
Abstract
Abstract
In our previous study, we found that osthole could promote the ability of proliferation and differentiation in normal neural stem cells (NSCs) under normal condition. Then, we used tert-butyl hydroperoxide (t-BHP) to establish the model of senescence NSCs to detect the effects of osthole. Interestingly, the immunofluorescence results showed that osthole (100 μM) could enhance the ability of proliferation and differentiation, and CCK-8 assay results showed that osthole could also enhance the cell viabilities. Then, SA-β-gal assay results showed that osthole could decrease the positive of senescence cells. Flow cytometric analysis results showed that osthole could decrease the mixture of G0 and G1 phase. Reverse transcriptase (RT)-polymerase chain reaction results showed that osthole could downregulate the expression of p16 mRNA, and western blot analysis results showed that the expressions of the target protein decreased in p16-pRb signaling pathway with osthole treatment. In conclusion, these results indicated that osthole could probably delay cells senescence through p16-pRb signaling pathway.
Introduction
A
Nowadays, the theory of stem cells senescence became the latest one that had explained the body aging up to now. Also, neural stem cells (NSCs) possessed the capacity of self-renewal and differentiation into neural cells, which induced neurons, oligodendrocyte, and astrocyte. NSCs aging negatively affects the proliferation and differentiation, and it is associated with pathogenesis of several neurological and neurodegenerative disorders, including Alzheimer's disease (AD) and aging-related neural disorders (Kuhn et al., 1996).
Tert-butyl hydroperoxide (t-BHP), an analog of hydroperoxide, induced premature senescence model in human WI-38 diploid fibroblast cells (Chen et al., 2008). We had learned that the expressions of p16 and p21 proteins increased with t-BHP incubation by reading abundant literatures. So, we used the t-BHP stimulation to establish the model of NSCs senescence.
Osthole (7-methoxy-8-isopentenyl coumarin, C15H16O3, 244.38 Da, Ost.) is a natural coumarin isolated from many kinds of herbs such as Cnidiummonnieri (L.), which has great medical value, and is also widely used in Asia (Gao et al., 2014). In recent decades, osthole was reported to have diverse pharmacological functions, such that it could improve cognitive impairments and BBB dysfunction after transient global brain ischemia in C57BL/6J mice (Chen et al., 2015), and it had neuroprotective effects of traumatic brain injury in rats (He et al., 2012), it also could protect the memory impairment of mice in AlCl3-induced acute senile model (Shen et al., 2002).
Our previous studies implied that osthole confer neuroprotection against cortical stab wound injury and attenuate secondary brain injury (Xia et al., 2015), it also could reverse Aβ peptide cytotoxicity on neural cells and promote the ability of proliferation and differentiation of NSCs in AD model (Hu et al., 2013; Yao et al., 2015). So, in this research, we found that osthole potently enhanced NSCs proliferation and neuronal differentiation, and had antiaging in t-BHP-induced NSCs senescence cell model in vitro through p16-pRb signaling pathway.
Materials and Methods
Preparation of osthole
Osthole (No.110822-200305, purity >98%, 244.39 Da; Fig. 1) was purchased from the National Institute for the Control of Pharmaceutical and Biological products (Beijing, China). Osthole was dissolved in dimethylsulfoxide (DMSO), in which the largest proportion was 0.1%. Stock solution of osthole (1 mM) was dissolved in phosphate-buffered saline (PBS) and stored at −20°C (Zhang et al., 2012).
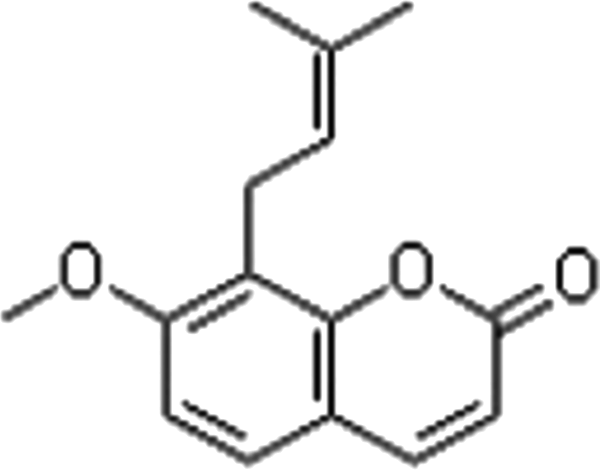
The chemical structure of osthole.
Neural stem cells culture
NSCs were isolated from hippocampus region of newborn mice (Van et al., 2002), which were born within 2 days. The brain tissue was cut into about 2 mm2 diamond and digested by trypsin (Sigma). Shortly, cells were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 (1:1; Gibco) medium supplemented with 1% B27 (Invitrogen), 20 ng/mL of basic fibroblast growth factor (bFGF; PeproTech), 20 ng/mL of epidermal growth factor (EGF; PeproTech), and 100 U/mL penicillin and phytomycin (Sigma). They were seeded at a density of 1 × 106 viable cells/mL in the 24-well plates (Corning), then the cultures were incubated at 37°C in the humidified atmosphere of 95% air and 5% CO2. The culture medium was replaced after 3 days. Subsequently, the cells began to grow into floating neurospheres. After about 7 days in vitro, primary neurospheres were dissociated into single cells and reseeded in 24-well plates.
Immunochemistry
NSCs were divided into three groups: (1) control group, (2) t-BHP group (100 μM, 2 hours), and (3) t-BHP+osthole group (t-BHP: 100 μM, 2 hours+ost: 100 μM, 2 hours).
The cells in 96-well plates were fixed in 4% paraformaldehyde for 30 minutes at 4°C, washed three times with PBS, and permeabilized with 1% Triton X-100 for 30 minutes at room temperature (Hamamoto et al., 2004). Then, we used 3% BSA to incubate the NSCs again for 30 minutes. The primary antibodies were used to incubate the cells at 4°C overnight: mouse anti-Nestin, rabbit anti-Sox2, anti-GFAP, rabbit mouse anti-NeuN, anti-NG2, rabbit anti-Ki67, and corresponding following antibodies (FITC-conjugated goat anti-mouse IgG, Cy™3 donkey anti-rabbit IgG; Jackson) were incubated for 1 hour at the next morning. We used 4′6-diamidoino-2-phenylindole (DAPI, 1:100 dilutions; Sigma) to stain the nuclear. The number of immunoreactive cells in each well was counted by using fluorescent microscope.
CCK-8 assay
NSCs viability in different group was determined by using the Cell Counting Kit-8 assay (CCK-8; Dojindo Laboratories, Japan) (Seo et al., 2013), which was a colorimetric kit for simple and accurate cell proliferation and cytotoxicity assays. The dissociated cells were incubated in 96-well plates at a density of 5 × 104 cells/well with 100 μL complete medium per well in triplicate for cultivating for 48 hours. Then, 10 μL CCK-8 solution was added to each well according to the manufacturer's instruction, NSCs were incubated at 37°C for 4 hours in the humidified atmosphere of 95% air and 5% CO2. The absorbance value was measured at wavelength of 450 nm using a microplate reader (MR-96A; Mindray, China), the absorbance value was highly correlated with the number of living NSCs.
SA-β-gal staining
SA-β-gal assay (KeyGEN BioTech, Nanjing, China) was stained to determine of cell senescence (Warrier et al., 2013). The NSCs were cultured in six-well plates for 2 days. Once NSCs reached 80–90% confluence, we used t-BHP to cause cells senescence. Then, we used osthole to treat for 2 hours. Next, the plates were fixed in 4% paraformaldehyde for 30 minutes at 4°C. After washing with PBS, they were incubated in 1 mL fixative SA-β-gal solution (fixed with 10 μL SA-β-gal staining solution A, 10 μL SA-β-gal staining solution B, 930 μL SA-β-gal staining solution C, and 50 μL X-Gal solution) at 37°C overnight. The second morning, plates were checked for blue coloration by optical microscope. An average percentage of SA-β-gal positive cells was obtained from three independent experiment.
Cell cycle analysis
Cell cycle analysis was determined by flow cytometry, the cells were fixed by 75% ethanol overnight at 4°C, and were washed by PBS. And NSCs were exposed to propidium iodide (PI) solution (PI at 50 μg/mL and RNase A at 1 mg/mL) for 30 minutes at 4°C in the dark (Yao et al., 2014). After being washed with PBS, the NSCs were measured by FACScan Flow Cytometry System at λ = 488 nm (Becton Dickinson).
Reverse transcription (RT)-polymerase chain reaction quantification of messenger RNA
Total RNA from NSCs with t-BHP and osthole treatment was isolated by adding 1 mL TRIzol reagent, according to the manufacturer's instructions (Invitrogen), then using RevertAid™ First-Strand cDNA Synthesis Kit (Thermo) to reverse transcribe into cDNA. We used the primers that gave efficiency close to 100%. The primers are as follows: p16-F (5′-ATG ATG GGC AAC GTT CAC GTA-3′), p16-R (5′-AAC GCA AAT ATC GCA CGA TGT-3′); β-actin-F (5′-GGG AAA TCG TGC GTG ACA T-3′); β-actin-R (5′-TCA GGA GGA GCA ATG ATC TTG-3′) (Invitrogen). The polymerase chain reaction (PCR) was for 35 cycles using a DreamTaq™ Green PCR Master Mix Kit (Thermo) (Marwarha et al., 2014). The banks were quantified using Image J to reflect the original messenger RNA (mRNA) level.
Western blotting
NSCs were washed with cold PBS and added the protein inhibitors, according to the manufacturer's instruction of total protein extraction kit (KeyGEN BioTech) (Trazzi et al., 2011), then protein concentration was estimated by the Braford method. Hundred microgram proteins were subjected to electrophoresis on an 8% sodium dodecyl sulfate—polyacrylamide gel and transferred to polyvinylidene difluoride membranes (PDVF; Thermo Fisher Scientific) (Li et al., 2017). The membranes were incubated with primary antibodies followed by appropriate secondary antibodies marked with horseradish peroxidase, and we used the method of diaminobezidin (DAB) to stain the PDVF membranes. The banks were quantified using Image J to reflect the proteins level.
Statistical analysis
All data are expressed as the mean ± SEM of three separate experiments. Differences between groups were determined using a one-way analysis of variance (ANOVA) using GraphPad Prism 5 (GraphPad Software). Student's test was used to compare two groups. *p < 0.05, **p < 0.01 were considered to be regarded as significance.
Results
Generation and characterization of NSCs in vitro
To generate NSCs, the whole hippocampus was harvested from newborn mice, and neurospheres were generated as described as section 2.2. After about 10 days in culture, individual cells proliferated to form neurospheres containing about 150 NSCs. Nestin and Sox2 staining were used to identify the neurospheres, which were the hallmark of NSCs (Yao et al., 2015). We found that more than about 90% of single NSCs were positive for Nestin and Sox2 in Figure 2A.

Generation and characterization of NSCs in vitro.
Osthole enhanced the viability of NSCs
To determine whether or not osthole had effects on the survival of NSCs, single NSCs at fourth passage were treated with t-BHP (100 μM) for 2 hours, and then, we used only osthole (100 μM) in growth medium for 2 hours (Yang et al., 2010). The viability of NSCs was tested by CCK-8 assay, the result showed that the viability of NSCs declined by 37.28% in t-BHP group, compared with control group. And the viability of NSCs increased by 14.29% in osthole group, compared with t-BHP group in Figure 3.
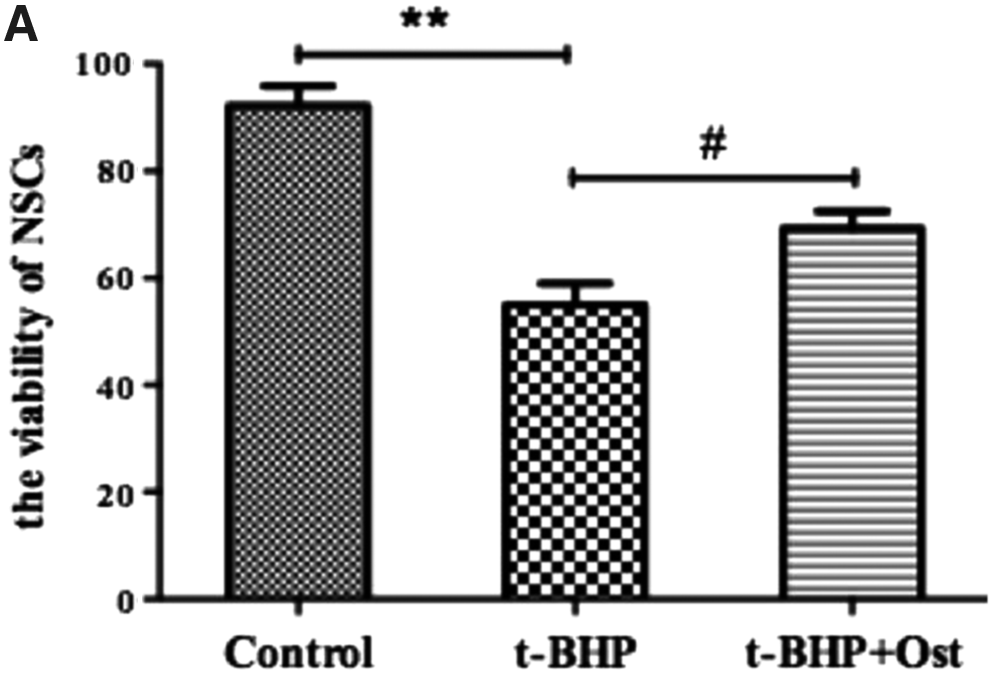
Osthole enhanced the viability of NSCs.
Osthole enhanced the ability of proliferation in NSCs
To determine the effects of osthole on NSCs proliferation, single NSCs at fourth passage was treated with t-BHP and osthole in growth medium. The proliferation potential of NSCs was also tested by in vitro Ki67 labeling in Figure 4A (Schonk et al., 1989). After treatment of t-BHP in 2 hours to establish a model of NSCs senescence, cells were incubated with osthole treatment for 2 hours. The results showed that significantly lower levels of Ki67-positive cells were found in t-BHP group compared with control group in Figure 4B (44.78% ± 3.51% vs. 78.12% ± 5.57%, #p < 0.05). We found that the Ki67 positive cells increased about 24.46% with osthole treatment, when compared with t-BHP group.

The effects of the ability of proliferation in NSCs.
Osthole enhanced the ability of differentiation in NSCs
To determine the effects of osthole on differentiation of NSCs in vitro, single NSCs dissociated from the spheres were cultivated in differentiation medium (DMEM-F12 + 10% FBS +1% P/S) with or without osthole. After about 10 days, NSCs were developed and differentiated into astrocytes (GFAP+), neurons (NeuN+), and oligodendrocyte precursors (NG2+), as verified by immune staining in Figure 5A. Quantitative analysis showed that osthole promoted NSCs differentiating into greater numbers of neurons by 8.35% and astrocytes by 10.78%, it also promoted the ability of differentiation into oligodendrocyte precursors, but had no significant difference in osthole group, compared with t-BHP group in Figure 5B.

The effect of the ability of differentiation in NSCs.
Osthole reduced the percentage of SA-β-gal+ NSCs
To test whether t-BHP treatment induced cells senescence in NSCs, we stained NSCs with senescence-associated-β-galactosidase (SA-β-gal), which is an acknowledged senescence marker in Figure 6A. Then t-BHP treatment induced the expression of SA-β-gal+ cells. The percentage of SA-β-gal+ NSCs increased from 6.53% in control group to 68.55% in t-BHP group, and the percentage of SA-β-gal+ NSCs induced by 22.14% in osthole group, compared with t-BHP group in Figure 6B.
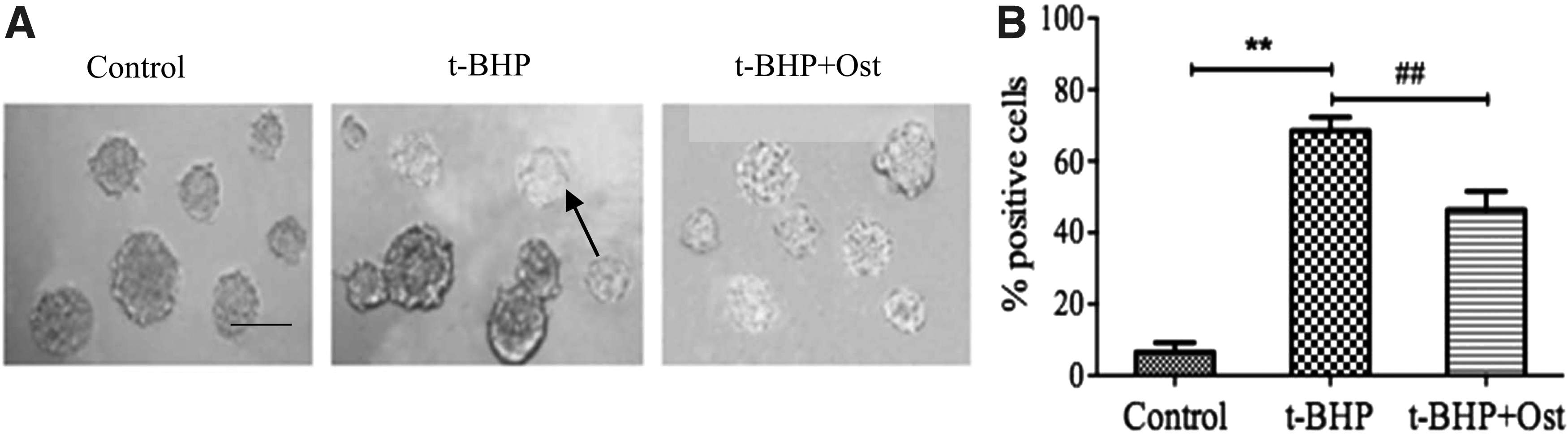
Senescence-associated beta-galactosidase (SA-β-gal) stains in NSCs.
Osthole decreased the proportion of G0 and G1 phase in senescence NSCs
To investigate how osthole effected NSCs cycle (Zhang et al., 2014), the cell cycle distribution was analyzed by FACScan flow cytometry in Figure 7A. The proportion of cells at G1 phase in t-BHP group was about 76.18% ± 6.50% (**p < 0.01), and declined by 17.22% (##p < 0.01) in osthole group in Figure 7B. We found that osthole could decrease the first peak in mixture of G0 and G1, compared with t-BHP group.

Osthole protected NSCs senescence against G1 arrest.
Osthole had effects on the levels of target gene and proteins in the t-BHP-induced NSCs
To observe that osthole could reverse t-BHP-induced NSCs senescence effectively, afterward we investigated the molecular mechanism underlying this effect. For this purpose, we detected the expression of p16 mRNA in Figure 8A. The result showed that osthole could reserve the enhancement of p16 mRNA expression in t-BHP treatment about 24.74% in Figure 8B (71.97% ± 4.66% vs. 47.23% ± 4.35%).
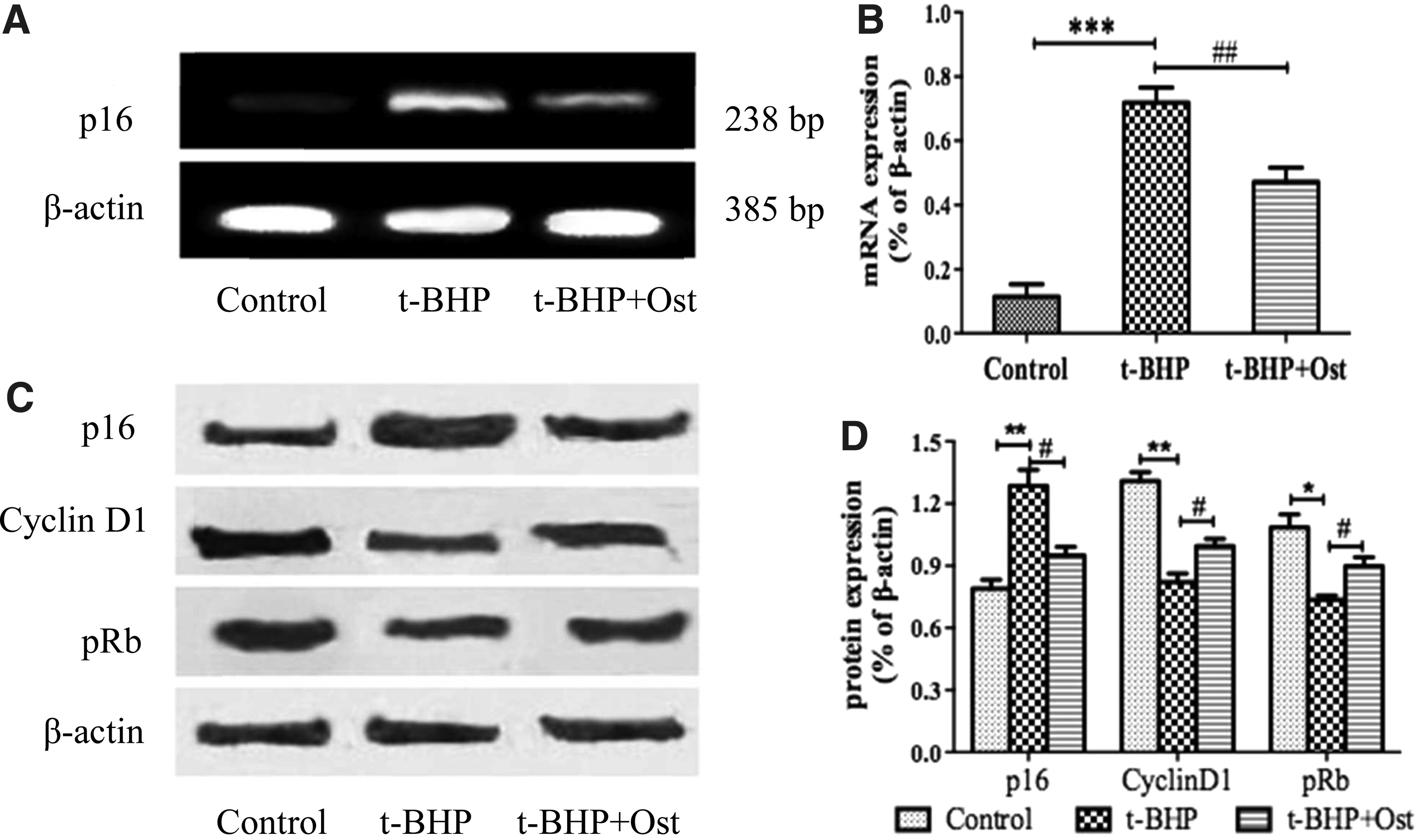
The expressions of mRNA and proteins in p16-pRb signaling pathway.
Then we tested the proteins of p16-pRb signaling pathway in Figure 8C, and the results showed that the levels of p16 protein increased significantly in t-BHP group, compared with control group (**p < 0.01) and declined by 33.5%. Finally, the results showed that in response to t-BHP treatment, cyclinD1 and pRb were downregulated. Furthermore, osthole could reverse the expression of these proteins to some extent and had a better effect when treated with t-BHP in Figure 8D (**p < 0.01, *p < 0.05 and #p < 0.01).
Discussion
Aging and aging-related disease seems to be a major social and economic burden for the modern society. Cellular senescence refers to a complex cellular response to a variety of stressors, which would lead to a gradual loss of their renewal and multipotential differentiation ability, also could cause abnormal proliferation and differentiation (Beltrami et al., 2012). NSCs senescence would result in severe disorders in the structure or function of tissue; it even could accelerate disease generation and development.
T-BHP, a reactive hydroperoxide produced from lipid peroxidation, is widely used in research related to senescence both in vitro and in vivo (Maurya and Rizvi, 2009; Rossi et al., 2007). In this study, we at first established the model of NSCs premature senescence in vitro. The CCK-8 assay results showed the viability of NSCs increased by 14.29% with osthole treatment. Then, we examined the ability of proliferation and differentiation by immunology staining. It is not difficult to find Ki67+ cells in t-BHP group declined by 33.34%, compared with control group, but with osthole treatment, it increased by 24.46%. It indicated that osthole could promote the ability of proliferation in NSCs senescence. Subsequently, we found the ability of differentiation into greater numbers of neurons and astrocytes with osthole treatment, compared with t-BHP group.
Then we examined the antisenescence effects of osthole by analyzing the SA-β-gal expression, because its expression can be characterized as special marker of cellular senescence (Lee et al., 1995). We found that osthole treatment resulted in the most pronounced reversal of t-BHP-induced NSCs senescence, it would suggest that osthole might have an antisenescence effect. From above results, we knew that osthole could promote the viability of NSCs and the ability of proliferation; we then tested the cell cycle by FACScan flow cytometry and found that the proportion of G0 and G1 phase in t-BHP group was significantly higher, but osthole could decrease its proportion.
The cell cycle and senescence are closely linked through some specific signaling pathways, and it is well known that p16INK4a-pRb and p53-p21Cip1/Waf1-pRb are the signaling pathways (DiMaio et al., 2006; Janzen et al., 2006). According to previous results, to investigate the molecular mechanism of osthole pharmacologic effects, we selected p16INK4a-pRb signaling pathway.
p16INK4a could prevent CDK4/6 from combination with cyclin D1 to inhibit pRb′ phosphorylation, which could conversely inhibit E2F release, and finally increased the proportion of G0 and G1 phase (Molofsky et al., 2006; Noda et al., 2009). Then we found that the mRNA and protein expression levels of p16INK4a increased significantly, however, the expression of its downstream proteins such as cyclinD1 and pRb declined in t-BHP group, but it had opposite expressions of them in osthole group. Therefore, we found osthole could delay t-BHP-induced NSCs premature senescence, and its mechanism might be related with suppressing p16INK4a-pRb signaling pathway.
Moreover, in conclusion, osthole had significant effects on NSCs senescence; in this study, it also provided evidence that if p16INK4a-pRb signaling pathway was suppressed, it might be potential for elucidating the molecular mechanism underlying the osthole antisenescence effect. It suggested that osthole may act as a potential and therapeutic target drug in treatment of aging-related diseases or other degenerative diseases.
Footnotes
Acknowledgments
This project was funded by the National Natural Science Foundation of China (No. 81173580), the Natural Science Foundation of Liaoning Province (No. 20170540616) and the Youth Project of Liaoning Education Department (No. L201712)
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.