
Abstract
Abstract
The immunomodulatory and self-renewable features of human adipose-derived mesenchymal stem cells (hAD-MSCs) mark their importance in regenerative medicine. Interleukin (IL)-23 as a proinflammatory cytokine suppresses T regulatory cells and promotes the response of T helper 17 and T helper 1 cells. This pathway initiates inflammation and immunosuppression in several autoimmune diseases. The current study aimed at producing recombinant IL-23 decoy receptor (RIL-23R) using hAD-MSCs as a good candidate for ex vivo cell-based gene therapy purposes to reduce inflammation in autoimmune diseases. hAD-MSCs was isolated from lipoaspirate and then characterized by differentiation. RIL-23R was designed and cloned into a pCDH813A-1 lentiviral vector. The transduction of hAD-MSCs was performed at multiplicity of infection = 50 with pCDH-EFI α-RIL-23R-PGK copGFP. Expressions of RIL-23R and octamer-binding transcription factor 4 (OCT-4) were determined by real-time polymerase chain reaction. Self-renewing properties were assayed with OCT-4. Bioactivity of the designed RIL-23R was evaluated by IL-17 and IL-10 expression of mouse splenocytes. The results showed that the transducted hAD-MSCs/RIL-23R, expressing IL-23 decoy receptor, can provide a useful approach for a basic research on cell-based gene therapy for autoimmune disorders.
Introduction
T
These characteristics of MSCs support the idea that creation of genetically modified MSCs could be the best option for combining cell and gene therapy to treat diverse forms of autoimmune diseases (Kumar et al., 2008; Lipsitz et al., 2017; Mundra et al., 2013; Myers et al., 2010; Park et al., 2015; Smith et al., 2018).
Adipose-MSCs (AD-MSCs) are a rich source for MSCs in the therapeutic purposes as adipose tissue is easily accessible and large amounts of AD-MSCs are easily obtainable (Aust et al., 2004; El-Badawy et al., 2016; Oedayrajsingh-Varma et al., 2006).
Currently, AD-MSCs are clinically applied for regenerative treatments and wound healing (Kanji and Das, 2017; Muhammad et al., 2017). Thus, the ability of human adipose-derived MSCs (hAD-MSCs) to serve as vehicles for cell-based gene therapy is promising (Bajek et al., 2016; Barranco, 2011; Bougioukli et al., 2018; Choi et al., 2015).
Autoimmune diseases are multifactorial disorders with complicated immune system dysregulation mediated by immune cytokines and immune cells (Onuora, 2016). Interleukin (IL)-23 plays an active role to proliferate the memory T helper 1 (Th1) cells. The hetero dimerized IL-23 receptor constituted of specific (IL23A) and common (IL12Rβ1) subunits (Wines et al., 2017). The transforming growth factor-β and IL-6 in most of the autoimmune diseases can induce T helper 17 (Th17) cells to increasingly secrete IL-23 and IL-17 (Zhou et al., 2007).
IL-23 is able to suppress the T regulatory cells and promote the response of Th17 and Th1 cells, initiating inflammation and immunosuppression in several autoimmune and inflammatory diseases (Villanueva, 2017). Based on strong evidence, the IL-23/IL-17 axis is important for the development of chronic inflammation (Gaffen et al., 2014). Recent studies determined that the suppression of IL-23, IL-23R, or IL-23/IL-17 axis potentially can be therapeutic targets for autoimmune diseases (Tang et al., 2012).
Targeting IL-23p19, but not IL-12p40, in gene knockout studies showed that the decrease of proinflammatory responses and resistance to different autoimmune diseases are due to the absence of IL-17-producing T cells (i.e., Th17 cells) (Cua et al., 2003).
Based on prior studies, extensive alternative splicing exerted on the IL-23R gene transcript (Kan et al., 2008). Generation of IL-23RΔ9 form (GenBank AM990318), which encodes a soluble version of the entire external domain of the specific receptor chain (IL23A), is an example of this splicing (Yu and Gallagher, 2010). After binding human IL-23 in solution, this soluble decoy receptor dependently inhibits STAT3 phosphorylation and functional maturation of human Th17 cells in vitro (Zhang et al., 2006).
Based on the immunosuppressive functions of hAD-MSCs and recombinant IL-23R (RIL-23R), they can cooperate to improve immunomodulation and prevent initiation of inflammation and subsequent autoimmune diseases. The current report was the first case of RIL-23R gene transduction into hAD-MSCs.
Materials and Methods
Cloning of RIL-23R complementary DNA into a lentiviral vector
The modified IL-23R complementary DNA (cDNA) was designed and amplified using the pORF-rIL23r plasmid (Biomatik, Cambridge, MA) using the forward primer of 5′-ACAACAGCTCGGCTTTGGTAT-3′, and reverse primer of 5′-TACTGGCAGCCTTGGAGTTC-3′. To generate the green fluorescent protein (GFP)-harboring lentiviral particles, CD813A-1 (System Bioscience, Mountain View, CA) and RIL-23R were digested by EcorV and Sal1, and then the cDNA was sub-cloned into pCDH813A-1 to achieve a recombinant vector (pCDH-EF-1α—RIL-23R-PGK-copGFP).
Generation, concentration, and titration of recombinant lentivirus
Recombinant lentivirus production was accomplished as previously described (Jiang et al., 2015; Klages et al., 2000). In summary, the transfection of plasmid DNA into 1 × 106 HEK-293T cells was based on calcium–phosphate method (21 μg of lentivirus involving pCDH-RIL-23R-copGFP, 21 μg of packaging plasmid psPAX2, and 10.5 μg of envelope plasmid per 10-cm plate). The lentivirus particles contained in the supernatant of HEK-293 T cells were collected at 36, 48, 60, and 72 hours post-transfection and then passed through a 0.45 μm filter.
The concentration process of lentivirus particles were performed according to precipitation method using 50% PEG-8000 (Sigma-Aldrich, St. Louis, MO) to reach final concentration of 5%. The concentration was continued with NaCl 5 M (Sigma-Aldrich) overnight to achieve final concentration of 0.15 M. Then, 6 × 104 HEK-293T cells were poured into the plate for titration, followed by contamination by 1, 4, and 16 μL of virus. Three days later, the cells detached and their fluorescence was analyzed by flow cytometry.
Isolation of MSC from human adipose tissue
Lipoaspirate samples were washed with phosphate-buffered saline (PBS) containing 3 × penicillin/streptomycin and amphotericin three times. The adipose tissue was added dispase (50 u/mL)/collagenase I (250 U/mL) (Sigma-Aldrich), followed by shaking for 30 minutes at 37°C and then centrifugation at 1500 rpm. The plated cells after suspending were distributed in the flasks with α-MEM (minimum essential medium eagle-alpha modification) containing 10% fetal bovine serum (FBS) for 3 days.
Transduction of hAD-MSCs
The trypsinization was done on the second passage hAD-MSCs that were seeded in six-well plates at a density of 1 × 105 cells in 1 mL Dulbecco's modified Eagle's medium (DMEM)-F12. The addition of virus (multiplicity of infection [MOI] = 50) was immediate along with 6 μg/mL of Polybrene (Sigma-Aldrich), followed by shaking on a rotator at 5 rpm for 18 hours. Next, the medium was renewed by DMEM-F12 containing 10% FBS. The flow cytometry was applied to calculate the GFP-positive transducted hAD-MSCs at 72 hours after transduction. The driven data were analyzed by BD Accuri c6 software.
Differentiation of hAD-MSCs
In this step, 2 × 104 cells per mL of third passage transducted hAD-MSCs/RIL-23R and not-transducted hAD-MSCs were poured in a six-well plate. The incubation was performed with α-MEM containing 10% FBS until reaching confluence. The medium was replaced by osteogenic medium with DMEM containing 10 μM dexamethasone (Sigma-Aldrich), 50 μg/mL ascorbic acid 2-phosphate (Sigma-Aldrich), and 10 mM β-glycerol phosphate (Sigma-Aldrich). The action was done twice a week for 21 days. At the end of the cultivation period, the cells were fixed with 10% formalin for 10 minutes and stained with Alizarin Red (Sigma-Aldrich) at room temperature for 2 minutes.
In adipogenesis, the third passage not-transducted and transducted cells were incubated with a differentiation medium of DMEM, supplemented with 50 μg/mL indomethacin (Sigma-Aldrich) and 100 nM dexamethasone (Sigma-Aldrich). The medium was renewed twice a week for 21 days. At the end of this period, the cells staining was performed with 0.5% Oil Red O (Sigma-Aldrich) in methanol for 2 minutes at room temperature.
Flow cytometry
The trypsinization of cells after third passage was carried out by 0.025% trypsin and 0.02% ethylene diamine tetra acetic acid (EDTA). The washing was continued twice using PBS, and staining was based on the manufacturer's instruction for running flow cytometry. The cells were labeled with the following antibodies (Nery et al., 2013): phycoerythrin (PE)-conjugated mouse antihuman CD11b, PE-conjugated mouse antihuman CD34, PE-conjugated mouse antihuman CD45, PE-conjugated mouse antihuman CD105, and PE-conjugated mouse antihuman CD73. PE-conjugated mouse immunoglobulin G (IgG)1, fluorescein isothiocyanate (FITC)-conjugated mouse IgG2, and PE-conjugated rat IgG2 were considered to be isotype controls (all the antibodies were purchased from e-Bioscience, San Diego, CA).
After incubation, to remove any unlabeled antibodies, the cell suspension washed with PBS. The cells were suspended in PBS and analyzed using FACScan flow cytometer (Becton Dickinson, San Diego, CA) and the data were analyzed using BD Accuri c6 software.
Bioactivity assay
The IL-23R secretion and function were investigated by bioassays of naïve T cells of C57BL/6 mouse cocultured with normal/transducted hAD-MSCs and HT1080 cells. The mice were killed in accordance with the laboratory animal protocol to dissect the spleen for digesting with 100 u/mL of dispase/collagenase. The spleen cells were gathered via centrifugation at 1200 rpm, and the red blood cells (RBCs) lysis via an RBC lysis buffer. The cells were then suspended in RPMI (Roswell Park Memorial Institute) culture medium supplemented with 10% fetal calf serum at a concentration of 1 × 106 cells per mL. To generate short-term cultures of activated T cells, splenocytes were activated by the addition of lectin phytohemagglutinin (PHA) at a concentration of 1 mg/mL in RPMI supplemented with 10% fetal calf serum for 4 days (Crowston et al., 2004).
The cells were then washed and maintained in IL-2 supplemented media at a concentration of 2 ng/mL for at least 7 days before use. A total of 2 × 104 cells per well were seeded in 96-well plates. The next day, cells were washed and preincubated with mitomycin C (0.4 mg/mL for 5 minutes) (Khaw et al., 1992). Following treatment the cells were washed immediately at least three times and incubated in RPMI (Sigma-Aldrich) growth medium. To determine whether transducted cells induced T helper 2 (Th2) cells proliferation in results from upregulated RIL-23R expression, IL-2-dependent PHA-activated T cells were added to the culture medium with an effector: target cell ratio of 20:1 (Gombert et al., 1996). T cell—adherent cell contact was inhibited in some experiments by seeding T cells into cell culture inserts (Becton Dickinson).
These inserts incorporate membranes that are transparent and contain 0.4 mm pores that allow free passage of soluble factors but are too small to permit cell migration (Crowston et al., 2004). Supernatants were harvested after 24 or 48 h and concentration of IL-10 and IL-17 determined by real-time polymerase chain reaction (PCR) (Sapski et al., 2017). Total RNA extraction was carried out from the T cells cocultured with normal/transducted cells. The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was considered as an endogenous reference gene.
Expression of RIL-23R and self-renewing assays
The octamer-binding transcription factor 4 (OCT-4) and RIL-23R expression was studied in hAD-MSCs and lentiviral-engineered hAD-MSCs. The GAPDH expression was the endogenous reference gene.
RNA extraction and real-time PCR
The RIL-23R expression was studied in hAD-MSCs and transducted hAD-MSCs. The GAPDH expression was the endogenous reference gene. Qiazol lysis reagent (Alameda, CA) was used to isolate total RNA from the cells. The reactions of standard reverse transcription were done using 5 μg of total RNA by oligo (dT) 18 as a primer according to the manufacturer's instructions for the cDNA synthesis kit (Fermentas). Additional PCR components were 2.5 μL of cDNA, 1 × PCR buffer (AMS), 200 μM of dNTPs, 0.5 μM of each primer pair, and 1 unit per 25 μL of reaction mix Taq DNA polymerase (Fermentas). The abovementioned pairs of primers detected the expression of RIL-23R gene.
Whole cell lysate of normal/transducted hAD-MSC was generated from confluent cell layers in six-well plates using lysis buffer made of 50 mM Tris-Cl, pH 7.4; 150 mM NaCl, 1% NP-40, 1% Na-deoxycholate, 1 mM EDTA, 0.1% sodium dodecyl sulfate (SDS) (all from Sigma-Aldrich) and Mini Complete, EDTA-free protease inhibitor cocktail (Roche). Proteins were separated by 12% SDS-PAGE, blotted to nitrocellulose membranes (GE Healthcare, Munich, Germany), blocked with tris-buffered saline (TBS) containing 5% nonfat dry milk, and analyzed for RIL-23R with anti-RIL-23R antibody (sc-293485) followed by conjugate-IgGκ BP-HRP (Santa Cruz, Santa Cruz, CA).
Ethical considerations
The human adipose tissue was collected after obtaining informed consent considering the Declarations of the Golestan University of Medical Sciences, Iran.
Statistical analyses
GraphPad Prism version 6 was used to perform statistical analyses. One-way analysis of variance and t-test were used to evaluate the differences between the groups. p-Value <0.05 was considered statistically significant.
Results
Cloning of RIL-23R cDNA in lentiviral vector
The recombinant open reading frame of hIL-23R gene (RIL-23R) was designed and amplified by PCR and sub-cloned into the lentiviral vector (Fig. 1A). To indicate the insertion of RIL-23R cDNA into the pCDH813A-1 vector, the shuttle vector was digested by EcorV and Sal1. The 1400 bp fragment that belonged to RIL-23R was successfully separated from the pCDH813A-1 vector (Fig. 1B). Moreover, RIL-23R cDNA was detected by colony-PCR in this vector. The cDNA was subsequently sequenced. No mutation was detected in cDNA sequence.
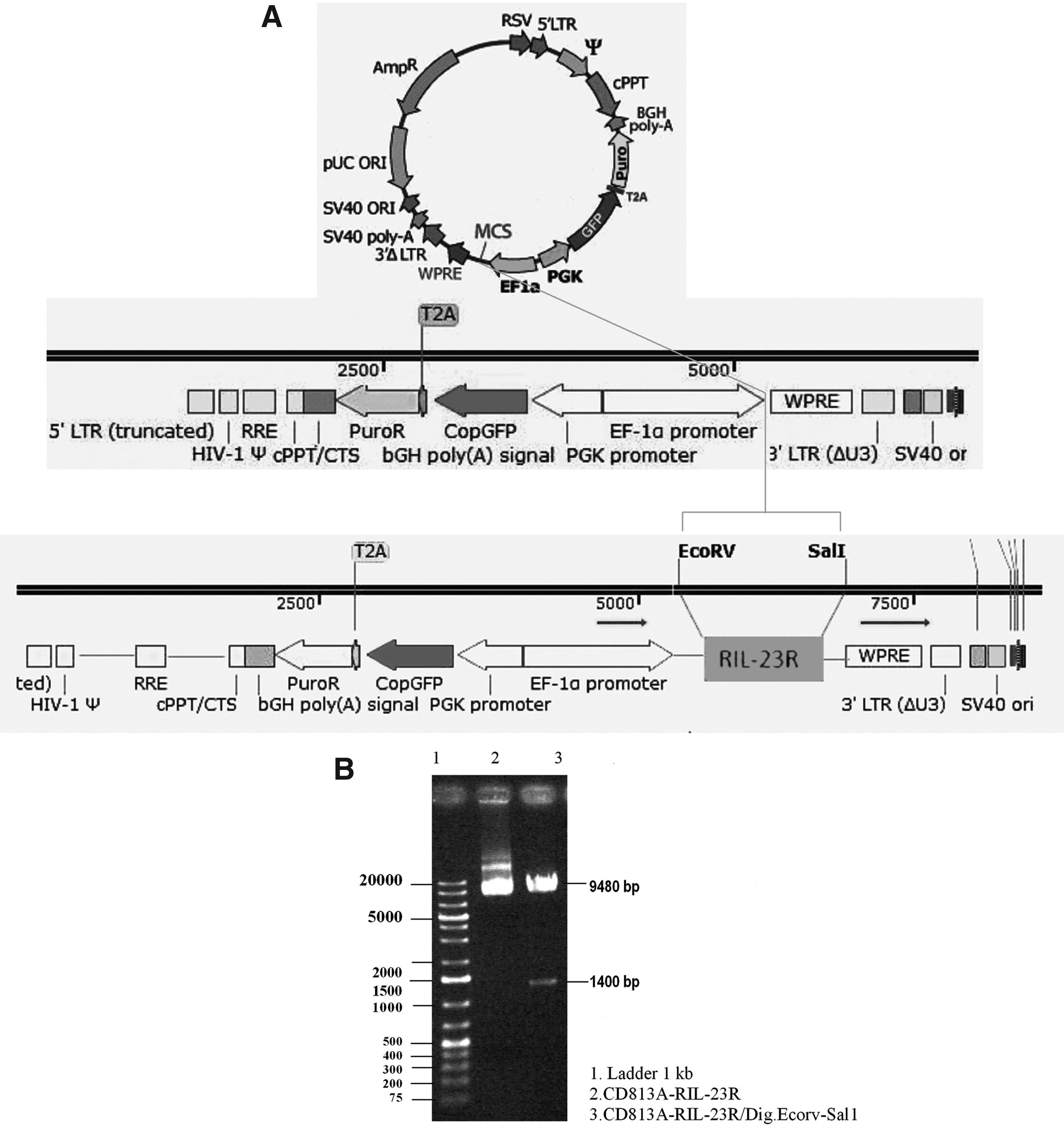
RIL-23R gene construct.
Recombinant viral particle production
This construct was co-transfected in HEK-293T cells (70% confluency) with the helper packaging vector-mediated Ca3 (PO4)2 protocol and successfully produced RIL-23R-lentiviral particles. The transfection efficiency was over 90% (Fig. 2). After concentration and titration of the virus, hAD-MSCs was transducted at MOI = 50.

Transfection of HEK-293T for recombinant viral particles production.
Isolation of adipocyte stem cells and differentiation into adipocytes and osteoblasts
Adipocyte cells were isolated from liposuction tissue with mechanical and enzyme digestion. After the cells were passaged, they displayed a strong proliferative ability (Fig. 3A). The confirmation of the cell multilineage capacity was done following their differentiation into adipocyte and osteoblast cells after 3 weeks. Observation of massive calcium depositions around differentiated cells after Alizarin Red staining confirmed the presence of osteoblasts in not-transducted hAD-MSCs (Fig. 3B) and transducted hAD-MSCs/RIL-23R (Fig. 3E).
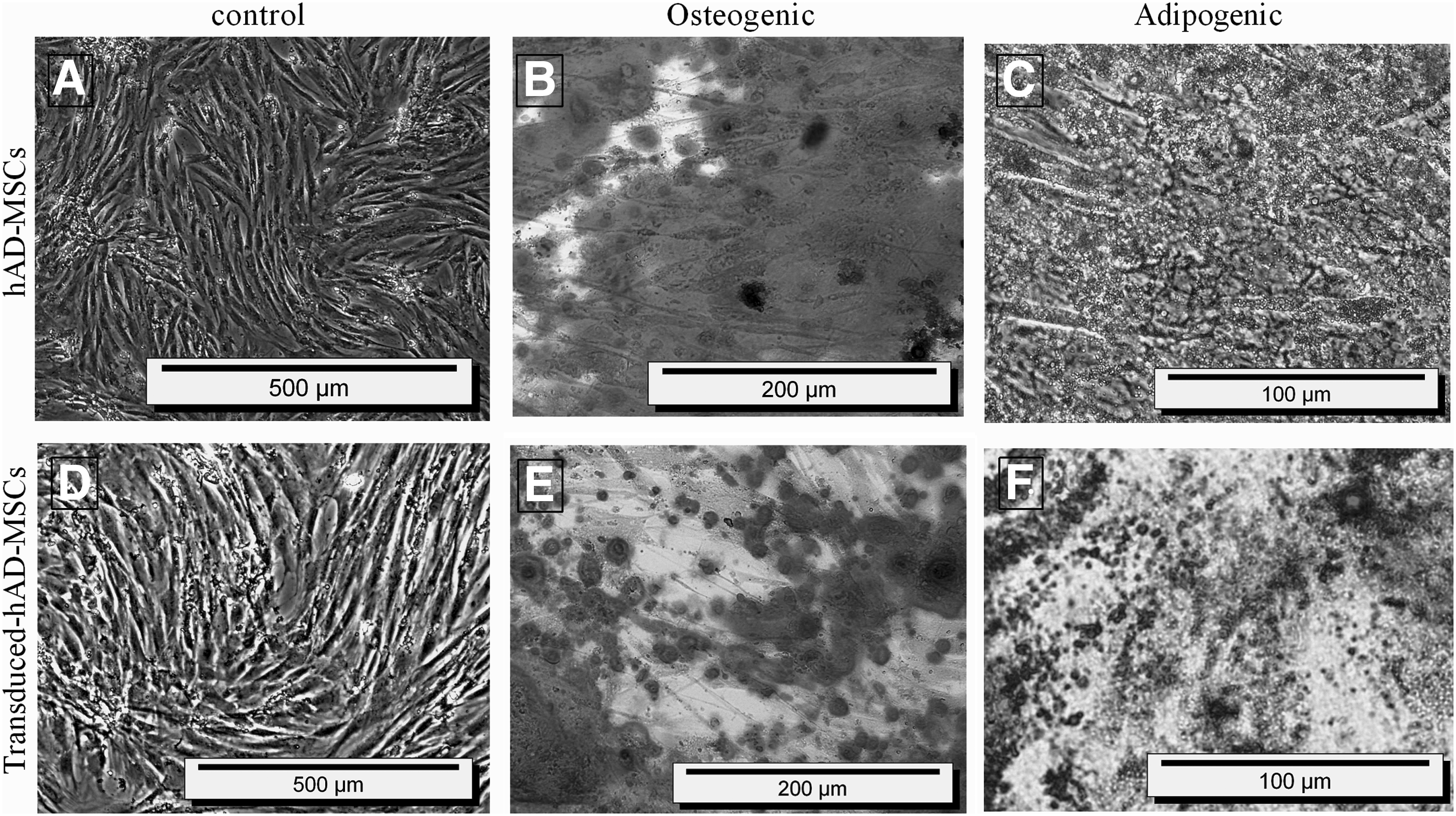
hAD-MSCs isolation from adipose tissue and characterization by differentiation.
Moreover, visualization of numerous lipid vacuoles after Oil Red staining revealed the adipocyte features of not-transducted hAD-MSCs (Fig. 3C) and transducted hAD-MSCs/RIL-23R (Fig. 3F). Therefore, lentiviral particles containing RIL-23R preserve the mesodermal properties of hAD-MSCs.
Characterization of hAD-MSCs by flow cytometry
We identified the presence of cell surface marker CD73, CD105, and the absence of hematopoietic (CD45, CD11b) and endothelial (CD34) antigens in not-transducted, transducted adipose MSCs/CD813A-1 and transducted adipose MSCs/RIL-23R (Fig. 4). Our data indicate that these three types of cells have similar expression profiles for the selected markers. These results also showed that the MSCs were not derived from endothelial or hematopoietic cells.
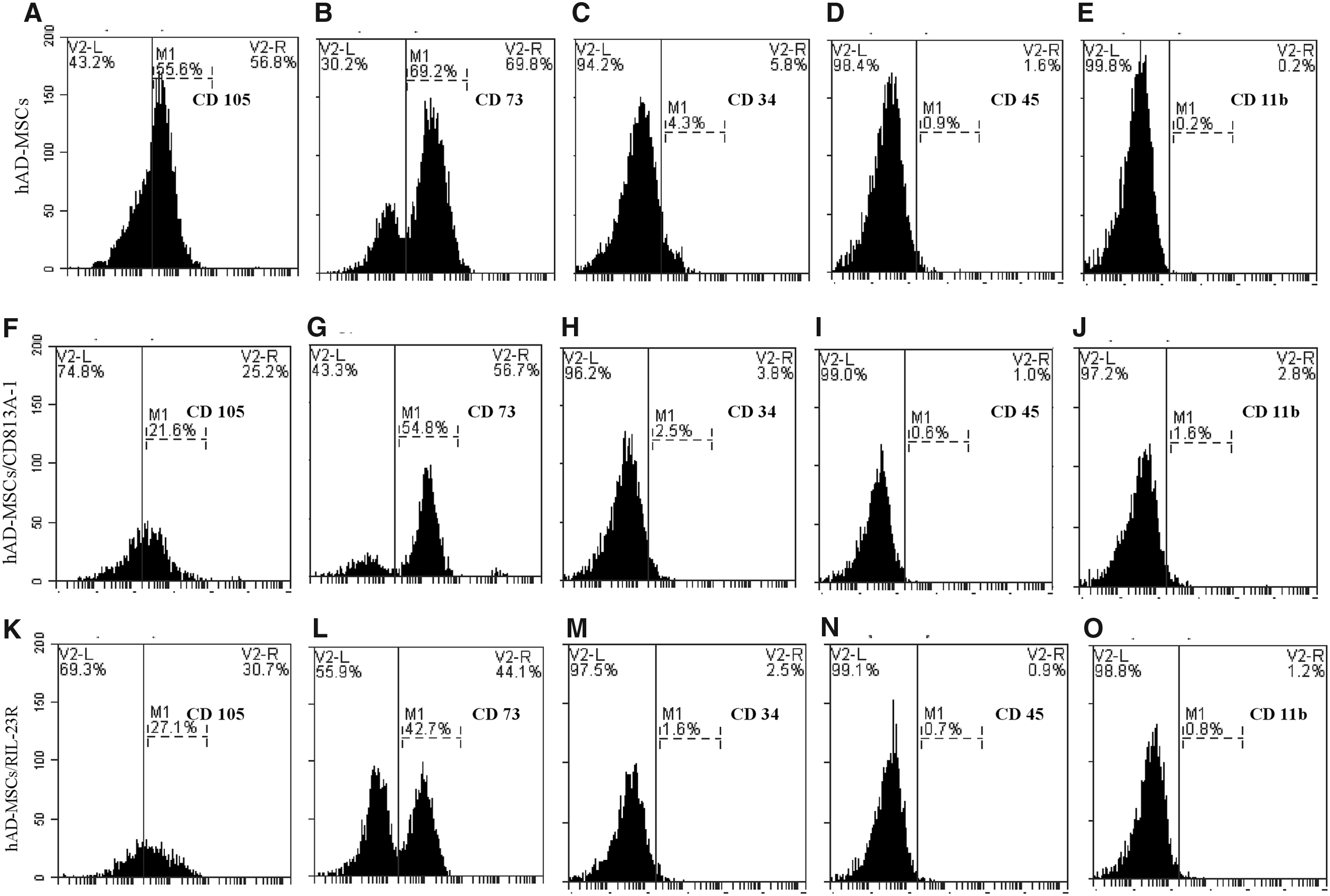
Expression of surface antigens in hAD-MSCs (
Transduction with recombinant lentiviral particles
hAD-MSCs were approximately 50% confluent when transducted with lentiviral particles (Fig. 5A). GFP-positive transducted hAD-MSCs/RIL-23R measured 72 hours post-transduction. The bright field image and GFP image of the same cells was shown in Figure 5B and C. The cells were selected by 2.5 μg of puromycin for 2 weeks. The bright field image and GFP image of the same cells was shown in Figure 5D and E. Flow cytometry was used to obtain ∼95% pure transducted cells 14 days post-transduction (Fig. 5F).

hAD-MSCs transduction with lentiviral particles.
RIL-23R and OCT-4 expression
For RIL-23R expression, total RNA of not-transducted hAD-MSCs and transducted hAD-MSCs/RIL-23R were isolated, and real-time PCR showed expression of RIL-23R in transducted hAD-MSCs/RIL-23R but not in not-transducted hAD-MSCs (Fig. 6A, B). Subsequently, the expression of RIL-23R was determined on protein level by immunoblot analysis, RIL-23R was virtually detectable in whole cell lysate of transducted hAD-MSCs (Fig. 6C).
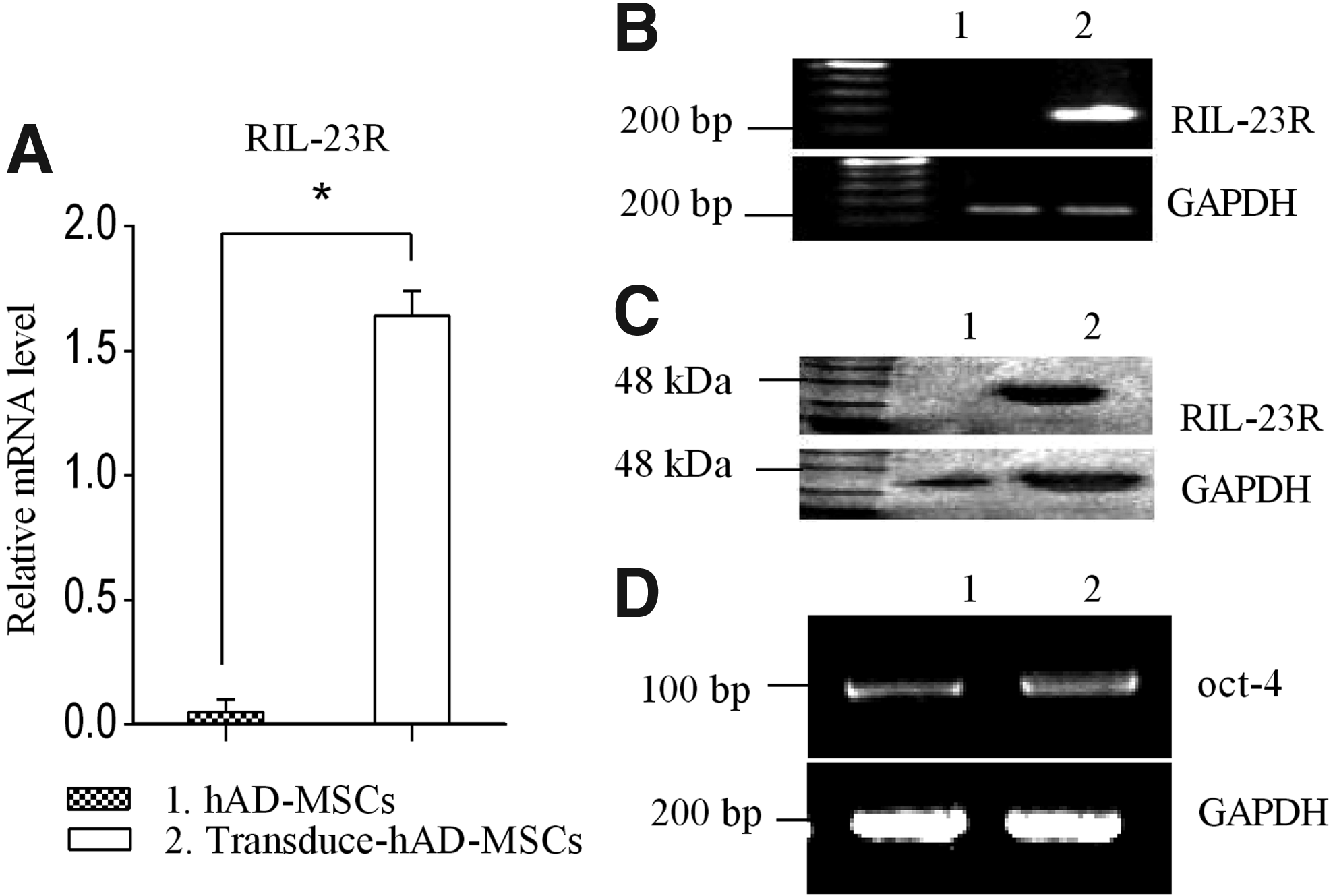
Expression of RIL-23R and OCT-4 in hAD-MSCs and transducted hAD-MSCs.
The OCT-4 expression confirmed that transducted hAD-MSCs/RIL-23R preserved their self-renewing potency and lentiviral transduction did not affect the mesenchymal properties (Fig. 6D).
RIL-23R functional assay
To demonstrate that the inhibitory effect of hMSCs on murine T cells is specific to the transducted hAD-MSCs we also used a different adherent human cell line in our experiments, HT-1080 (human fibrosarcoma cell line). These cells have similar morphological features to hMSCs, without having any known MSC-like properties (the multipotent ability to differentiate into osteocytes, chondrocytes, and adipocytes); also the fibroblasts do not have any inhibitory effect on the T cells (Nazarov et al., 2013), therefore, they were used as controls in our experiments. We used these cell lines in the same ratios as the hAD-MSCs, and the same experimental settings.
We hypothesized that RIL-23R can suppress inflammation mediated by IL-17 by its inhibition of IL-23. This function of RIL-23R was examined by coculturing of normal/transducted hAD-MSCs and HT1080 cells with splenocyte CD4+ T cells and measuring the level of IL-17 and IL-10 by real-time PCR. To determine whether inhibition was due to RIL-23R and not a property of hAD-MSCs, the CD4+ T cells were cocultured with transducted hAD-MSCs and transducted HT1080 cells compared with naïve T cells cocultured with hAD-MSCs and HT1080 cells.
Real-time PCR showed the IL-10 expression (Fig. 7A) and suppression of IL-17 expression (Fig. 7B) in T cells cocultured with transducted hAD-MSCs and transducted HT1080 cells compared with T cells cocultured hAD-MSCs and HT1080cells. Results of quantitative real-time PCR confirmed the presence of a definite transcript by gel resolution (Fig. 7C).
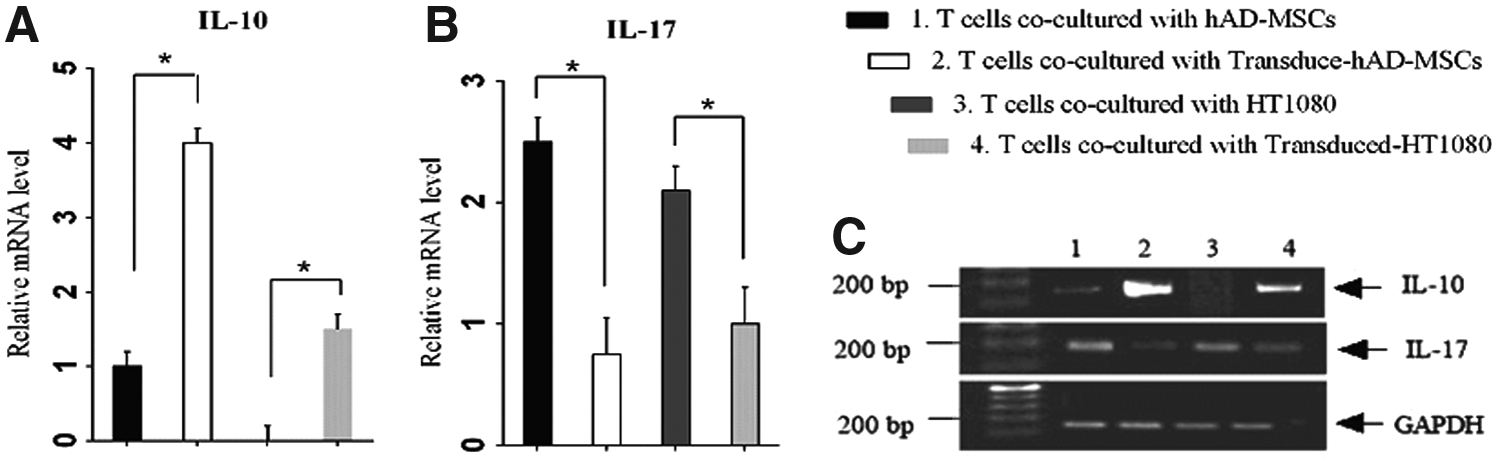
Expression of IL-17 and IL-10 in T cells cocultured with normal/transducted hAD-MSCs and HT1080 cells.
Discussion
Autoimmune disease occurs when body organs are attacked by autoimmune cells as a result of an unfit immune response directed to autoantigens (Collison, 2018). The most immunosuppressive drugs for the treatment of autoimmune disease belong to the corticosteroids family (Hartono et al., 2013). Many medical investigators are seeking to find new immunotherapeutic strategies with lower side effects. Such strategies include gene or recombinant protein (Khan et al., 2016) therapies aimed at affecting specific immune cells and molecules such as cytokines (Qian et al., 2006), chemokines (Grivennikov et al., 2010), and costimulatory molecules (Podojil and Miller, 2009).
It is noted that MSCs have immunomodulatory, self-renewing, and multilineage differentiation properties (Karp and Leng Teo, 2009; Kfoury and Scadden, 2015; Khaldoyanidi, 2008). These characteristics have led to recognition of the true capacity of MSC-based cell therapy. Due to their efficiency to be transducted with different genes (Bougioukli et al., 2018; Mohammadi et al., 2016; Mundra et al., 2013; Myers et al., 2010; Park et al., 2015). Genetic modifications of these cells are performed to enhance the MSCs efficacy in tissue repair/regeneration (Trounson and McDonald, 2015).
The high amount of MSCs found in adipose tissues and relative easy isolation make them a very good source of adult stem cells for further use in regenerative medicine applications. Some studies (Aust et al., 2004; El-Badawy et al., 2016; Oedayrajsingh-Varma et al., 2006) have shown that manifestly more stem cells can be obtained from adipose tissue compared with the same volume of bone marrow.
The current study successfully isolated hAD-MSCs from lipoaspirate tissue samples and characterized them by flow cytometry and osteogenic and adipogenic differentiation. The hAD-MSCs showed specific cell surface expression types with high expression of CD105, CD90, CD44, and CD73 and low expression of CD34, CD14, and CD45 (Kocan et al., 2017). Our isolated cells showed CD90 and CD73, but not CD34 and CD45. Additionally, the expression of CD11b as an adhesion molecule was negative, which may help cells with better migration (Baer, 2014). These results confirmed successful isolation of hAD-MSCs.
IL-23R as a heterodimeric receptor consists of a common IL-12 receptor beta 1 (IL-12Rβ1) and a specific IL-23R receptor (IL23A or IL-12Rβ2). The natural form of IL-23R is encoded by at least 12 exons. Here, we designed RIL-23R that can produce a secretion form of the receptor with an antagonistic function against IL-23p19 (Kan et al., 2008; Yu and Gallagher, 2010; Zhang et al., 2006). Some studies have shown that the IL-23R exhibited tumor suppressor functions (Airoldi et al., 2004). In addition, evidence indicates that soluble receptors play roles as agonists or antagonists during disease and normal homeostasis (Felix and Savvides, 2017; Wessels et al., 2011). Therefore, it was proposed that the generation of this decoy receptor might provide immune maintenance in autoimmune diseases.
The MSCs can be used as a gene delivery system because of their ability to be easily transformed and home in on injured tissues and their lack of immunogenic properties (Karp and Leng Teo, 2009; Khaldoyanidi, 2008). Additionally, they have been transducted with different vectors to optimize transgene expression. In some cases, such as therapy for long-life illnesses, permanent transduction of MSCs may be needed and can be attained by using retroviruses to modify MSCs with high efficiency to get long-term expression.
It was demonstrated that transduction of hAD-MSCs with lentiviral vector expressing RIL-23R can improve the survival of RIL-23R secretion in cell transplantation.
The current study obtained more than 95% lentiviral transduction efficiency of AD-MSCs according to the selection of puromycin, which was a beneficial agent for optimal therapeutic purposes (Ben-Dor et al., 2006). All transducted and not-transducted cultures were capable of differentiating to adipogenic and osteogenic lineages, could express stem cell markers such as OCT-4, and retained the nature of hAD-MSCs. There were no significant variations in proliferation capacity or cell surface marker expression between transducted and not-transducted cultures. Transducted cells, similar to not-transducted cells, demonstrated similar morphology and differentiation (van Vollenstee et al., 2016).
Real-time PCR verified the expression of RIL-23R. The current study results and those of other similar studies did not note any negative effects of transgene expression by lentiviral transduction on multipotencial properties.
Transcription factor of OCT-4 expression was similar in both hAD-MSCs and hAD-MSCs/RIL23R.
The IL-23 is effective in the function of Th17 cells but unable to induce the differentiation of these cells in vitro. The ability of the soluble isoform of IL-23R to inhibit IL-23p19 signaling caused the supposition that RIL-23R, as an IL-23 inhibitor, may bind to the p19 chain of IL-23 and inhibit the functional maturation of Th17 cells, resulting in the generation of T cells that secrete large amounts of IL-10 to shift to Th2 cell type (Payne et al., 2013). Then, overexpression of IL-10 and suppression of IL-17 were expected. Real-time PCR can show expression level of genes; however, bioassay exactly confirmed gene expression potency. Mouse CD4+ T cells were isolated from splenocyte and cocultured in the presence of not-transducted HT1080 and transducted HT1080/RIL-23R cells for 3 days.
The ability of the RIL-23R to suppress IL-17 and express IL-10 was analyzed by real-time PCR. RIL-23R induced the proliferation of Th2 cells and improved their function by IL-10 expression.
In conclusion, it was the first report of RIL-23R transduction into hAD-MSCs, which may be an effective approach to use such cells as a good vehicle for cell-based gene therapy in autoimmune disorders.
Footnotes
Acknowledgment
This study was supported by grants from the Research and Technology Council of Golestan University of Medical Sciences (Grant No. 940805182).
Ethics Approval and Consent to Participate
This study was conducted with the recommendations in the guide for the Care and Use of Laboratory Animals and was approved by the Ethics Committee of the Golestan University of Medical Sciences, Gorgan, Iran.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.