (Fig. 1C–E).
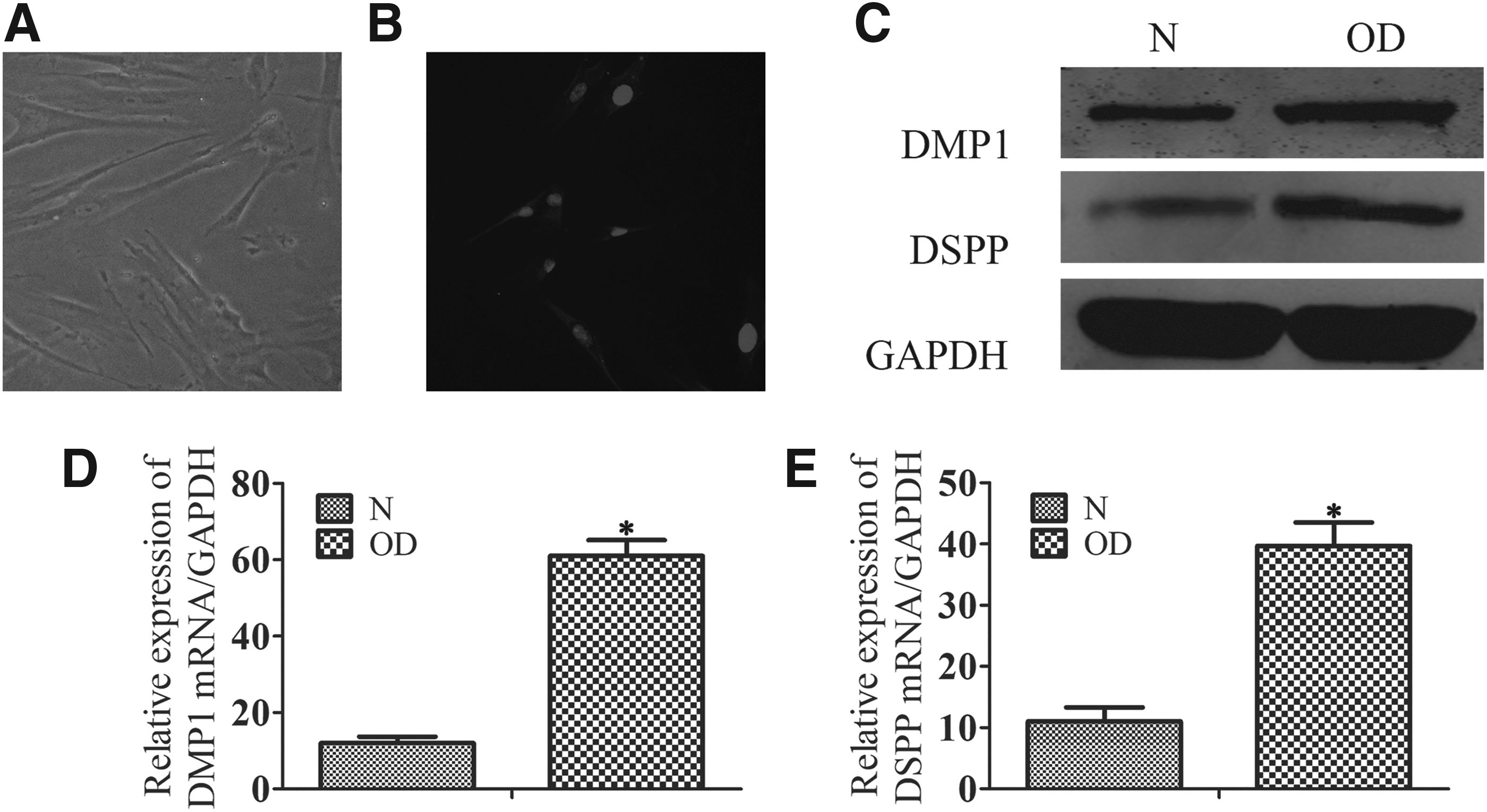
Morphologic characteristics and odontoblast differentiation of DPSCs. DPSCs were cultured in completed medium or odontogenic differentiation medium.
Wedelolactone accelerates DPSC odontoblast differentiation
To determine the effects of wedelolactone on odontoblast differentiation, we measured the activity of ALP, one of odontogenic markers, in a dose-dependent manner. Wedelolactone, 2.0 μg/mL, most significantly increased the number of ALP-positive DPSCs, while 5.0 μg/mL of wedelolactone treatment led to a decrease in the number of ALP-positive DPSCs, but the number was still more than that of the wedelolactone-untreated control group (Fig. 2A). Consistent with the ALP-positive cell results, the mRNA levels, alizarin red S staining for mineralized matrix, and ALP staining also displayed a similar change in DPSCs (Fig. 2B–E).
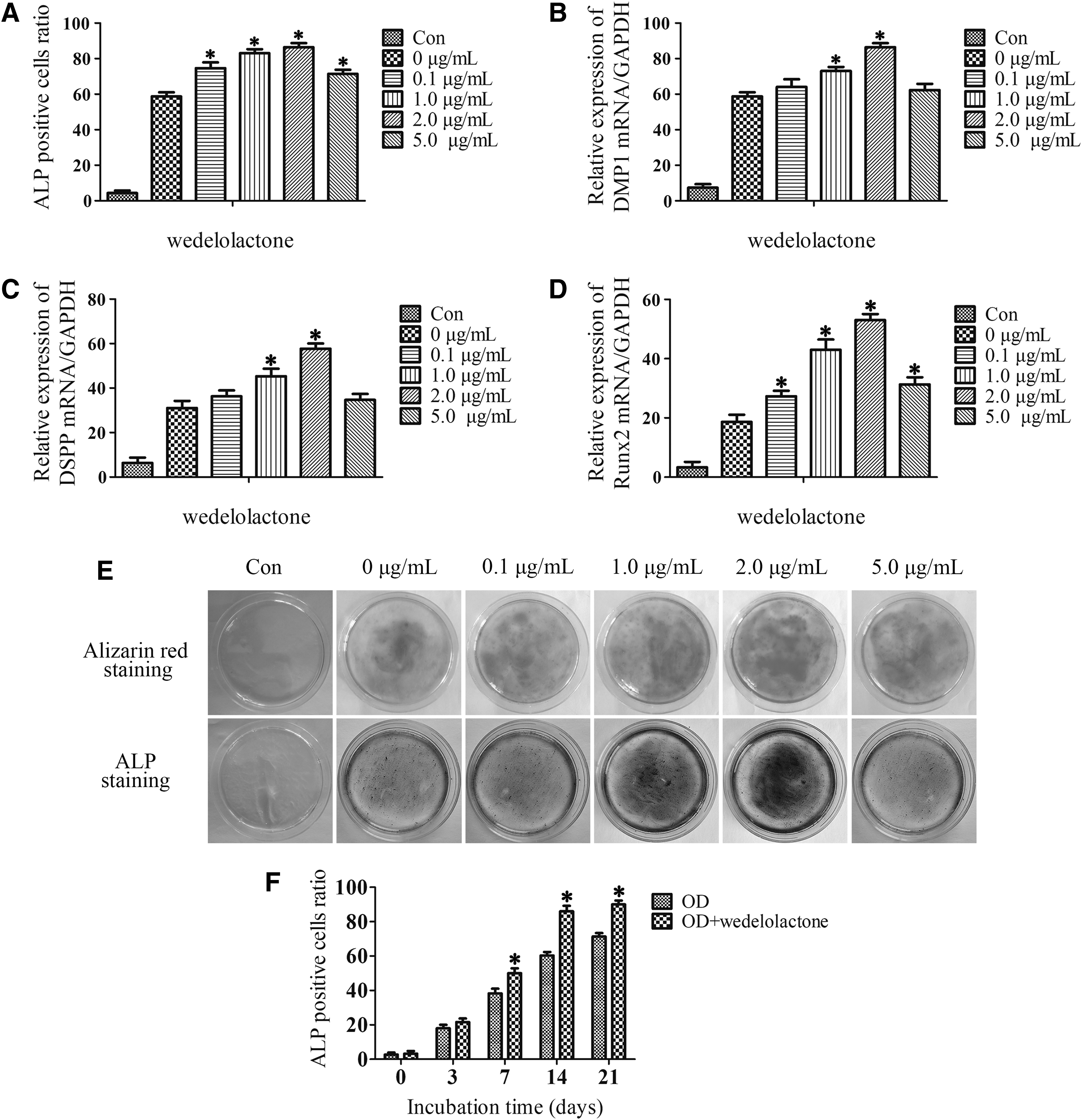
Wedelolactone accelerates DPSC odontoblast differentiation. DPSCs were cultured in odontogenic differentiation medium and wedelolactone was added (0, 0.1, 1.0, 2.0, and 5.0 μg/mL) for 0, 3, 7, 14, and 21 days.
Therefore, 2 μg/mL was selected as the optimal concentration for subsequent experiments. Meanwhile, the ALP-positive DPSCs significantly increased after 14 days in the wedelolactone (2 μg/mL)-treated group compared with the untreated control group in the odontogenic medium after incubation periods of 0, 3, 7, 14, and 21 days with or without wedelolactone (Fig. 2F). These data indicate wedelolactone may promote odontoblast differentiation of DPSCs.
Effect of wedelolactone on the production of Sema3A and NRP1 from DPSCs
Sema3A and NRP1 are reported to be produced from DPSCs and can accelerate odontoblast differentiation of DPSCs (Song et al., 2017, 2018; Yoshida et al., 2016). To examine the effects of wedelolactone on stimulating the production of Sema3A and NRP1 from DPSCs, western blot analysis and RT-PCR were performed. It was observed that the levels of Sema3A and NRP1 protein and mRNA expressions were upregulated after DPSCs were treated with wedelolactone (Fig. 3).
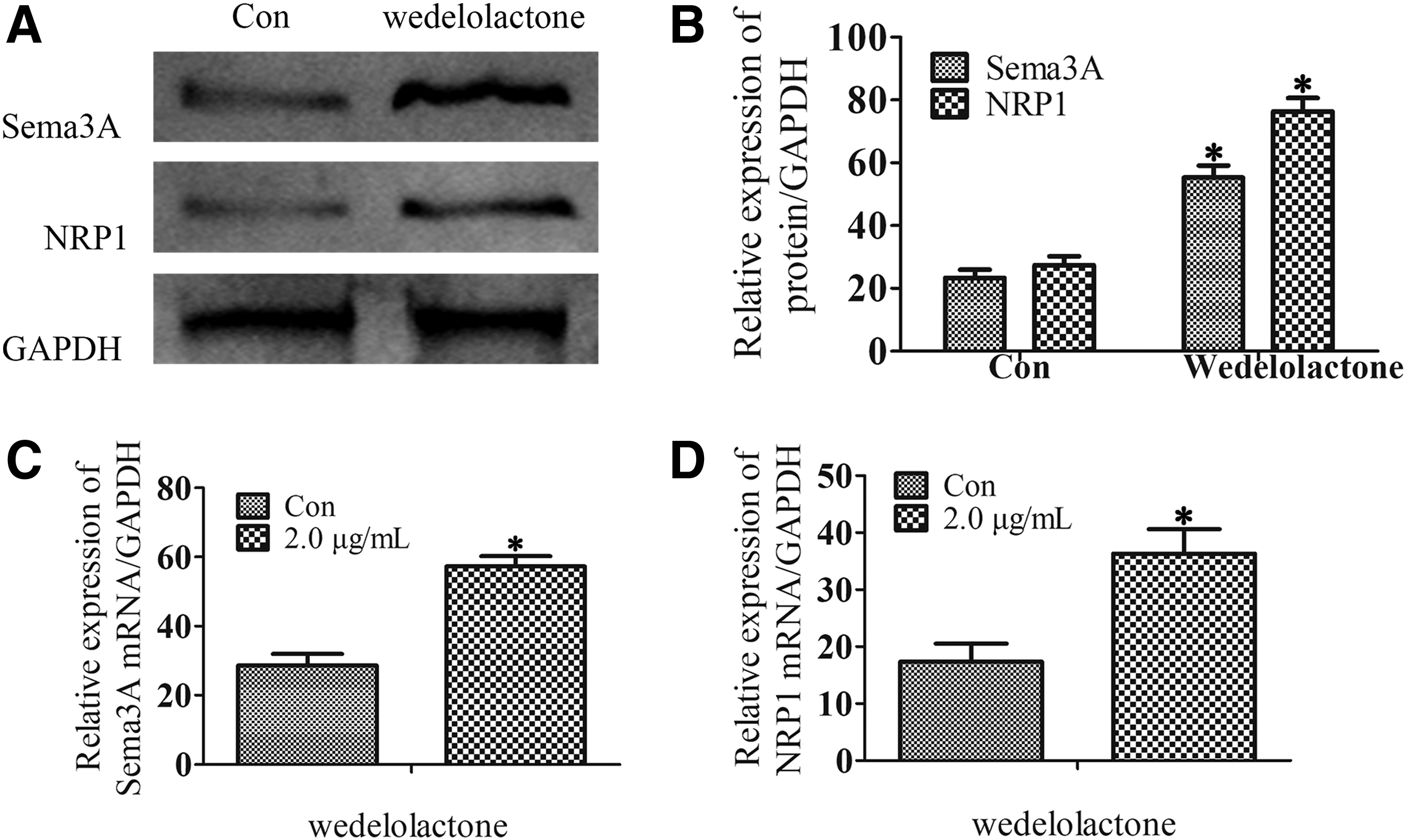
Effect of wedelolactone on the production of Sema3A and NRP1 from DPSCs. DPSCs were cultured in odontogenic differentiation medium and wedelolactone (2.0 μg/mL) was added for 21 days.
Wedelolactone regulates odontoblast differentiation through Sema3A/NRP1 pathway-mediated β-catenin activation
It is well known that the canonical Wnt/β-catenin pathway plays an important role in promoting odontoblastic differentiation of DPSCs (Lian et al., 2016). Our data also showed that NRP1 accelerated odontoblast differentiation of DPSCs through canonical Wnt/β-catenin signaling (Song et al., 2017). To confirm the mechanism by which wedelolactone promoted odontoblast differentiation, β-catenin was examined after treatment of DPSCs with wedelolactone.
Western blot showed that the nuclear β-catenin expression was dramatically higher in the wedelolactone groups compared with the control group (Fig. 4A–C). Furthermore, in the group treated with DKK1 still suppressed β-catenin nuclear translocation, even if the cells treated wedelolactone together (Fig. 4D–F). Immunofluorescence research also confirmed the same results (Fig. 4G). These data suggest a positive role for wedelolactone in differentiation of DPSCs through the canonical Wnt/β-catenin pathway.
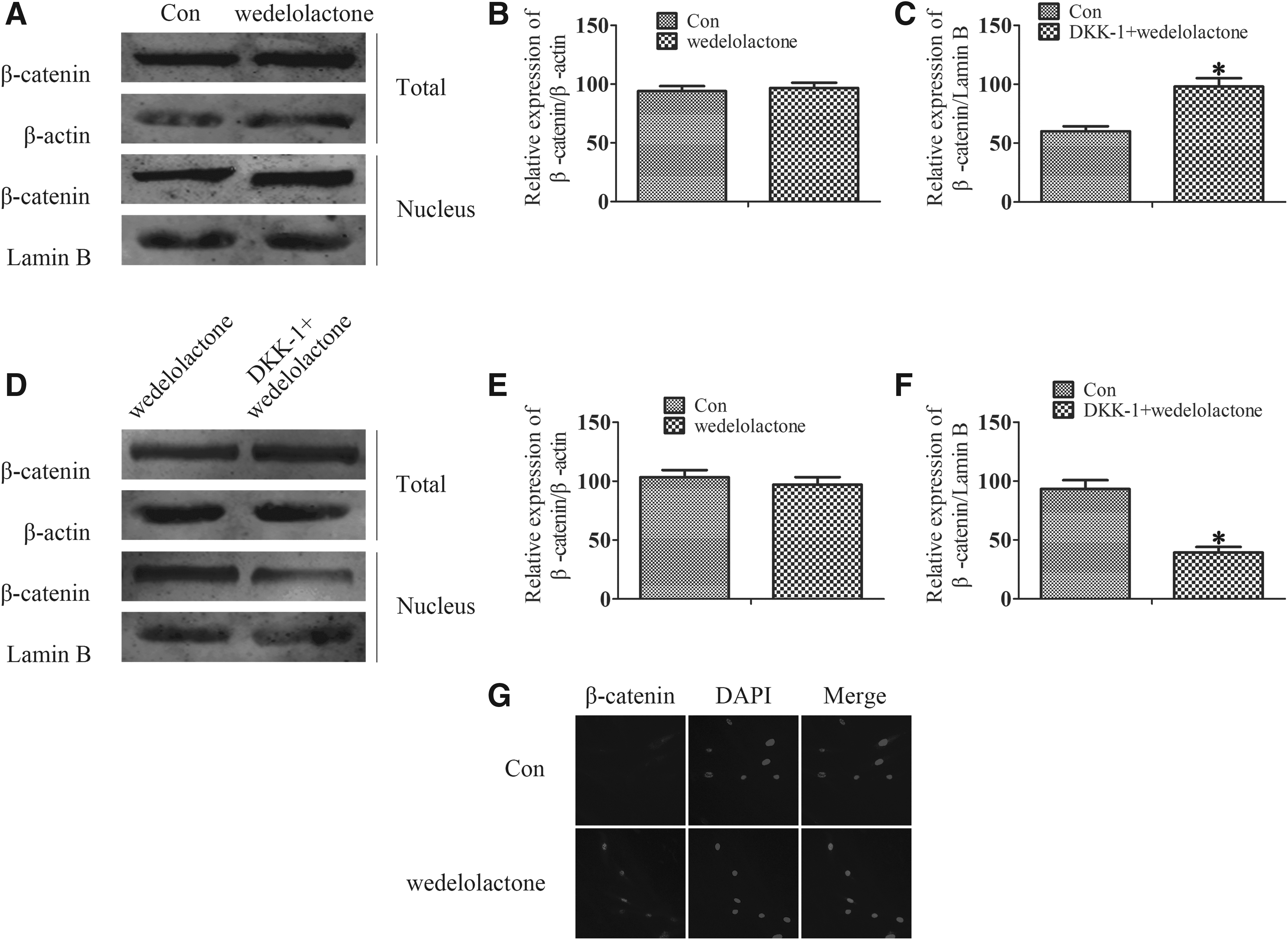
Wedelolactone regulates odontoblast differentiation through Sema3A/NRP1 pathway-mediated β-catenin activation. DPSCs were cultured in odontogenic differentiation medium and wedelolactone (2.0 μg/mL) was added for 21 days.
Wedelolactone regulates odontoblast differentiation through inhibiting NF-κB pathway
Several evidences indicate that chronic inflammation impedes DPSC differentiation and dentine reparative processes (He et al., 2005). There is, as everyone knows, NF-κB, a noted transcription factor, concerned in the process of inflammation. Wedelolactone was checked to see whether or not it exerted an inhibiting effect on this process. TNF-α improved the phosphonation of p65, the degradation of IκBα, and the nuclear migration of p65, whereas wedelolactone inhibited all these processes (Fig. 5A, B). BMS-345541, an NF-κB pathway inhibitor, exhibited similar to wedelolactone (Fig. 5C). Immunofluorescence results showed that p65 translocated to the nucleus following exposure to TNF-α, but wedelolactone effectively restrains TNF-α-induced p65 nuclear translocation (Fig. 5D). Together, these results demonstrate that wedelolactone can inhibit NF-κB pathway.

Wedelolactone regulates odontoblast differentiation through inhibiting NF-κB pathway. DPSCs were cultured in odontogenic differentiation medium and wedelolactone (2.0 μg/mL) or BMS-345541 was added with or without TNF-α (20 ng/mL) for 21 days.
Discussion
Dental defect is the damage or abnormality of the texture and physiologic anatomical appearance of teeth. It is often manifested as the damage of normal dental form, occlusion, and adjacent relation. There are many ways for treating dental defect, and the common methods are dental filling, drug therapy, and remineralization therapy, or other methods. However, most methods for dental defect normally can only inhibit the further destruction of the dental tissues without increasing tooth regeneration. Dentinal regeneration requires several physiological processes of DPSCs, containing cell proliferation, migration, differentiation, and mineralization.
Hence, we examined whether wedelolactone could accelerate odontoblast differentiation of DPSCs in vitro, and demonstrated that it remarkably induced odontoblast differentiation through Sema3A/NRP1 pathway-mediated β-catenin activation and NF-κB pathway inhibition (Fig. 6).
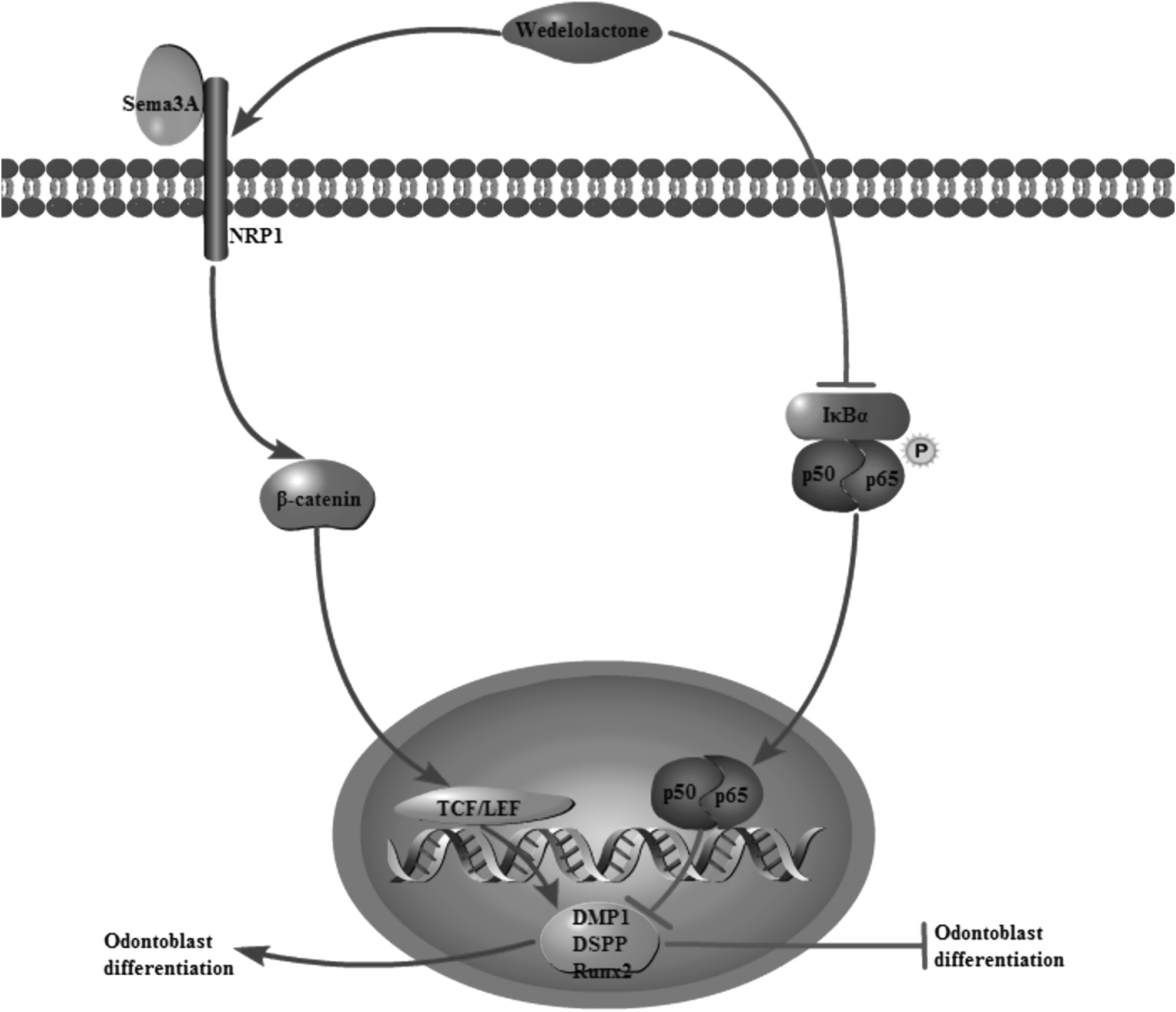
Schematic of wedelolactone regulates odontoblast differentiation. Wedelolactone activates canonical Wnt/β-catenin pathway, increases the expression of DMP1, DSPP, and Runx2, and promotes odontoblast differentiation of DPSCs. Wedelolactone also decreases p-p65, degrading the nucleus translocation of p65 of NF-κB pathway and enhancing the odontoblast differentiation capacity.
Wedelolactone is a natural polyphenolic catechol-type compound with antiosteoporosis effects that are exerted via multiple mechanisms/targets (Liu et al., 2014, 2016). Canonical Wnt/β-catenin pathway is crucial for regulation of osteoblast and DPSC development, including proliferation, differentiation, and survival. Activation of the canonical Wnt/β-catenin pathway involves translocation of β-catenin to the nucleus, regulation of Runx2, and activation of Wnt target genes (Nishikawa et al., 2010). Our results showed that wedelolactone was able to activate the canonical Wnt/β-catenin pathway in DPSCs, as shown by increase of β-catenin in nuclear fractions.
In addition, on 21 days of treatment with wedelolactone, the protein and mRNA expression levels of Sema3A and NRP1 increased. In DPSCs, NRP1 was shown to accelerate odontoblast differentiation, which induces the downstream activation of the canonical Wnt/β-catenin pathway (Song et al., 2017). These data suggested wedelolactone treatment resulted in the promotion of odontoblast differentiation.
Wedelolactone has been reported to inhibit the inflammatory response through regulating NF-κB-mediated transcription, indicating that wedelolactone is an inhibitor of NF-κB pathway (Yuan et al., 2013). In this study, the anti-inflammatory effect of wedelolactone and its regulating mechanisms in DPSCs induced by TNF-α were investigated. Further investigations about the effects of wedelolactone on NF-κB pathway were carried out and the result that wedelolactone could be similar to BMS-345541 to upregulate the expression of IκBα and to inhibit the phosphonation and the nuclear migration of p65, demonstrating that wedelolactone could suppress the activation of NF-κB pathway.
In conclusion, wedelolactone could be a novel medicine for the therapy of dental defect caused by the inflammation response or noninflammation response through affecting the canonical Wnt/β-catenin pathway and the NF-κB signaling pathway. However, the connections between wedelolactone and dental defect remain unclear. Wedelolactone may directly activate the expression of odontoblast genes to control odontoblast differentiation or also may indirectly control odontoblast differentiation by the production of proinflammatory cytokines. And there are also some limitations in this study.
What is the effect of wedelolactone participation in odontogenic differentiation in vivo? Does wedelolactone control odontoblast differentiation also through other signaling pathways? Does wedelolactone have an effect in the crosstalk network between the destruction of the dental tissues and the formation of reparative dentinogenesis? Further researches are warranted to address these questions.
Footnotes
Acknowledgments
The study was partly supported by Six Talent Peaks Project in Jiangsu Province (No. 2016-WSN-103) and Nantong Science and Technology Project (MS32015030).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.