
Abstract
Abstract
Spermatogonial stem cells (SSCs) self-renew and produce a large number of differentiated germ cells to maintain normal spermatogenesis. However, the growth factors crucial for SSC self-renewal and the mechanism underlying this process remain unclear. In the present study, a serum-free culture media was used to evaluate the effect of several growth factors on the expression of some SSC markers and self-renewal related genes. The putative SSCs were cultured on buffalo Sertoli cell feeder layer in KO-DMEM +10% KOSR. The colony formation was observed between 7 and 10 days. The putative SSC colonies also expressed markers specific for undifferentiated type A spermatogonia and pluripotency markers. After 15 days, relative mRNA expression study revealed that 20 ng/mL concentration of Glial cell line-derived neurotrophic factor (GDNF) upregulated the expression of PLZF, TAF4B, and THY1. Furthermore, supplementation of a combination of 20 ng/mL GDNF, 10 ng/mL basic fibroblast growth factor (bFGF), 1000 IU/mL leukemia inhibitory factor (LIF), and 1 ng/mL colony stimulating factor 1 (CSF1) upregulated the expression of PLZF, TAF4B, BCL6B, and ID4 genes. These results demonstrated that our defined culture media in combination with GDNF, bFGF, LIF, and CSF1 well supported SSC self-renewal.
Introduction
Among the adult stem cells of a male organism, spermatogonial stem cells (SSCs) are the most fascinating for their ability to generate an infinite number of spermatozoa throughout the life, as well as for their role in transfer of the genome to future generations. These properties render SSCs a special place in transgenerational research for the important role they could play in the production of organisms of choice.
Pertinently, the most important aspect of the research focuses on development of techniques for efficient isolation of these cells from the testicular tissues and their subsequent expansion under in vitro cultures so as to generate sufficient numbers for their modification (genetic or chemical), tailored differentiation, and reintroduction into their niche for carrying the introduced modifications across the generations. Given their small number under natural conditions within the testicular tissue (1 in 3000–4000 cells in adult mouse testes), their efficient isolation, purification, and expansion demand rigorous and vast studies across a number of species for establishment of the most efficient and reproducible protocols.
A good number of studies have demonstrated that the testicular niche where the SSCs reside possesses a large number of growth factors, which could presumably be responsible for determining the fate of the stem cells (Brinster, 2002; Kubota et al., 2004a, 2004b). Among the various growth factors whose presence has been established within the testicular tissue, Glial cell line-derived neurotrophic factor (GDNF), basic fibroblast growth factor (bFGF), leukemia inhibitory factor (LIF), and epidermal growth factor (EGF) are considered to play essential roles for maintenance and propagation of SSCs in mouse and cattle testes (Aponte et al., 2008; Kanatsu-Shinohara et al., 2003; Kubota et al., 2004a, 2004b).
Pertinent to their exploitation in enhancing the fertility and usage as vehicles for carrying the induced genes to future generations, studies were initiated for establishment and expansion of buffalo (Bubalus bubalis) SSCs in our laboratory, given the significance of bubaline species as milch, meat, and draught animal within the subcontinent. The studies have assumed more importance considering the fact that the species is regarded as a poor breeder due to its low reproductive efficiencies in terms of delayed onset of puberty, long calving intervals, poor conception rate, anestrus, and seasonality (Mahla et al., 2012).
The present study was thus undertaken to develop serum-free culture system for buffalo SSCs and their subsequent expansion in undifferentiated state. The self-renewal under defined culture media was assayed through related gene expression data, so as to establish an optimized medium for the purpose. The need for development of serum-free culture system was felt from previous studies in mouse and other species where it was reported that the use of serum introduces certain complex and undefined parameters thereby rendering it difficult to determine the regulatory mechanisms for SSC self-renewal and differentiation (Kubota et al., 2004a). It has also been reported that serum supplementation adversely affects proliferation and colony formation in mouse SSCs (Kubota et al., 2004b).
Although the culture and maintenance of mouse SSCs in serum-free medium are well established, scanty reports are available in other species. In bubaline species, Yu et al. (2014) demonstrated the successful in vitro culture of undifferentiated SSCs for 7 days by supplementing HyClone AdvanceSTEM cell culture media, which was completely devoid of serum. By utilizing Knockout Serum Replacer (KOSR), Aoshima et al. (2013) observed a continuous growth of mouse SSCs in vitro. In the present study, we examined the possibility of establishing putative buffalo SSC cultures in KOSR based medium supplemented with GDNF, bFGF, LIF, and CSF1. Based on gene expression data, the optimized concentration of growth factors was also elucidated to develop an optimized medium for buffalo SSC culture and propagation.
Materials and Methods
Unless mentioned otherwise, chemicals were purchased from Sigma Chemical (St Louis, MO). Plastic wares were procured from Nunc (Roskilde, Denmark) and nylon mesh filters from Millipore (Bedford, MA). Dolichos biflorus agglutinin (DBA) was purchased from Vector Laboratories (Burlingame, CA). Primary antibodies against OCT-4, SOX-2, and NANOG were purchased from Sigma Chemical and those against THY1, UCHL1, and PLZF were from Santa Cruz Biotechnology, Inc. (Dallas, TX). The guidelines of institutional ethics committee were followed for obtaining and handling of tissue samples.
Isolation and enrichment of putative SSCs
Prepubertal buffalo (3–6 months of age) testes were collected from a local abattoir. Isolation and enrichment were performed as described previously with some minor modifications (Izadyar et al., 2002; van Pelt et al., 1996). Briefly, minced seminiferous tissue was subjected to two-step enzymatic isolation followed by filtration through 80- and then 60-μm nylon mesh filters to enrich the SSC population. To separate other contaminating cells, the filtered cells were seeded onto DSA lectin–bovine-serum-albumin-coated (BSA-coated) dishes at a density of 2 × 105 cells/cm2 and were incubated overnight at 37°C in a CO2 incubator (5% CO2 in air).
Thereafter, the cells were subjected to discontinuous Percoll density gradient centrifugation (Kadam et al., 2013). The enriched SSCs found in the interface between 28%, 30%, and 32% Percoll suspensions were collected and used for the subsequent in vitro culture. The viability of cells was determined by trypan blue dye (0.4%) exclusion method using hemocytometer.
In vitro culture of putative SSCs
The enriched SSCs were seeded onto homologous Sertoli cell feeder layer and cultured in SSC medium [Knockout-DMEM (Invitrogen Corporation, Carlsbad, CA) supplemented with 10% Knockout serum replacer (Invitrogen)] containing 40 ng/mL GDNF at 37°C. The Sertoli cell feeder layer was prepared according to the protocol previously established in our laboratory (Kadam et al., 2013). The SSC colony formation was observed between 7 and 10 days of culture.
On day 15, the putative SSC colonies were split mechanically with a micro-blade and subcultured as treatment and control groups. The colony area was measured using an inverted microscope (Eclipse Ti; Nikon, Tokyo, Japan) with NIS-Elements BR 3.1 software (Nikon). The experimental groups were supplemented with growth factors as per the design of experiments. The medium was replaced every third day with fresh SSC medium supplemented with or without growth factors.
Characterization of putative SSCs using alkaline phosphatase activity and immunofluorescence staining
Alkaline phosphatase staining was performed using commercial kit (catalog no. 86C; Sigma Chemical) according to the manufacturer's instructions.
For immunofluorescence staining, on 12th day of culture, SSC colonies were fixed in 4% paraformaldehyde in DPBS for 30 minutes and permeabilized with 0.1% Triton X-100 in DPBS for 30 minutes. After blocking nonspecific sites with 5% BSA, the cells were incubated with Fluorescein-labelled DBA (1:100, FL-1031) for 1 hour at room temperature. For detection of PLZF (sc-22839), THY1 (sc-6071), UCHL1 (sc-25800), OCT-4 (O8389-100 UG), SOX-2 (SAB2103958-5), and NANOG (AV100591-50UG) expression, the SSC colonies were incubated overnight (16 hours) at 4°C with respective primary antibodies (1:50) and then with either FITC or PE labelled secondary antibody (1:500).
The nuclei were stained with Hoechst 33342. In negative control the use of primary antibody was omitted else the same staining procedure was used. The colonies were examined under a fluorescence microscope (Nikon).
Karyotype analysis of putative SSCs
The chromosomal integrity of putative SSCs was analyzed by karyotyping as described previously (Kala et al., 2012), with minor modifications. Briefly, the putative SSC colonies were incubated with 0.1 μg/mL colcemid for 4 hours at 37°C. The colonies were washed, trypsinized, and resuspended in 0.56% KCl and incubated for 30 minutes at 37°C. Subsequently, the SSCs were washed and then fixed in freshly prepared chilled fixative (3:1, methanol: glacial acetic acid) for 20 minutes followed twice by the same treatment each for 10 minutes. The metaphase spreads were prepared by dropping the cells from a height of 25–30 inches onto ice-cold glass slides and stained with 2% giemsa for 10 minutes. The glass slides were rinsed and observed under oil immersion (1000 × ) using an inverted microscope.
Quantitative real-time PCR analysis
Total RNA was extracted using RNAqueous-Micro Kit (Ambion, Austin, TX). The purity and concentration of RNA were determined by NanoQuant (Tecan, Salzburg, Austria). Superscript III First Strand cDNA Synthesis Kit (Invitrogen) was used for cDNA synthesis. Quantitative real-time PCR was performed on a CFX 96 I Cycler (Bio-Rad, Hercules, CA) using 10 μL reaction volume containing 5 μL of SYBR Green Master Mix (Maxima SYBR Green Mastermix; Fermentas; Fisher Scientific, Pittsburgh, PA), 0.2 μL of 10 μM of each primer, and 20 ng of cDNA. The thermal cycling conditions consisted of initial denaturation at 95°C for 10 minutes, followed by 40 cycles of 30 seconds at 95°C, 30 seconds at the appropriate annealing temperature (Table 1), and 30 seconds at 72°C, followed by 95°C for 10 seconds and melting curve.
Primers Used for Gene Expression Studies
Relative gene expression levels were determined using the 2−ΔΔCt method, where ΔCt = Ct target–Ct internal reference and ΔΔCt = ΔCt target – ΔCt calibrator. In all experiments, GAPDH served as the internal reference gene. Gene expression data were normalized against GAPDH expression and were analyzed using CFX Manager (Bio-Rad). The calibrator in each study consisted of cDNA from the corresponding control group. Relative mRNA expression is expressed as n-fold mRNA expression relative to the calibrator. The specificity and integrity of the PCR products were ensured by melting curve analysis (Supplementary Figs. S1 and S2; Supplementary Data are available online at www.liebertpub.com/cell), whereas the appropriateness of size was confirmed by agarose gel electrophoresis as shown in Supplementary Figure S3. In all experiments, three trials were performed, each with three replicates.
Statistical analysis
Data are expressed as mean ± SEM. Differences in the means were tested using one-way ANOVA. GraphPad prism (version 7) software was used for statistical analysis.
Results
Isolation, enrichment, in vitro culture, and characterization of putative SSCs
The average cell yield after two-step enzymatic isolation was ∼1 × 106 cells/g of testis, and the viability of the isolated cells was 80–85%. The SSC colony formation was observed between day 7 and 10 of culture, and the area of these colonies increased consistently up to day 15 as shown in Figure 1. The colony area between day 7 and 10 (30,100 ± 5054 μm2) and day 15 (85,410 ± 4116 μm2) differed significantly (p < 0.05). We observed a high alkaline phosphatase activity (Fig. 1) and a normal karyotype (Supplementary Fig. S4) in these cultured putative SSCs. Immunofluorescence analysis revealed that these colonies expressed DBA, PLZF, UCHL1, and THY1 (Fig. 2), which are specific markers for undifferentiated type A spermatogonia. These putative SSCs also expressed pluripotency related markers like OCT-4, SOX-2, and NANOG as shown in Figure 3.
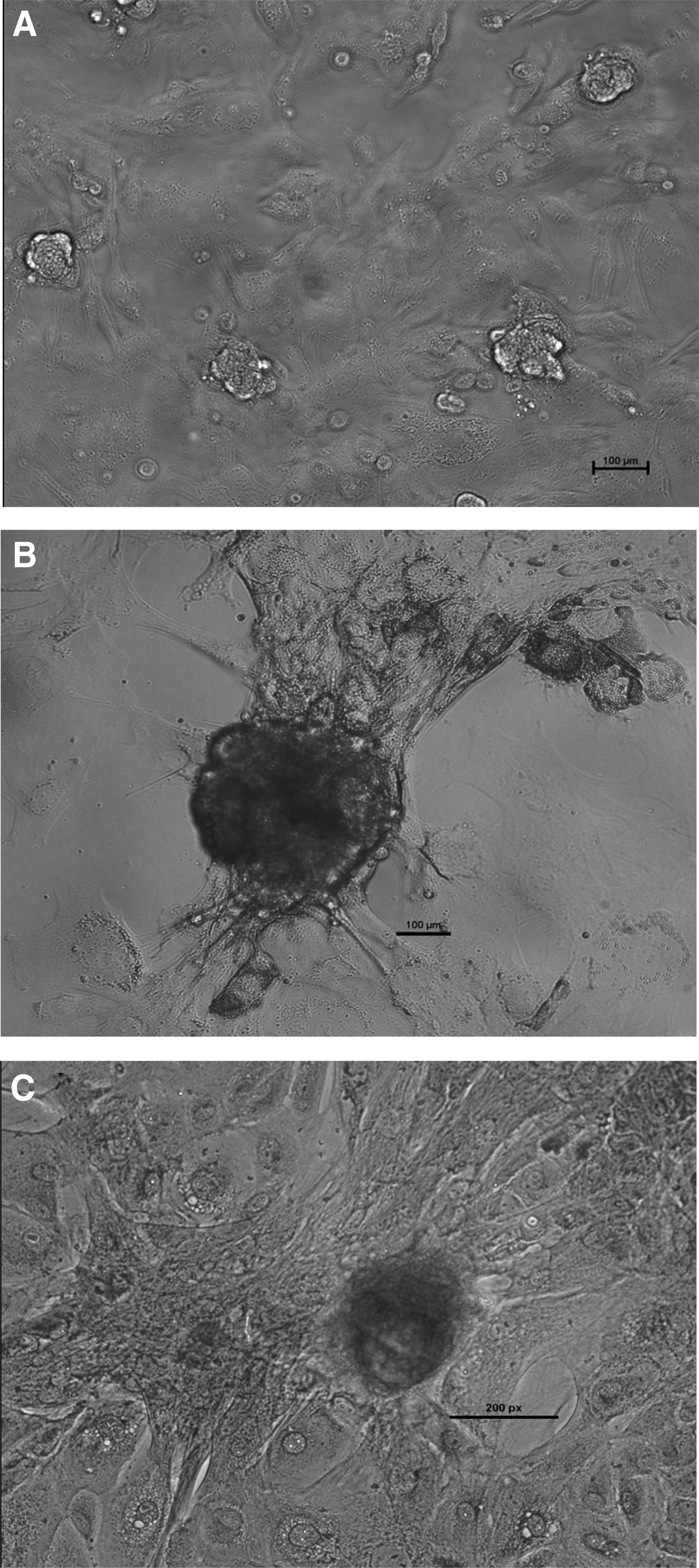

Immunocytochemical analysis of markers specific for undifferentiated type A spermatogonia DBA, PLZF (200 × ; 200 μm); THY1 (100 × ; 200 μm), UCHL1 (100 × ; 100 μm); Control (200 × ; 100 μm) in putative SSC colonies. Hoechst 33342 nuclear stained image (column A), secondary antibody labeled image (column B), merged image of column A and B (column C). DBA, Dolichos biflorus agglutinin.
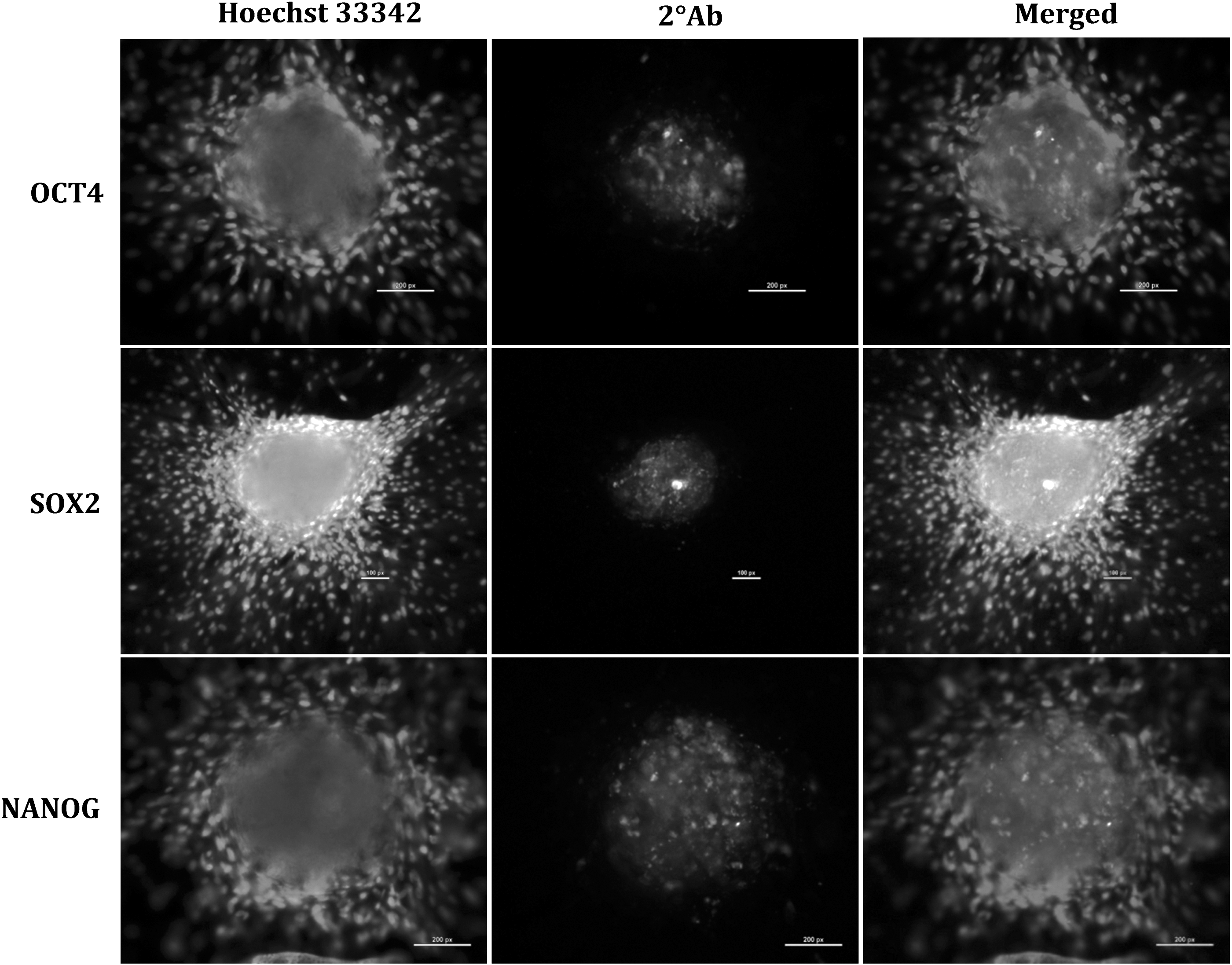
Immunocytochemical analysis of pluripotency markers OCT4 (200 × ; 200 μm), SOX2 (100 × ; 100 μm), and NANOG (200 × ; 200 μm) in putative SSC colonies. Hoechst 33342 nuclear stained image (column A), secondary antibody labeled image (column B), and merged image of column A and B (column C).
Effect of different concentrations of GDNF on quantitative expression of some SSC markers and self-renewal related genes
This experiment was aimed at examining the effect of different GDNF doses on the expression of some SSC markers and self-renewal related genes. Before being seeded on the Sertoli cell feeder layer, putative SSC colonies were divided into five groups: Group 1: No GDNF (Control); Group 2: 10 ng/mL GDNF; Group 3: 20 ng/mL GDNF; Group 4: 40 ng/mL GDNF; and Group 5: 100 ng/mL GDNF. On 15th day of culture, the relative expression of PLZF, TAF4B, THY1, BCL6B, UCHL1, and ID4 genes was examined to determine the GDNF concentration, at which the expression of majority of genes under study was upregulated.
Upon analysis it was found that the relative mRNA abundance of PLZF was significantly higher (p < 0.05) following supplementation of 20 ng/mL GDNF compared to the control (Fig. 4). Similarly, TAF4B and THY1 expressions were also significantly higher (p < 0.001) on addition of 20 ng/mL GDNF. However, BCL6B expression was highest at increase of 40 ng/mL GDNF concentration. No significant effect of GDNF supplementation was observed on UCHL1 and ID4 expression.

Effect of different concentrations of GDNF on quantitative expression of some SSC markers and self-renewal related genes on the 15th day of culture. Data are Mean of fold change ± SE. Bars with asterisks indicating significant differences from control (*p < 0.05; **p < 0.01;***p < 0.001). GDNF, Glial cell line-derived neurotrophic factor.
Effect of supplementation of GDNF in combination with LIF, bFGF, and CSF1 on quantitative expression of some SSC markers and self-renewal related genes
Based on its effect on gene expression in SSCs, 20 ng/mL concentration of GDNF was taken as the optimum concentration. This experiment was aimed at examining the effect of further supplementation with LIF, bFGF, or CSF1 on relative expression of some SSC markers and self-renewal related genes. Putative SSC colonies were divided into five different groups and subcultured for 15 days: Group 1: 20 ng/mL GDNF; Group 2: 20 ng/mL GDNF +1000 IU/mL LIF; Group 3: 20 ng/mL GDNF +10 ng/mL bFGF; Group 4: 20 ng/mL GDNF +1 ng/mL CSF1; and Group 5: 20 ng/mL GDNF +1000 IU/mL LIF +10 ng/mL bFGF +1 ng/mL CSF1. SSC culture supplemented with 20 ng/mL GDNF was taken as the control. On the 15th day of culture, the relative expression of PLZF, TAF4B, THY1, BCL6B, UCHL1, and ID4 was assayed and compared within the groups.
It was observed that supplementation of SSC culture with GFs (GDNF + LIF + bFGF + CSF1) resulted in a significantly higher (p < 0.001) expression of PLZF in comparison to the control (Fig. 5). Similarly, TAF4B and ID4 expressions were also higher (p < 0.05) in the presence of a cocktail of all growth factors in the culture media. UCHL1 expression was twofold higher (p < 0.05) following GDNF + LIF supplementation, while BCL6B overexpressed (p < 0.001) on addition of GDNF + bFGF. The growth factor supplementation had no significant effect on THY1 expression. From these observations it could be inferred that the optimized culture medium for buffalo SSC propagation would consist of the cocktail of all the growth factors.
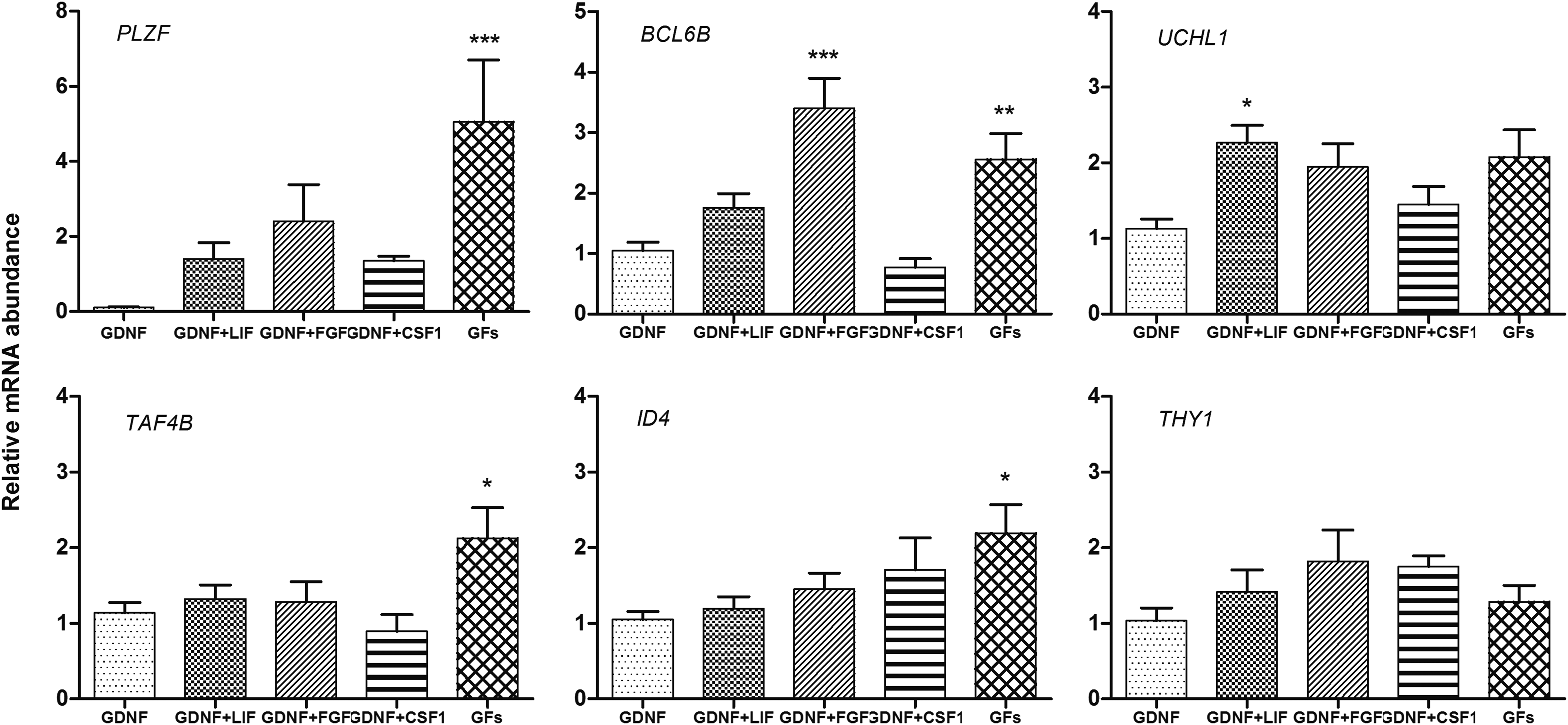
Effect of different combinations of bFGF, LIF, and CSF1 along with GDNF on quantitative expression of some SSC markers and self-renewal related genes on the 15th day of culture. GFs denote a combination of GDNF + bFGF + LIF + CSF1. Data are Mean of fold change ± SE. Bars with asterisks indicating significant differences from control (*p < 0.05; **p < 0.01;***p < 0.001). bFGF, basic fibroblast growth factor; LIF, leukemia inhibitory factor.
Discussion
In the present study, we successfully propagated putative buffalo SSC cultures under serum-free and growth factor defined conditions. Prepubertal buffalo calf testes (3–6 months of age) were used for SSC isolation because at this age the predominant germ cell type present in testis is type A spermatogonia (Ahmad et al., 2013; Aponte et al., 2006). The average cell yield after isolation was ∼1 × 106 cells/g of testis, and the viability ranged between 80% and 85%. Similar cell yield and viability after isolation were also reported in pig and cattle (Dirami et al., 1999; Izadyar et al., 2002).
After differential plating and discontinuous Percoll density gradient centrifugation, the putative SSCs were cultured on a mitomycin-C inactivated homologous buffalo Sertoli cell feeder layer, due to its ability to provide essential growth factors (GDNF and bFGF) and nutrients for maintenance and self-renewal of SSCs (Meng et al., 2000). The importance of Sertoli cell feeder layer in SSC proliferation and colony formation has already been elucidated for cattle and buffalo (Kadam et al., 2013; Oatley et al., 2004).
In the present study, the time duration for colony formation was 7–10 days, and the area of these colonies increased consistently up to day 15. Similar time duration for colony formation and expansion has been reported for SSCs cultured in media with less serum concentration (Aponte et al., 2006, 2008; Kanatsu-Shinohara et al., 2003). In contrast to the results of this study, the duration for SSC colony formation was between 15 and 18 days in our previous report (Kadam et al., 2013), in which 10% FBS was used as a supplement. The delayed colony formation rate could have been due to the detrimental effect exerted by serum on SSC self-renewal (Kubota et al., 2004b).
The use of high concentration of serum has also been shown to abrogate SSC expansion while causing an overwhelming growth of somatic cells (Bahadorani et al., 2012; Kanatsu-Shinohara et al., 2005). Aponte et al. (2008) also showed that the use of StemPro-34 serum free media for bovine SSC culture outperformed conventional MEM in terms of higher number of SSC colonies and spermatogonia within these colonies. In the present study, the use of stem cell specific media rather than DMEM may also have contributed to the earlier colony formation compared to our previous study (Kadam et al., 2013).
We observed a normal karyotype and high alkaline phosphatase activity in the cultured SSCs. The putative SSC colonies expressed DBA, PLZF, UCHL1, and THY1, which are specific markers for undifferentiated type A spermatogonia (Goel et al., 2010; Reding et al., 2010; Sharma et al., 2016). The colonies also expressed pluripotency related markers such as OCT-4, SOX-2, and NANOG, determining their stemness.
Although the Sertoli cell feeder layer supports the survival of SSCs and promotes their self-renewal by secreting GDNF and other required growth factors and cytokines, the importance of including additional GDNF in primary culture was reported previously in cattle (Aponte et al., 2006, 2008) and buffalo (Kadam et al., 2013). For establishment of primary SSC cultures, we used 40 ng/mL of GDNF and then proceeded with determining the ideal concentration required by comparing the self-renewal related genes and SSC markers expressed by SSC colonies supplemented with different concentrations. The duration of this experiment was chosen for 15 days based on our previous study (Kadam et al., 2013) in which, if not subcultured, putative buffalo SSC colonies degenerated after 18 days of culture.
The relative mRNA expression of PLZF, a transcription repressor expressed in undifferentiated spermatogonia with a role in stem cell self-renewal (Buaas et al., 2004; Kadam et al., 2013), was significantly higher (p < 0.05) following supplementation of 20 ng/mL GDNF. Similarly, the expression of TAF4B, a gonadal-specific component of TFIID essential for germ cell proliferation (Falender et al., 2005), and THY1, a unique surface marker for SSCs in cattle (Reding et al., 2010), was also higher (p < 0.001) in SSCs supplemented with 20 ng/mL GDNF. However, the expression of BCL6B, a GDNF dependent transcriptional factor essential for SSC self-renewal (Oatley et al., 2006, 2007), was higher in SSCs supplemented with 40 ng/mL GDNF.
Our results showed that the expressions of PLZF, TAF4B, and THY1 genes were upregulated by supplementing 20 ng/mL of GDNF. Similar to our results, Aoshima et al. (2013) also observed the reduction in GDNF concentration required for maintaining the growth of mouse SSC by supplementing KOSR along with BSA. Interestingly, Yu et al. (2014) demonstrated the successful in vitro culture of undifferentiated SSCs for 7 days in a stem cell specific media without supplementing additional GDNF. In this case, the Sertoli cell feeder layer may have provided the required GDNF for SSC growth and maintenance.
By assessing the results of the present study and of previous reports (Aoshima et al., 2013; Kadam et al., 2013; Yu et al., 2014), it is indicative that the ideal concentration of GDNF required for SSC growth and maintenance is highly variable and depends on factors like the type of media used (stem cell specific/basal), addition of serum/serum free supplements, and the inclusion of feeder layer.
Apart from GDNF, many other growth factors secreted from SSC niche (Sertoli, Leydig, and peritubular myoid cells) also contribute to the growth and self-renewal of SSCs. This has been demonstrated through in vitro studies conducted on mouse, bovine, and buffalo SSCs (Aponte et al., 2008; Kadam et al., 2013; Kanatsu-Shinohara et al., 2003), in which certain combinations of growth factors such as bFGF, LIF, and EGF along with GDNF were used. Recently, Oatley et al. (2009) observed that the addition of CSF1 to the culture media along with GDNF and bFGF significantly enhanced the self-renewal of mouse SSCs. However, it has remained elusive whether CSF1 influences the activity of SSCs in other species. Therefore, we evaluated the effects of different combinations of bFGF, LIF, and CSF1 along with GDNF on the expression of some SSC markers and self-renewal related genes.
The growth factors, as well as their concentrations, were determined from previous studies in same or different species (Kadam et al., 2013; Oatley et al., 2009; Yu et al., 2014). Our results showed that PLZF expression was highly upregulated (p < 0.001) on addition of GDNF + LIF + bFGF + CSF1. Similarly, expressions of ID4, a recently identified unique marker for mouse putative SSCs (Oatley et al., 2011), and TAF4B were also higher (p < 0.05) in the presence of all the growth factors. THY1 expression remained unaffected with supplementation of different combinations of growth factor.
Our results showed that the supplementation of a cocktail of GDNF, bFGF, LIF, and CSF1 upregulated the expression of PLZF, BCL6B, TAF4B, and ID4. Similar results were reported in our previous study (Kadam et al., 2013), in which supplementation of bFGF and EGF along with GDNF upregulated the expression of PLZF and TAF4B in buffalo putative SSCs, despite the lower survival time of the cultures. Yu et al. (2014) also reported a higher proliferation of buffalo SSCs by supplementing combination of GDNF, bFGF, and LIF in comparison to the individual effects.
In the present study, we were able to maintain the putative SSC colonies under serum-free but growth factor defined conditions for 28–30 days, which to our knowledge is the only such report in bubaline species. During this period, the colonies were passaged after every 15 days. After third passage since the primary culture, the colonies couldn't retain their compact structure and the cells diffused to develop a monolayer. Previously in buffalo, only Yu et al. (2014) could maintain the SSC culture for up to three passages. However, the duration of culture and time interval between two passages were not revealed.
In conclusion, we reported the use of serum free media, KO-DMEM supplemented with KOSR, to establish buffalo putative SSCs in vitro. We also observed that a concentration of 20 ng/mL of GDNF supported the self-renewal of SSCs under these culture conditions. Further supplementation of a cocktail of GDNF, bFGF, LIF, and CSF1 upregulated the majority of SSC markers and self-renewal related genes. Future studies aimed at developing an improved serum- and feeder-free culture system would facilitate in identifying the individual effects of growth factors on self-renewal of buffalo SSCs. Consequently, long-term in vitro culture of buffalo SSCs could be established.
Footnotes
Acknowledgments
The present work was funded by National Agriculture Innovation Project (NAIP) Grant to M.S.C. (C-2067 and 075) and S.K.S. (C 2–1-(5)/2007). The authors also acknowledge ICMR for providing PhD fellowships to Ankur Sharma.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.