
Abstract
Abstract
The forkhead box C1 (Foxc1) protein, a member of the forkhead/winged helix transcription factor family, is required in stem cell developmental processes. Recently, multiple studies have indicated the crucial role of Foxc1 in mesenchymal stem cell differentiation, but the precise effects and mechanisms on dental pulp stem cells (DPSCs) remain unclear. In this study, we evaluate the role of Foxc1 on the odontogenic differentiation and proliferation of DPSCs. Our results show that Foxc1 decreases time dependently in odontogenic differentiation of DPSCs. Meanwhile, overexpression of Foxc1 could significantly inhibit the mineralization of DPSCs and the expression of odontogenic-related genes, such as runt-related transcription factor 2 (Runx2), dentin sialophosphoprote (DSPP), and dentin matrix acidic phosphoprotein 1 (DMP-1). Foxc1 overexpression does not significantly alter the proliferation of DPSCs. In addition, Foxc1 reduces the expression of p-Smad1/5, an important modulator of bone morphogenetic protein (BMP)/Smad signaling pathway, inhibiting BMP/Smad signaling pathway. In conclusion, our data demonstrated that Foxc1 inhibits odontogenic differentiation of DPSCs and odontogenic-related gene expression through the BMP/Smad signaling pathway which may be useful for the dental regeneration and repair.
Introduction
D
DPSCs have been a focus of research in stem cell-based tissue engineering during the recent years. Numerous studies have shown that DPSCs are highly accessible and multipotent and can differentiate into various cells, such as osteoblasts, odontoblasts, chondrocytes, endothelial cells, nerve cells, and adipocytes (Gronthos et al., 2002; Lee et al., 2016; Liu et al., 2018; Nakashima et al., 2009; Zhang et al., 2017). It is shown that DPSCs have the potential to promote dentinogenesis, which makes them become an ideal cell type in engineering tissue (Lian et al., 2016). Consequently, detection of the molecular responsible signal in the process of DPSC odontogenic differentiation may provide a valuable tool for regeneration and self-repairing capability of these cells.
Forkhead box C1 (Foxc1) is a transcriptional regulatory protein belonging to a large forkhead box or FOX genes which were evolutionarily conserved (Benayoun et al., 2011; Lam et al., 2013). These genes encode proteins characterized by a common 100-amino acid winged-helix DNA-binding domain as the forkhead domain (Omatsu et al., 2014). It has been reported that Foxc1 has important functional diversity and participates in a wide spectrum of cellular processes, such as cell cycle progression, differentiation, proliferation, survival, apoptosis, migration, and DNA damage response (Carlsson et al., 2002; Wang et al., 2012; Yoshida et al., 2015).
Foxc1 mutant mice result in a dramatic reduction in vertebrae, ribs, and skull vault growth, indicating an important role of the gene in the normal development of skeleton in mice (Machida et al., 2014; Rice et al., 2005; Sun et al., 2013). A growth of evidence indicates that the inhibited expression of Foxc1 can increase the expression of early markers of osteogenic differentiation in mesenchymal stem cells (MSCs) (Hopkins et al., 2016). However, the role of Foxc1 in regulating odontogenic differentiation of DPSCs remains unclear.
Bone morphogenetic proteins (BMPs) are members of the transforming growth factor-β. Their osteogenic activities have been demonstrated in numerous clinical and animal studies (Feng et al., 2017; Katagiri et al., 1994; Yoshida et al., 2000). BMP/Smad signaling, which is an extensively studied pathway relevant directly to bone formation, has been identified for controlling osteogenesis in the developing bone structure (Chen et al., 2012; Yoshida et al., 2000). Meanwhile, research has shown that BMP/Smad signaling pathway plays an essential role in osteogenesis and adipogenesis of MSCs (Lee et al., 2010; Liu et al., 2014).
Recently, many studies also have shown that dental pulp development is regulated by BMP/Smad signaling (Casagrande et al., 2010; Wang et al., 2014). Previous research shows that there are functional links between Foxc1 and BMP/Smad signaling (Hopkins et al., 2016; Rice et al., 2005). The Foxc1 promoter in direct interactions with Smad protein and Foxc1 expression was regulated by BMP/Smad signaling during osteogenic differentiation in C2C12 myoblast cells (Hopkins et al., 2016). Thus, we attempted to verify whether Foxc1 expression was related to BMP/Smad signaling during odontogenic differentiation of DPSCs.
In this study, we mainly investigated the effect of Foxc1 on the progression of odontogenic differentiation of DPSCs and the BMP/Smad pathway. This study was designed to determine how Foxc1 interacts with other associated signaling pathways to control DPSC odontogenic differentiation functions. We first find that overexpression Foxc1 can inhibit odontogenic differentiation of DPSCs by inhibiting the BMP/Smad signaling. Our finding may provide a foundation for odontogenic differentiation in the process of DPSCs promoting dentinogenesis.
Materials and Methods
Isolation, culture, and chemical treatment of DPSCs
As previously described, the clonal populations of DPSCs were isolated (Feng et al., 2013). Concisely, the dental pulp tissues were obtained from the third molars of healthy patients (donors aged from 18 to 26 years) at the Dental Clinic of Nantong University. This study was agreed by the Ethics Committee of the Affiliated Hospital of Nantong University.
The subjects included in the study were free of carious lesions and oral infections. All the methods in the study were carried out in accordance with the approved guidelines. Tooth surfaces were cleaned and cracked openly using a bone cutter to reveal the pulp chamber. Pulp tissues were digested with 4 mg/mL solution of collagenase/dispase for 1 hour at 37°C and then mixed with the centrifugation and resuspension 4 mL phosphate-buffered saline (PBS) and the 100 μM
Cultures were incubated at 37°C in 5% CO2 and the medium changed every 3 days. The clonal populations of DPSCs were isolated using a limiting dilution protocol, and cells at the third or fourth passages were used for subsequent experiments (Soukup et al., 2006; Suchanek et al., 2007).
Odontogenic differentiation
A total of 4 × 104 cells/cm2 of DPSCs were plated cultured in the growth medium to undergo differentiation stimulation. To induce odontogenic differentiation, after reaching 80% confluence, DPSCs were cultured in odontogenic medium containing DMEM, 10% FBS (Biochrom), 0.1 μM dexamethasone (Sigma), 10 mM β-glycerophosphate (Sigma), and 50 μg/mL ascorbic acid (Sigma). The medium changed every 3 days.
Cell proliferation analysis using MTT
DPSCs infected with retrovirus containing blank vector (Control) and overexpressing Foxc1 (oeFoxc1) were seeded into 96-well plates at concentration of 4 × 103 cells/well and cultured for 14 days. Proliferation of the cells was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma). The cells were washed with PBS, and 20 mL of MTT (5 g/L; Beyotime, Shanghai, China) was added to each well. After 4 hours of incubation at 37°C with MTT, the precipitate was extracted with dimethyl sulfoxide (Sigma), and the optical density (OD) was determined at the wavelength of 490 nm.
Western blot analysis
DPSCs were washed with cold PBS (pH 7.0) and lysed at 4°C in lysis buffer (50 mM Tris–HCl [pH 7.4], 150 mM NaCl, 20 mM EDTA, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], and a protease inhibitor mixture). After centrifugation at 12,000 rpm for 12 minutes, protein concentrations were determined using the Bradford assay (Bio-Rad, CA). The resulting supernatant (50 μg of protein) was subjected to SDS-polyacrylamide gel electrophoresis.
The separated proteins were transferred to polyvinylidene fluoride membranes at 350 mA for 2.5 hours in a blotting apparatus (Bio-Rad). Membranes were blocked with 5% nonfat milk in TBST (Tris-buffered saline plus 0.1% Tween 20), and the membranes were immunoblotted with various antibodies (1:1000) at 4°C overnight and subsequently with the corresponding secondary antibodies (1:1000) for 2 hours at room temperature. Concomitantly, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was run as a reference protein.
The following primary antibodies were used: GAPDH (anti-rabbit; Santa Cruz), Foxc1 (anti-rabbit; Abcam), runt-related transcription factor 2 (Runx2, anti-rabbit; Santa Cruz), dentin sialophosphoprote (DSPP, anti-rabbit; Santa Cruz), dentin matrix acidic phosphoprotein 1 (DMP-1, anti-rabbit; Santa Cruz), p-Smad1/5 (anti-rabbit; Santa Cruz), and Smad1/5/8 (anti-rabbit; Santa Cruz).
Immunofluorescent staining
DPSCs were fixed with methanol, permeabilized with 1% NP-40, blocked with 10% BSA, and washed with PBS containing 0.1% Triton X-100. The cells were incubated with one of the following first antibodies overnight at 4°C: Foxc1 (anti-rabbit; Abcam). After being washed, cells were incubated with Texas Red-labeled secondary antibody for 2 hours at room temperature. Nuclei were stained with DAPI (4, 6-diamidino-2-phenylindole dihydrochloride) (1:800; Santa Cruz). After being washed and mounted, images were obtained using a fluorescence microscope (Germany).
Lentiviral production and transfection
A lentiviral vector expressing human Foxc1 and a blank vector were purchased from Santa Cruz. Establishment of Foxc1 overexpressed (oeFoxc1) DPSCs as experimental group. DPSCs infected by retrovirus containing blank vector (Control) were used as the control group in the following detections. Cells were used for the following experiments 72 hours after transfection.
Reverse transcription and real-time polymerase chain reaction analysis
Total RNA was extracted from induced DPSCs for 14 days. Total cellular RNA was isolated from cells and reversely transcribed using the QuantiTect Reverse Transcription Kit to obtain complementary DNAs (cDNAs). Polymerase chain reaction (PCR) cycling program was initial denaturation at 95°C for 10 minutes, denaturation at 95°C for 10 seconds, and extension at 60°C for 30 seconds. The target gene expression was normalized relative to the level of β-actin. The primers for GAPDH, Foxc1, Runx2, DSPP, and DMP-1 are listed in Table 1.
All the primer sequences were determined using established GenBank sequences.
DMP-1, dentin matrix acidic phosphoprotein 1; DSPP, dentin sialophosphoprote; Foxc1, forkhead box C1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Runx2, runt-related transcription factor 2.
Alizarin red S staining
DPSCs have grown in odontogenic differentiation medium for 14 days and were fixed with 4% paraformaldehyde for 1 hour and washed twice with PBS. Alizarin red S solution (Sigma) was applied, and DPSCs were placed in an incubator at 37°C for 2 hours. Mineralization was quantified by extracting the Alizarin red S stain with 100 mM cetylpyridinium chloride (Sigma) at room temperature for 2 hours. Absorbance of the extracted Alizarin red S stain was measured at 570 nm.
Statistical analysis
The results were analyzed by the mean ± standard deviation, and the statistical significance was estimated using SPSS v17.0. The significance of differences between the experimental groups and the controls was analyzed by the independent samples t-test and ANOVA test. Values of p < 0.05 was considered statistically significant.
Results
Related gene expression during odontogenic differentiation of DPSCs
To determine the stage of odontogenic differentiation in vitro, DPSCs were cultured in differentiation medium for 0, 3, 5, 7, and 14 days, and the expression levels of odontogenic differentiation markers were analyzed by western blot and real-time PCR. The formation of mineralized nodules was evaluated by Alizarin red staining. Alizarin red S staining revealed the presence of mineralized nodules of 3, 5, 7 to 14 days and increased in time-dependent manners under differentiation medium (Fig. 1A, B). Western blot also showed that the protein expression of Runx2, DSPP, and DMP-1 was increased time dependently in the DPSCs (Fig. 1C, D). Runx2, DSPP, and DMP-1 mRNA were also increased time dependently (Fig. 1E). This suggests that the odontogenic differentiation of DPSCs increased with time.
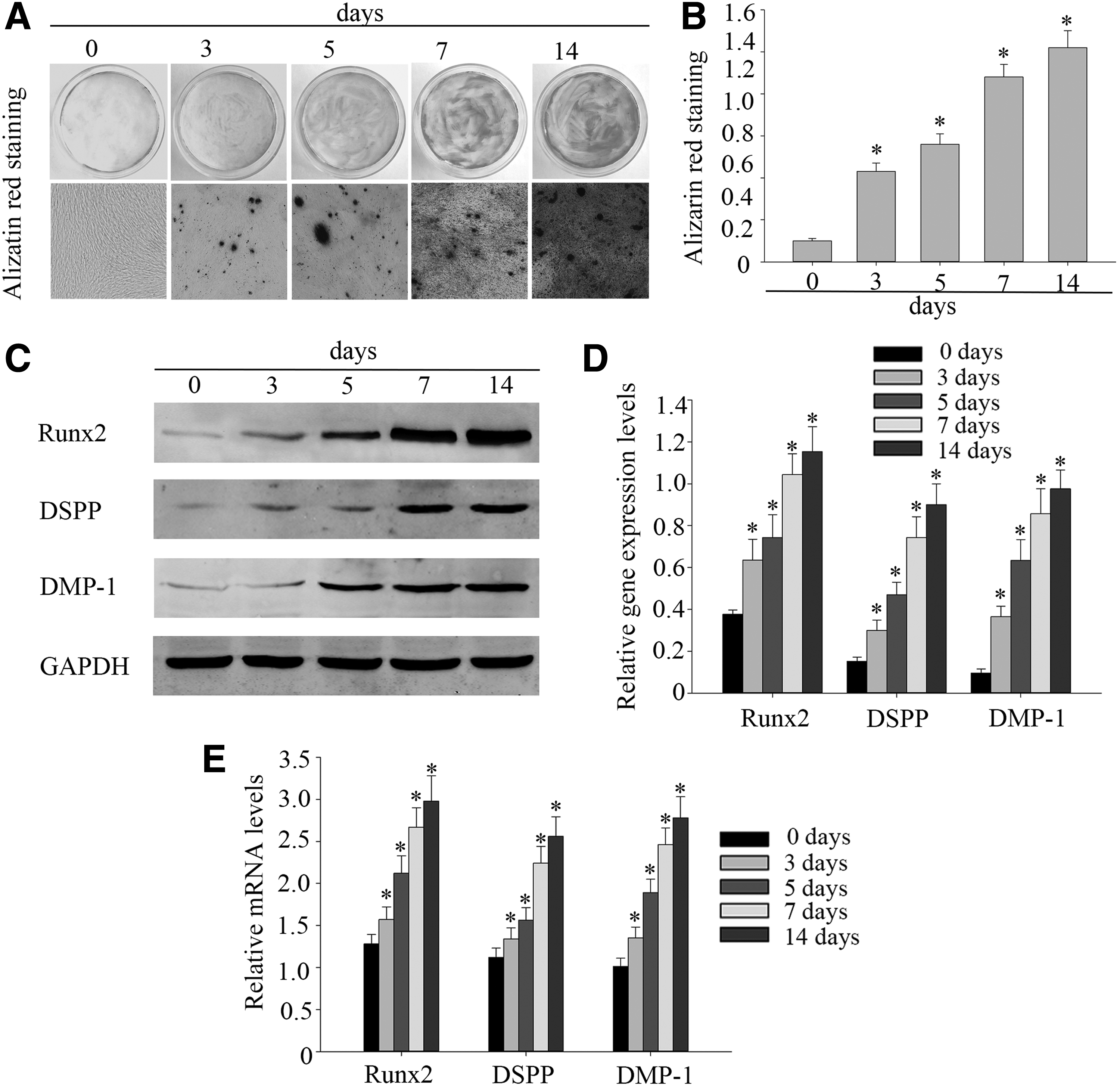
Related gene expression during odontogenic differentiation of DPSCs. DPSCs were cultured in odontogenic differentiation medium for up to 14 days.
Expression of Foxc1 is downregulated during odontogenic differentiation of DPSCs
DPSCs were cultured in differentiation medium for 0, 3, 5, 7, and 14 days. During in vitro odontogenic differentiation, we noted the Foxc1 protein levels which decreased time dependently (Fig. 2A, B). The mRNA changes of Foxc1 were similar to those observed in the protein levels (Fig. 2C). Furthermore, the Foxc1 expression was confirmed using immunofluorescence staining. The downregulation of Foxc1 was observed on the 14th day of induced DPSCs (Fig. 2D). The above results showed that the expression of Foxc1 was downregulated during odontogenic differentiation of DPSCs.
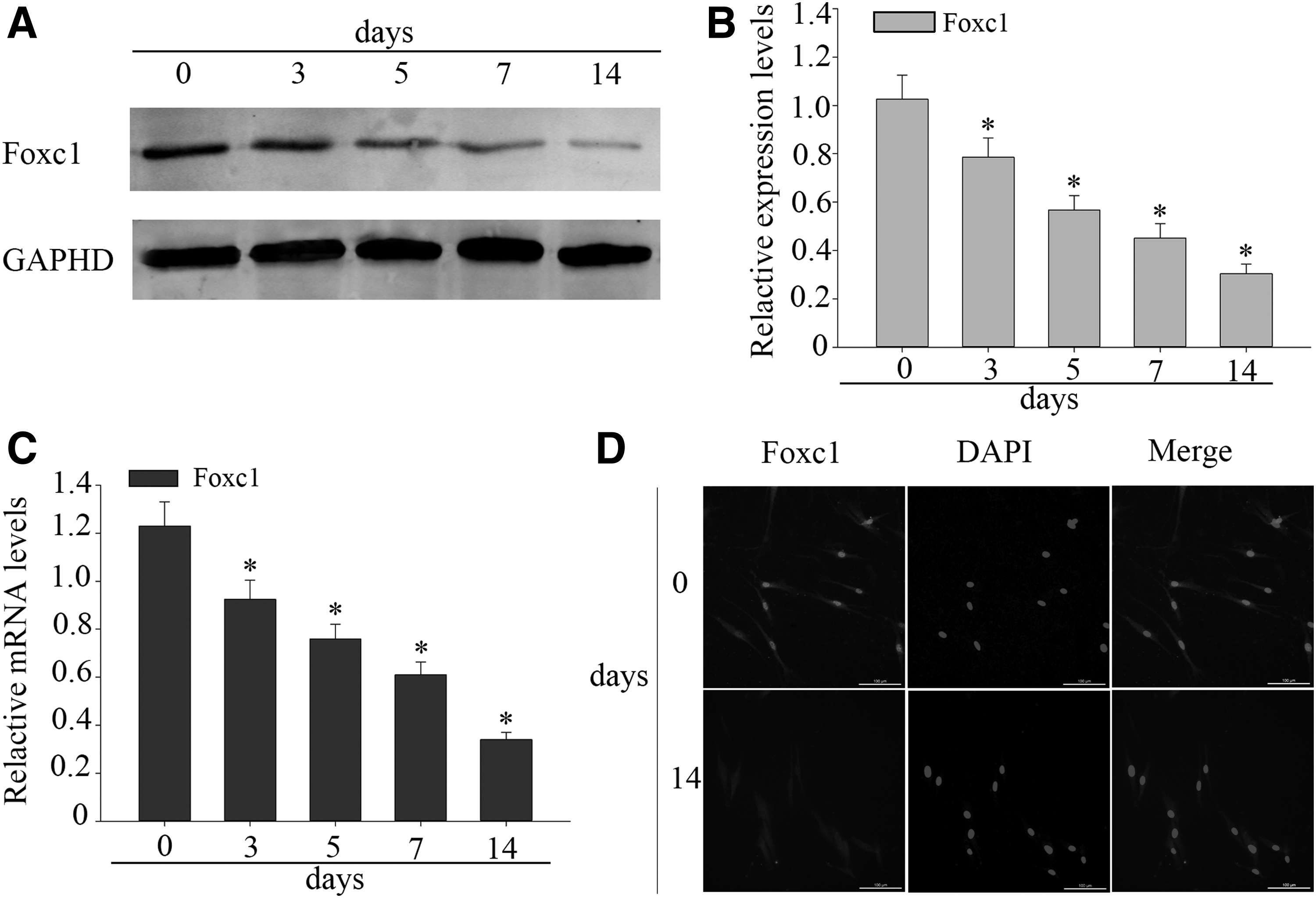
Expression of Foxc1 is downregulated during odontogenic differentiation of DPSCs.
Foxc1 is stably expressed and no effect to proliferation of DPSCs
To determine the biological effect of Foxc1 in cell viability, the same number of DPSCs was seeded in 24-well plates 72 hours after transduction, and cell viability was detected. The OD values of untransduced DPSCs (control group) were taken as 100%, and percentages of cell viability were then obtained. As illustrated, cell viability increased significantly in DPSCs transduced with Foxc1 (oeFoxc1 group) (123.8% ± 1.2%) (Fig. 3A). To determine the effect of Foxc1 on DPSC proliferation and differentiation, we transfected a full-length Foxc1 cDNA into DPSCs. When DPSCs grew to 80% confluence, the cells were transfected with Foxc1 for 72 hours. The expression of Foxc1 increased and reached its peak on the 7th day post-transduction and maintained high expression during the following 14 days (Fig. 3B).
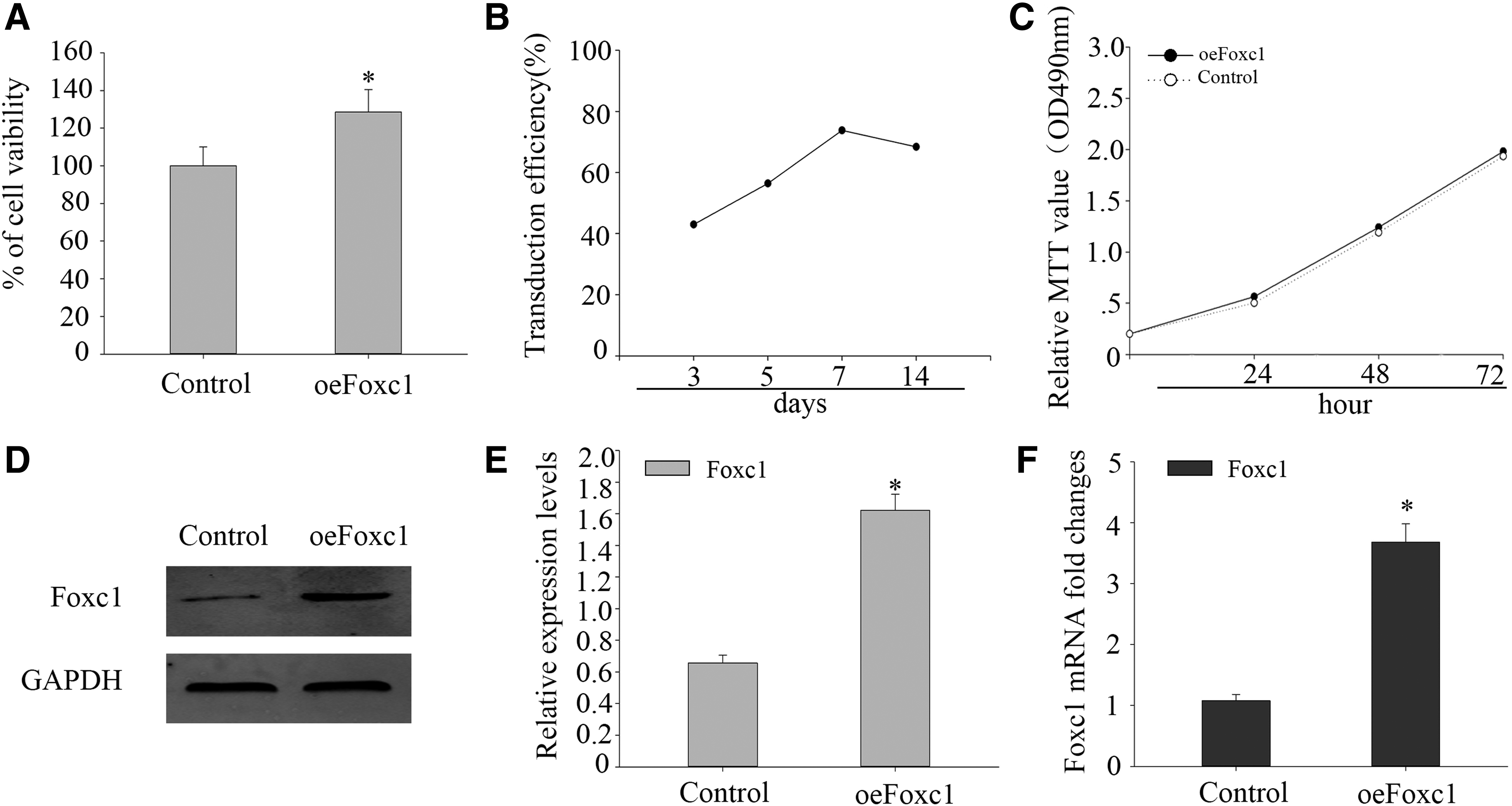
Foxc1 is stably expressed and no effect to proliferation in DPSCs.
There was a significant statistical difference in Foxc1 expression between the oeFoxc1 group and control group. Next, MTT assay was used to reveal the proliferation of DPSCs. The MTT assay data indicated that no obvious change can be seen in the proliferation between oeFoxc1 group and control group (Fig. 3C). Western blotting and real-time PCR showed that Foxc1 gene was stably expressed in Foxc1-transduced DPSCs; Foxc1 expression was significantly higher in oeFoxc1 group (Fig. 3D–F).
Foxc1 overexpression suppressed BMP/Smad signaling activity in DPSCs
To examine the expression of BMP/Smad signaling on odontogenic differentiation of DPSCs, we examined the expression levels of Smad1/5/8 and p-Smad1/5. As is shown in Figure 4, there is no obvious change in the expression of Smad1/5/8; however, the expression levels of p-Smad1/5 increased in time-dependent manners (Fig. 4A, B). These results indicated that the BMP/Smad pathway may manipulate the odontogenic differentiation of DPSCs. To examine the role of Foxc1 in regulating the BMP/Smad signaling pathway in DPSCs, the cells were transduced with Foxc1 or not and cultivated for 14 days under odontogenic conditions.
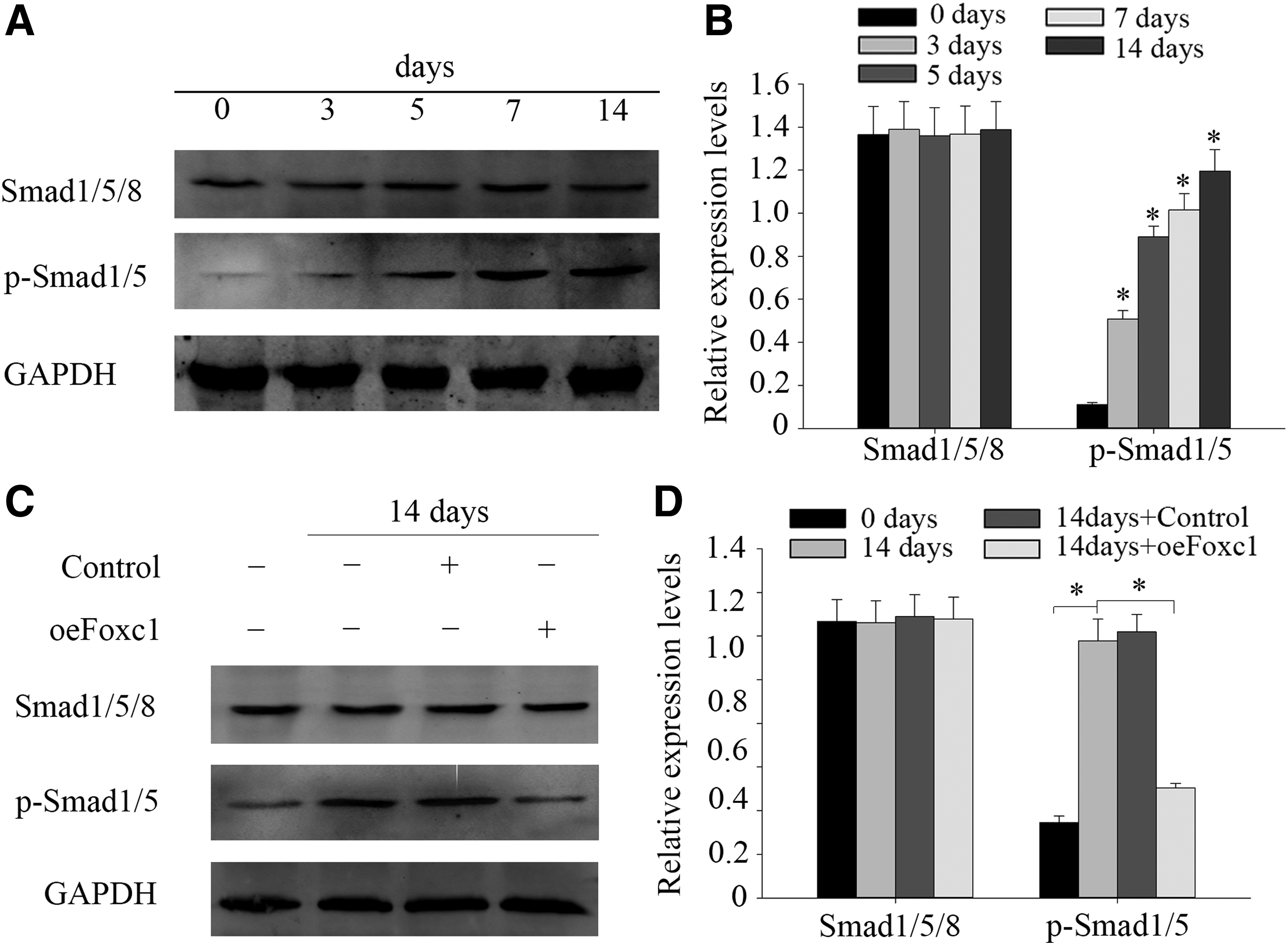
Foxc1 overexpression suppressed BMP/Smad signaling activity in odontogenic differentiation DPSCs. DPSCs were cultured in odontogenic differentiation medium for up to 14 days.
Western blot was performed to see whether Foxc1 overexpression affected the Smad1/5/8 and p-Smad1/5 levels in DPSCs or not. Smad1/5/8 protein expression was not altered with the untransduced group and the overexpression Foxc1 group. Meanwhile, results showed that the p-Smad1/5 expression in cells treated with Foxc1 decreased after 14 days' transduction, while the protein rarely changed in untransduced cells (Fig. 4C, D). These results demonstrated that Foxc1 overexpression suppressed the BMP/Smad signaling activity in DPSCs.
Overexpression of Foxc1 inhibits odontogenic differentiation in DPSCs
To examine the role of Foxc1 in regulating the odontogenic differentiation in DPSCs, the cells were transduced with Foxc1 under odontogenic conditions for 14 days. Results of western blot showed that the odontogenic markers Runx2, DSPP, and DMP-1 were highly expressed in normal cells and untransduced cells while rarely expressed in Foxc1 transfected cells after 14 days (Fig. 5A, B). This suggested that Foxc1 overexpression might provide the transduced cells an inhibited odontogenic differentiation inclination. With Foxc1 overexpression, we observed a detraction in Alizarin red S staining compared with the normal group and oeCon group after 14 days of culture in odontogenic differentiation medium (Fig. 5C, D). The above results showed that overexpression of Foxc1 inhibited the odontogenic differentiation in DPSCs.
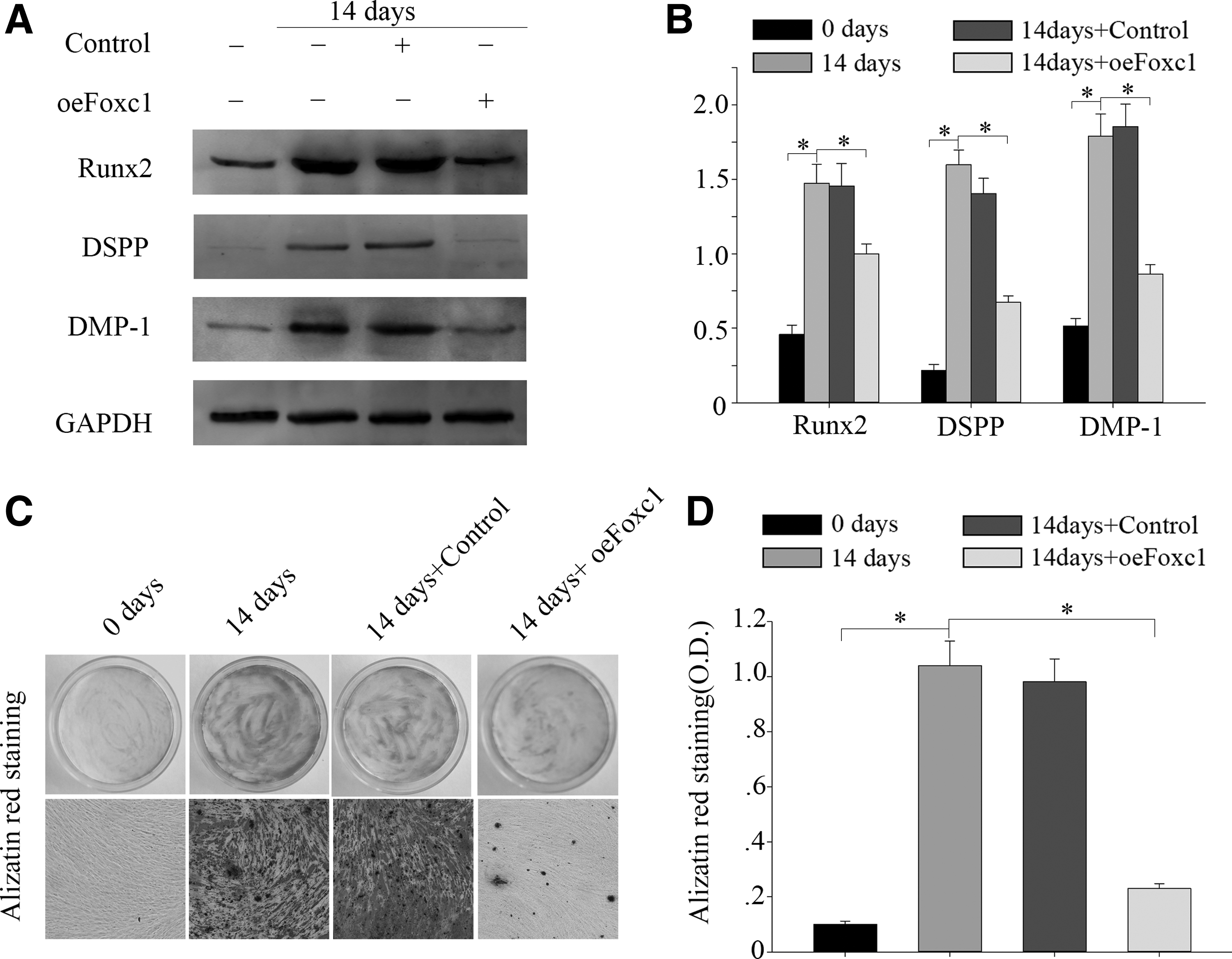
Overexpression of Foxc1 inhibits odontogenic differentiation in DPSCs. DPSCs were cultured in odontogenic differentiation medium for up to 14 days.
Discussion
The present research reported the role of Foxc1 in odontogenic differentiation of DPSCs for the first time. Since Foxc1 could control differentiation functions in osteoblast progenitor cells, we examined whether it would influence odontogenic differentiation in DPSCs. In our studies, we found that Foxc1 decreased in a time-dependent manner under odontogenic differentiation of DPSCs, and Foxc1 overexpression could inhibit the expression of odontogenic related markers Runx2, DSPP, and DMP-1. Furthermore, our results also showed that Foxc1 played a critical negative regulative role in odontogenic differentiation by controlling BMP/Smad signaling pathway.
Foxc1 is an important gene encoding transcription factors which has been proved to play a critical role in many embryonic organs and bone tissue differentiation (Bin et al., 2016; Motojima et al., 2017; Rice et al., 2005). Foxc1 is an important regulator of calvarial bone development and different types of bone differentiation (Hopkins et al., 2016; Rice et al., 2003, 2005). The mice which have knocked out the Foxc1 have extensive abnormalities, including obvious skeletal abnormal development, tissue homeostasis injury, and open eyelids (Rice et al., 2003, 2005; Sun et al., 2013; Wang et al., 2016). Foxc1 as a potential prognostic biomarker can regulate cellular functions that are also required for basal-like breast cancer development (Ray et al., 2010).
However, the reports on how Foxc1 mediates bone formation are limited. In the present study, it is the first time that we investigated the role of Foxc1 on the odontogenic differentiation of DPSCs. We found that Foxc1 expression was decreased gradually when DPSCs were stimulated to differentiation. Meanwhile, Foxc1 stable overexpression led to the expression and activity's decline of DSPP, DMP-1, and Runx2, as well as decreased the mineralization of DPSCs.
Odontogenic differentiation of DPSCs is mediated by various signaling pathways. Studies have shown the importance of BMP/Smad signaling pathway in regulating osteoblast differentiation of MSCs (Aquino-Martinez et al., 2017; Yang et al., 2013; Yoshida et al., 2000). A recent research shows that CCN3 can promote dentinogenesis and regulate odontoblastic differentiation of DPSCs by BMP/Smad signaling pathway (Wang et al., 2014).
Meanwhile, it is also reported that Foxc1 participates in the BMP signaling and negatively regulates Msx2 and Alx4 (Rice et al., 2003). Foxc1 is regulated by BMP4 and directly acts on the Msx2 in the progenitor cells to the osteoblast differentiation (Hopkins et al., 2016; Mirzayans et al., 2012). Foxc1 has been known to be an essential regulator of bone formation and differentiation; yet, how to control the Foxc1 functions of these processes is not completely clear. Thus, we testify whether increasing expression of Foxc1 can change the BMP/Smad signaling pathway. Our results indicate that Foxc1 regulates BMP/Smad signaling by inhibition of Smad1/5 phosphorylation.
Moreover, odontogenic differentiation markers, including DSPP, DMP-1, and Runx2, were suppressed by Foxc1 overexpression. These results suggested that one of the pathways that was inhibited by Foxc1 might be the BMP pathway in DPSCs. Some studies have shown that MSCs express a significantly high level of Foxc1, which can differentiate into osteoblasts and chondrocytes but lack the potential differentiation of adipocytes (Matsuoka et al., 2015). This shows that Foxc1 is closely related to chondrogenic differentiation in MSCs. However, we do not study the chondrogenic or adipogenic differentiation of DPSCs in this experiment. Our future research will consider experiments related to the chondrogenic or adipogenic differentiation of DPSCs.
In summary, the present study suggests the effect of Foxc1 overexpression on the odontogenic differentiation of human DPSCs. Our further studies will prove whether Foxc1 would also contribute to activation or mediation of other associated pathways. Meanwhile, the role of Foxc1 will be focused on developing deeper understanding in osteogenic differentiation. Hopefully, our research can provide some inspiration for improving dental regeneration and repair.
Footnotes
Acknowledgments
This work was partly supported by Top Six Types of Talents Financial Assistance of Jiangsu Province Grant (BRA2016527), the National Natural Science Foundation of China under Grant (Nos. 81671616, 81500809, 81471603), and Graduate Student Innovation of Science and Technology Projects in Nantong University Grant (No. YKC16112).
Author Disclosure Statement
The authors declare no conflicting financial interests exist.