
Abstract
Abstract
The human umbilical cord blood (HUCB) is an excellent source of adult stem cells, having the benefit of being younger than the bone marrow stem cells. The role of stem cells in the lesion repair mechanism is still being studied. We evaluated the capability of HUCB to interfere into the fibroblast dedifferentiation plasticity through cocultivation. Direct and indirect cocultures were maintained for 24, 48, and 72 hours. Coculture viability was evaluated by MTT assay. The messenger RNA was extracted, and the expression of p16 and p21 genes was estimated by quantitative reverse transcriptase polymerase chain reaction (qRT-PCR). The direct or indirect contact did not interfere with fibroblast cell viability. However, these direct and indirect contacts reduced the expression of p16 and p21 genes. A sigmoidal curve was applied to adjust gene expression against time, and a mathematical function was established for gene expression according to cell culture type. These results suggest that the differentiated cells were influenced by immature cells (HUCB) either by the direct contact or by signaling molecules, which alter their behavior and plasticity. Therefore our data may contribute to paracrine effects other than the commonly known to be responsible for the repair of lesions in stem cell therapy.
Introduction
C
The hematopoietic stem cells can reestablish all hematopoietic systems with a high rate of patient's survival. These cells are found in the bone marrow, peripheral blood, or umbilical cord. Stem cells can differentiate and self-renew into different types of tissues and specialized cells (Gluckman, 2012). The differentiated cells are responsible for the maintenance of organs and present beneficial effects through paracrine signals at the injury site. The use of human umbilical cord blood (HUCB) cells is easy to harvest without causing harm to the patient, and once the procedures become simpler, it can be widely applied. Adult stem cells can be maintained and expanded in the correct medium and growing conditions (Roura et al., 2015).
Recently, an experimental study has shown that mice fibroblasts may increase the modulation of the plasticity when cultured with the mononuclear cells of the umbilical cord blood. In this study, the fibroblast modulation of plasticity was confirmed by the expression, after induction, of pluripotent genes and by differentiation into cells with the mesodermic origin and into neural progenitors (Marinowic et al., 2015).
The paracrine effects of mesenchymal stem cell therapy have different therapeutic applications. Secreted signaling molecules promote immunomodulatory, antiapoptotic, pro-inflammatory, self-renewal, and differentiation effects. This occurs according to the regeneration needs of the injured tissues, by the presence of local mediators (Meirelles Lda et al., 2009). Changes in the plasticity levels stimulated by contact with stem cells may set a new mechanism of an indirect action of tissue recovery mediated by the use of cell therapy in several models (Vishnubalaji et al., 2012). This cell dedifferentiation can be mediated by a direct contact or through the action of signaling molecules secreted by the stem cells at the lesion niche.
The ideal coculture for dedifferentiation, whether contact-dependent or paracrine-dependent mediation, is still unknown. Therefore, since fibroblasts have been considered an ideal model to study the cellular and molecular features such as proliferation, cell–cell interactions, and signaling, this study will evaluate the capability of cells from HUCB to interfere into the skin fibroblast dedifferentiation plasticity through cocultivation.
Materials and Methods
Fibroblast samples
The present study was reviewed and approved by the Research Ethics Committee of the Pontifical Catholic University of Rio Grande do Sul (approval no. 1.449.268) through the system Plataforma Brasil. Fibroblasts were obtained from the residual skin fragments from one healthy, infectious disease-free patient (aged 41 years, female) who underwent surgical treatment for plastic surgery in the Plastic Surgery Program at São Lucas da PUCRS Hospital, upon signing the ethical consent form, under the guidance of the Committee of Research Ethics.
The fibroblasts were cultured in Dulbecco's Modified Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific), 100 U/mL penicillin (Gibco; Thermo Fisher Scientific), 100 U/mL streptomycin (Gibco; Thermo Fisher Scientific), and 100 μg/mL gentamicin until the five passages. For the cocultures, the fibroblasts were grown in a 24-well plate at the concentration of 2.6 × 104 cells per well at 37°C with 5% CO2 during 24 hours.
Sample of umbilical cord blood mononuclear cells
The samples (three) were collected by puncturing the arteries and veins of the umbilical cord and placenta with a 20-mL syringe containing 1 mL of heparin 5000 IU/mL. For the separation of the mononuclear fraction, the collected materials were diluted in RPMI 1640 Medium (1:1; Gibco; Thermo Fisher Scientific). This suspension was fractionated in a density gradient generated by centrifugation over Histopaque® 1.077 g/L (Sigma-Aldrich, St. Louis, MO) at 400 × g for 30 minutes at 25°C. The mononuclear fraction over the Histopaque interface was collected and washed twice with Dulbecco's Phosphate-Buffered Saline (DPBS) buffer.
The cell viability was determined using the exclusion method by trypan blue assay. The mononuclear fraction of each umbilical cord was cocultured with the fibroblast in the same well (direct coculture) and using an insert with 0.4 μm of porosity (indirect coculture). The cell density was 104 cell per well. The cultured fibroblasts were maintained at 37°C and 5% CO2 until the cells were collected after 24, 48, and 72 hours. The mononuclear cells were sharing, directly and indirectly, the medium with fibroblasts. The fibroblasts were removed with the addition of 1 mL trypsin at a concentration of 0.25%, so the cells could be released from the bottom of the plate. The content was collected and centrifuged at 400 × g for 5 minutes. The cell precipitates were used for RNA extraction.
MTT analysis
For each time period analyzed, 96-well plates with 200 μL of DMEM-supplemented medium were prepared, each well contained 4.5 × 104 cells, which were cultivated for 24 hours and then washed with 1 × DPBS. The wells containing only the cell cultures were used for the negative toxicity-free control group. After 24 hours, the culture medium was removed, and 200 μL of 5 mg/mL MTT solution in the DMEM-supplemented medium was added. The cells were then incubated for 4 hours at 37°C and 5% CO2. MTT was then solubilized in 200 μL of dimethyl sulfoxide (Sigma-Aldrich) for the analysis. The optical densities were measured at 570 nm in an ELISA reader (Bio-Rad, Hercules, CA). The cell viability was calculated using the following formula:
Molecular analysis through qRT-PCR
The molecular analysis of genes related to cell cycle was performed in both insert and coculture cells of 24, 48, and 72 hour groups. RNA was extracted using an SV Total RNA Kit (Promega, Fitchburg, WI). The samples were exposed to lysis buffer, heated at 70°C for 3 minutes, and centrifuged at 12,000 × g for 10 minutes at 4°C. The supernatant was collected, and 200 μL of 95% alcohol was added to it. The contents were transferred to a silica column and centrifuged at 12,000 × g for 1 minute. In addition, 600 μL of buffer was added to the solution and centrifuged again at 12,000 × g for 1 minute. Diluted DNAse enzyme was added to the contents and incubated for 15 minutes at room temperature. The columns were then washed with wash buffer, and the silica-retained RNA was eluted with the addition of 100 μL of RNAse-free water.
The extracted RNA was quantified using NanoDrop Lite (Thermo Fisher Scientific) from 1 μL of each extracted RNA sample. For gene expression analysis, a quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) technique was performed using StepOne Plus (Life Technologies; Thermo Fisher Scientific) equipment. RNA amplifications (10 ng) were performed for each sample. The trials were performed on individual plaques for each gene using the GAPDH gene as an endogenous expression control. For cell cycle and senescence analysis, the complementary primers to the messenger RNA (mRNA) sequence of p16 and p21 genes were used. The gene characteristics are described in Table 1.
CDK, cyclin-dependent kinase.
The amplification of RNA was performed using SYBR® Green One-Step qRT-PCR and HotStart-IT Kit (Affymetrix, Cleveland, OH) according to the manufacturer's instructions. In addition, 45 thermal cycles at 95°C for 15 seconds, 60°C for 20 seconds, and 72°C for 45 seconds were performed. The primers used are described in Table 2.
F, forward; R, reverse.
Analytical theory of gene expression competition
To explain the basic structures of the networks considered in our analysis, the sigma cycle in the two gene competitors was used. The sigmoidal curve was fitted to our data using CurveExpert Professional 2.6.4 software, and their validity was manually verified in Microsoft Excel.
Statistical analysis
For the data analysis related to gene expression, the 2−ΔΔCt relative quantification method was used for subsequent analyses using the endogenous control GAPDH gene and one-way analysis of variance test (SPSS Software, Inc.) with the Bonferroni posttest. The results were considered statistically significant at p < 0.05 for the samples that were confirmed for the genes in the cell cycle.
Results
The direct or indirect contact between the fibroblast culture and the mononuclear cells of the HUCB did not promote cell viability as observed in the MTT assay. During all analyzed time points, the cell viability of fibroblasts remained 100% regarding the control of the reaction (Fig. 1).
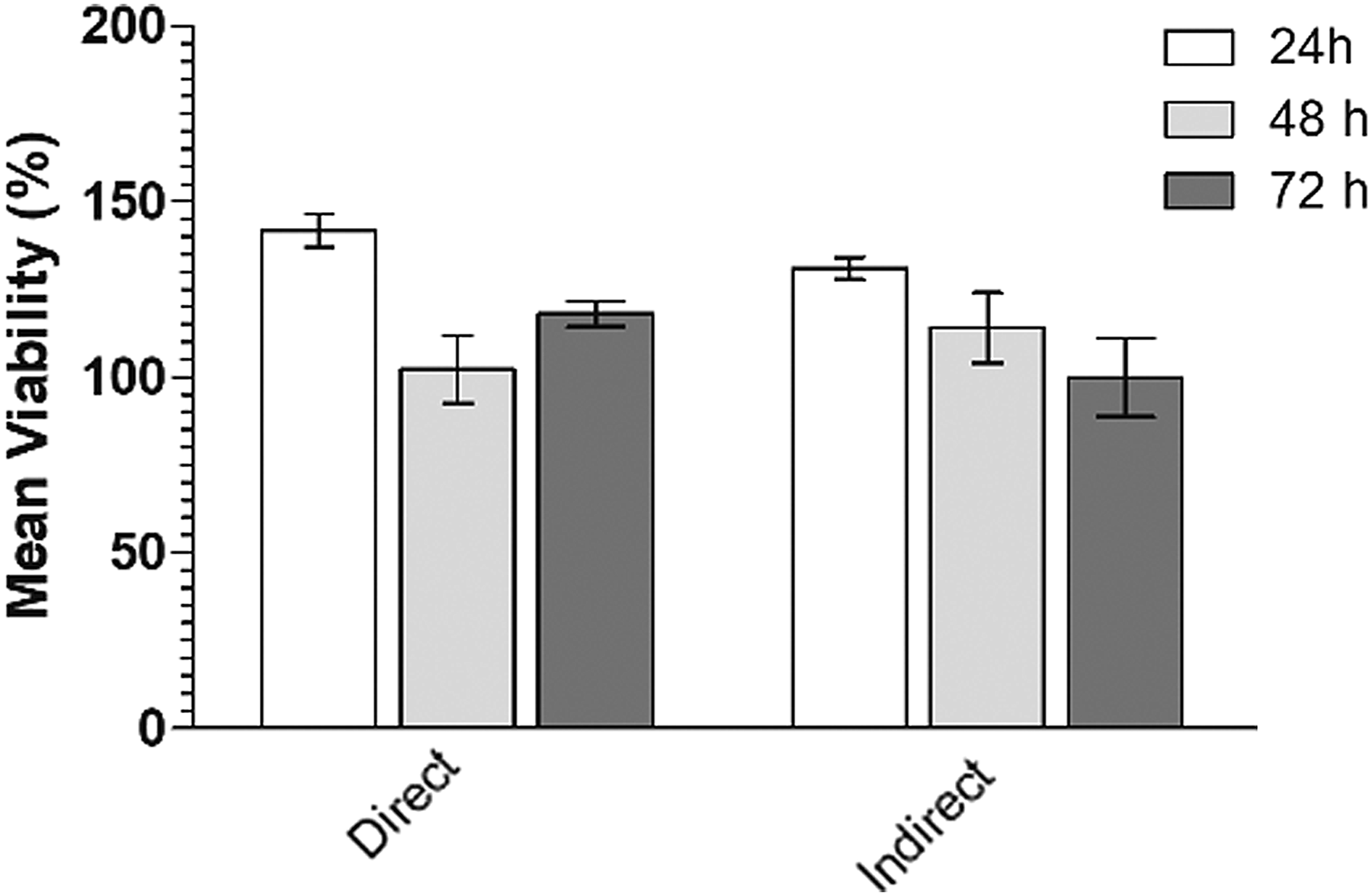
The cell viability measured by MTT assay. The data are presented as mean values and express the viability as a percentage of the control value.
The directly cocultured fibroblasts showed a reduction in p16 gene expression after 24 hours of exposure (1.48 times). In the indirect coculture, the RE of p16 at 24 hours was 2.23 times less compared with the control. At 48 hours, an analysis showed a reduction of 14.43 times in p16 expression and, after 72 hours, a reduction of 5.81 times, presenting the statistically significant difference between the time points (p = 0.0005). A comparison of the relative p16 expression between direct and indirect cocultures showed a statistically significant difference at 24 and 48 hours (p = 0.0005 and p = 0.005, respectively). A decreased p16 expression was observed at 72 hours in the direct cocultures compared with the insert but without a statistically significant difference (Fig. 2).

p16 mRNA expression in fibroblast at 24, 48, and 72 hours of direct and indirect coculture with umbilical cord mononuclear cells, ***p < 0.001; +++p < 0.001. mRNA, messenger RNA.
During direct coculture, the RE of p21 gene was decreased by 13.88 times after 24 hours of exposure compared with the control. At 48 hours, fibroblasts presented a decreased RE (2.91 times; p = 0.0005), and after 72 hours, an increase in p21 expression was observed (1.67 times; p = 0.0005). The cells cocultured through the insert showed a decrease of 20.65 times in the relative p21 expression after 24 hours, and at 48 and 72 hours, the RE decreased by 1.68 and 2.35 times, respectively (p = 0.0005).
No difference was observed in p21 expression between the direct and indirect cocultures after 24 hours. After 48 and 72 hours, lower (p = 0.0005) and higher (1.97 times) p21 expression in the direct coculture than the control was observed. In addition, the direct coculture was less expressed by 2.35 times in the insert group, and this difference was statistically significant (p = 0.0005) (Fig. 3).
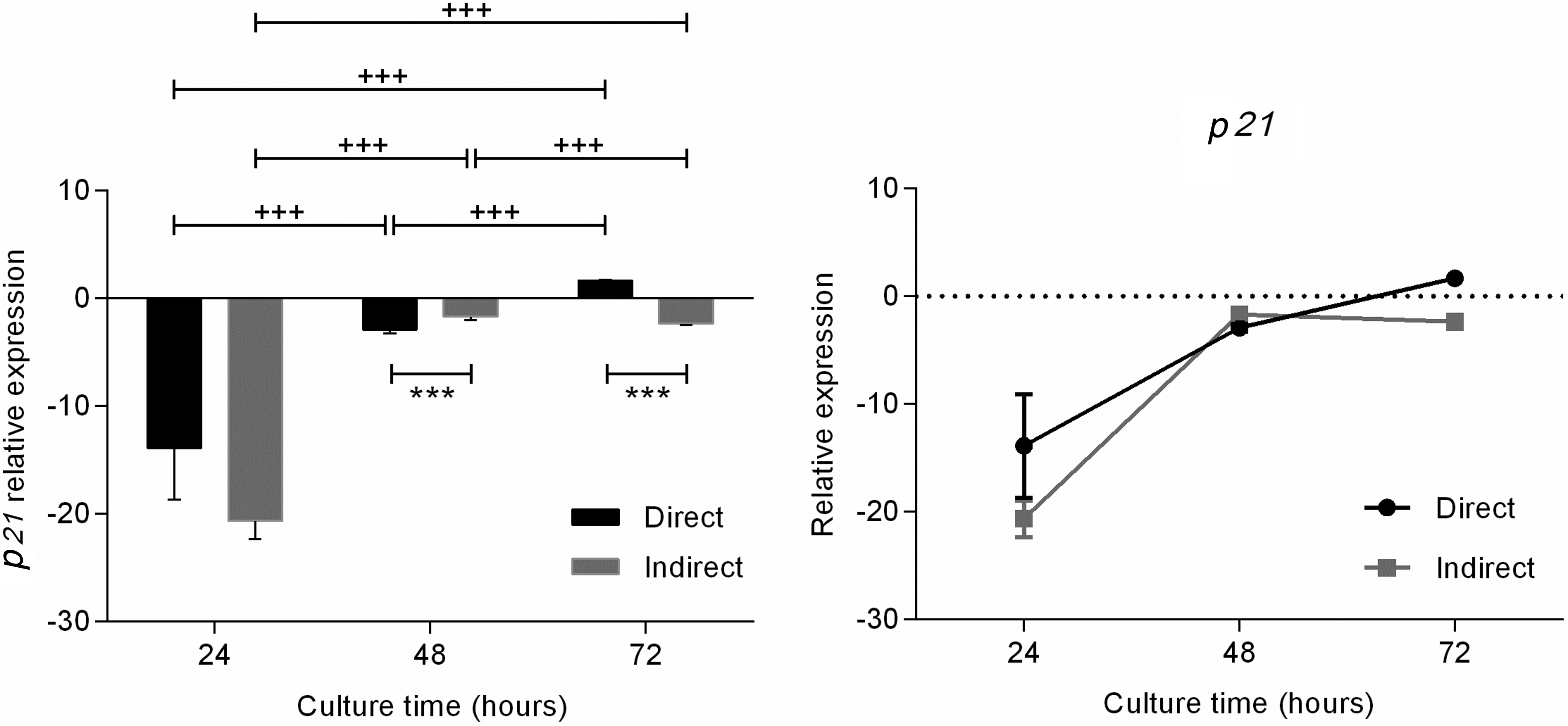
p21 mRNA expression in fibroblast at 24, 48, and 72 hours in direct and indirect coculture with umbilical cord mononuclear cells, ***p < 0.001; +++p < 0.001.
At 24 hours, p21 direct coculture was expressed lower by 12.3 times than p16 (p = 0.0005), and in the indirect culture, p21 was expressed lower by 18.3 than p16 (p = 0.00005). At 48 hours, the expression of p21 and p16 did not present a statistically significant difference in the direct culture. However, in cells cultured through the insert, p16 expression was 12.7 times lower than p21 expression (p = 0.0005). After 72 hours, p16 expression was 9.5 times lower than the p21 expression in the direct culture and 3.5 times lower in the indirect culture (Fig. 4).
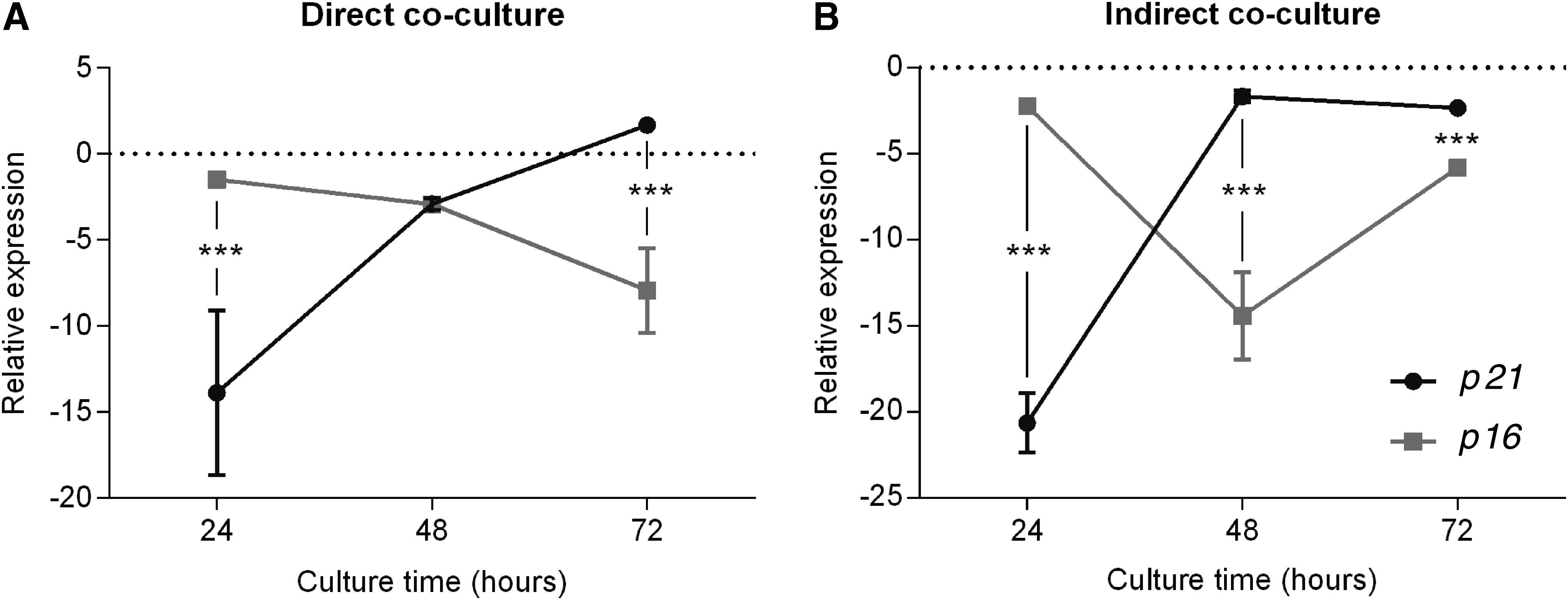
Relationship between p16 and p21 mRNA expression in fibroblast at 24, 48, and 72 hours in direct coculture
The expression of p16 and p21 genes in each culture method was plotted against time, and sigmoidal curves were fitted to obtain the RE functions:
Discussion
Cell reprogramming can be achieved in different manners, including nuclear transfer, a fusion of somatic cells with embryonic cells, and reprogramming by gene transfection (Cowan et al., 2005; Giordano et al., 2007; Nakagawa et al., 2008; Tada et al., 2001). Several characteristics are involved in classifying an in vitro cell culture as senescent or young because of its morphology, beta-galactosidase activity, and an irreversible decrease in the regularity of cell cycle. In addition, the chromosomes of a senescent cell form distinct domains of heterochromatin called senescence-associated heterochromatin foci (Narita, 2007).
The cellular senescence consists of the loss of proliferative potential produced by the accumulation of cell doublings. The study of cellular senescence in cell culture has become an experimental system of great value for understanding the changes of aging processes (Bringold and Serrano, 2000).
Marinowic et al. (2015) showed that the direct coculture of 3T3 fibroblast lines with the HUCB mononuclear cells altered the fibroblast plasticity evidenced by KLF4 gene expression and a potential for inducing neuronal differentiation. Fibroblasts were cultured for 6 days with HUCB mononuclear cells and reprogrammed. Subsequently, the fibroblasts were differentiated into the neuronal lineage, presenting positive staining for different neuronal markers.
The p16 and p21 tumor suppressor proteins are the key regulators of the cellular senescence process that are activated by increasing cell age (Beausejour et al., 2003). The p21 protein is a negative regulator of the cell cycle and may interfere with the proliferation rate by controlling cell cycle progression. It is a cyclin-dependent kinase (CDK) inhibitory protein that controls the transition between the cell cycle phases being specific and selective by acting on the cyclin D-CDK4 and D-CDK6 complexes that act in the G1 phase (Rizzo et al., 2011). During the first 24 hours of exposure to the HUCB mononuclear cells, cocultured fibroblasts, both directly and indirectly, showed a considerable decrease in p21 expression.
The analyses at 48 and 72 hours presented expression values similar to the fibroblasts that were not cocultured. The considerable decrease in p21 expression after 24 hours suggests an increase in the cell cycle mediated by the contact of the fibroblasts with the HUCB mononuclear cells. The primary cultures of human fibroblasts have a limited proliferative life, and as soon as they reach cell senescence, their growth is irreversibly ended after a determinate number of divisions (Bond et al., 1999). The decrease in p21 expression after 24 hours was mediated by the direct contact between cells and by sharing the culture medium. This finding suggests that soluble molecules potentially secreted by HUCB mononuclear cells influence the fibroblast gene expression when associated with their proliferative capacity.
The HUCB mononuclear cells play a role in altering the expression of proteins responsible for the cell cycle, thus influencing their proliferative capacity and consequently their plasticity, preparing the differentiated cells to contribute to a possible role in tissue repair. Our results suggest that the first 24 hours of contact with the HUCB mononuclear cells decreases p21 expression. This decrease can stimulate cells to increase the rate of cell division, a feature associated with the youth of cells in culture. After 48 and 72 hours of contact, few changes were observed in p21 expression, where the negative regulation of this gene expression did not influence the rates of cell division.
The considerable decrease in the p21 expression regarding control was normalized during the analyzed periods for both direct and indirect cocultures, reaching values similar to the control in the analysis at 72 hours.
The p16 protein is a crucial tumor suppressor, commonly inactivated in most human tumors, and a negative regulator of the cell cycle (Ruas and Peters, 1998). The p16 expression did not show considerable changes after 24 hours of contact between fibroblasts and HUCB mononuclear cells. After 48 hours, the insert-mediated coculture displayed the lowest p16 expression, which was the highest reduction in the gene expression during the three analyzed time points and on both types of cultures. After 72 hours, a decrease in p16 expression was observed in both direct and indirect cocultures compared with the control.
During the direct coculture, a gradual reduction in p16 expression was observed, thus achieving the lowest RE value after 72 hours of cell-to-cell contact. The factors secreted by HUCB mononuclear cells reduce p16 expression gradually and, thus, increase their rate of cell proliferation by themselves after 72 hours. The change in the behavior related to the fibroblast plasticity mediated by the contact with the mononuclear fraction was interceded by the reduced p16 expression, particularly at 72 hours. This is different with regard to p21 expression, where the lowest expression values occurred during the first 24 hours.
As there was a direct contact, alterations in the gene expression related to cell cycle control occurred throughout the period of the cell exposure, during the initial phase mediated by a decrease in p21 expression and the final phase mediated by a decrease in p16 expression. The insert-mediated culture showed lower RE after 48 hours, revealing that after this period, the HUCB mononuclear cells did not influence p16 expression in the fibroblasts.
Huang et al. revealed that the supernatant of the umbilical cord mesenchymal stem cell culture and the direct coculture of these cells with trophoblasts stimulated a remarkable increase in the migration and invasion of trophoblast cells in addition to an increase in the cell proliferation capacity.
The results indicate that during all analyzed time points for either direct or insert-mediated culture, there was a decrease in the gene expression associated with the negative control of the cell cycle. However, this appears to be an antagonism in the expression profile between p21 and p16. At 24 hours, the large decrease in p21 expression of the two groups appears to inhibit the decrease in p16 expression, and in subsequent times, the decrease in p16 expression appears to influence the decrease in p21 expression in both direct coculture and insert mediated.
Several paracrine effects have been attributed to tissue repair mechanisms mediated by stem cell therapy. The stem cells act at the injury site by releasing several paracrine factors that have potential to improve the general condition caused by a certain damage or tissue recovery in the lesions of various orders (Zanirati et al., 2015). These paracrine effects include immunomodulatory, antiapoptotic, proangiogenic, chemoattractant, antifibrosis, and support for growth and differentiation of endogenous progenitor cells (Karlsson et al., 2008; Meirelles Lda and Nardi, 2009).
Recent studies have redirected attention to one of the mechanisms of action of the stem cells, which is regulating the innate and adaptive immune system and triggering several crucial effector functions in normal tissue and in pathological situations (Ben-Ami et al., 2014; Singer and Caplan, 2011; Uccelli et al., 2008). In addition, age-related deficits may be associated with an impaired immune response mediated by the stem cells of various organs.
The robust reduction of stem cells during aging may occur because of cell senescence and can be caused by several intrinsic stimuli, oncogenes, and natural and pathological changes in the stem cell microenvironment (Rao and Mattson, 2001; Singer and Caplan, 2011; Wang et al., 2011). In addition, umbilical cord stem cells can secrete several crucial growth factors such as epidermal growth factor, fibroblast growth factor, glial cell derived neurotrophic factor, brain derived neurotrophic factor, and platelet derived growth factor among others, which are crucial modulators of the various tissue genesis (Guo et al., 2015).
p16 and p21 genes are two independent CDK inhibitors and are similarly involved in various processes, including the regulation of the critical G1 to S phase transition of the cell cycle, senescence, and apoptosis. The expression of p16 and p21 is under the control of different activators and suppressors that regulate both proteins with different mechanisms (Ruas and Peters, 1998). The similarities between these two genes suggest the presence of a direct or indirect interaction or regulation between these two key cell proliferation regulators.
Al-Khalaf and Aboussekhra (2013) using cell lines with the different levels of expression of p16 and embryonic cells, p16-proficient (p16+/+) and p16-deficient (p16−/−), showed that p16 stabilizes the CDKN1A mRNA, revealing that a link exists between p16 and p21 expression, because the expression of p21 gene is modulated in a p16-dependent manner.
In this study, we conclude that both the direct and indirect contacts between fibroblasts and HUCB mononuclear cells can decrease the expression of p16 and p21, which are associated with the negative control of the cell cycle. However, in the direct or indirect cocultivation, both p16 and p21 are negatively expressed in fibroblasts in a competitive sigmoid form. The cocultivation appears to decrease the expression of the two genes with the difference in the level of decrease in the first evaluation time, suggesting impairment in the decrease of p16 expression when p21 levels are greatly reduced. In a due course of time, this expression profile undergoes an inversion, with p16 having a low level of expression compared with p21.
This result suggests that differentiated cells can be influenced by immature cells, altering their behavior and plasticity and, thus, may contribute to other paracrine effects for the lesion repair using stem cell therapy. The residual cells of the penumbra zone mediated by the stem cell action at the lesion site can be stimulated to contribute to a greater plasticity and support the recovery of the damaged tissue.
The indirect effects attributed to stem cells in the promotion of tissue recovery play an extremely crucial role, resulting in tissue, cognitive, and motor recovery in experimental models, without effectively presenting transdifferentiation mechanisms at the lesion site. The conditioning and enhancement of residual cell plasticity after injury may contribute to the paracrine factors for tissue regeneration. We suggest in this study a possible stem cell action in the promotion of the dedifferentiation of compromised cells in relation to their plasticity; this is an innovative mechanism attributed to cellular therapy.
Footnotes
Acknowledgments
The authors thank the student Kiyo Costa Higuchi for mathematical contributions. DaCosta is CNPq researcher. Marinowic, Zanirati, Azevedo, De Souza, and Bruzzo were awarded with a fellowship from Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES) or CNPq (Brazil). This work was supported by research grants from the Conselho Nacional de Desenvolvimento Cientifico e Tecnologia (CNPq) and Pandurata Alimentos LTDA.
Author Disclosure Statement
The authors declare that there are no financial conflicts of interest.