
Abstract
Abstract
The establishment of porcine epiblast stem cells (pEpiSCs) and induced pluripotent stem cells (piPSCs) derived from diametrical derivations is of great importance in developing biomedical models. However, pEpiSCs and piPSCs have been technically much harder to culture than mouse embryonic stem cells, showing problematic properties such as spontaneous differentiation and apoptosis after cryopreservation. Therefore, we demonstrated that Y-27632 as a Rho-associated coiled-coil containing kinase inhibitor could prevent dissociated pEpiSCs and piPSCs from undesirable differentiation and apoptosis in cryopreservation protocols. pEpiSC 2, 8 lines, Sendai virus-induced pluripotent stem cells (Sev-iPSCs), and lentivirus-induced pluripotent stem cells were cultured with 10 μM Y-27632 before collecting dissociated cells retrieved from colonies using various enzymes. Dissociated single cells were transferred into freezing mediums (open pulled straw vitrification, STEM-CELLBANKER® (SCB), 10% dimethylsulfoxide in serum) for cryopreservation. The rates of viability and colony formation obtained from dissociated porcine stem cells after freezing/thawing were examined in the presence of Y-27632. The characteristics of pluripotency and in vitro differentiation were also examined in these stem cells treated with Y-27632 after cryopreservation. As a result, the viability and efficiency of colony formation of dissociated pEpiSCs (2, 8 lines) and Sev-iPSCs treated with 10 μM Y-27632 using the SCB cryopreservation protocol were significantly increased when compared with those of nontreated Y-27632 (p < 0.05). Pluripotency genes (OCT-3/4, NANOG, and SOX2) were positively expressed in Y-27632-treated porcine pluripotent stem cells. Also, in vitro differentiation of these stem cells was successfully induced in the presence of 10 μM Y-27632. These results indicated that treatment of Y-27632 for single-cell dissociation and the SCB cryopreservation protocol could facilitate handling porcine pluripotent stem cells and provide the widespread use of these stem cells.
Introduction
Embryonic stem cells (ESCs) derived from the inner cell mass of preimplantation embryos have the capacity for indefinite self-renewal and are able to differentiate into all cell types of the body. One of the two different populations of mouse ESCs (mESCs) has been generally produced from the epiblast of embryos at the gastrulation stage (Brons et al., 2007; Tesar et al., 2007; Vallier et al., 2009). Mouse epiblast stem cells (mEpiSCs) closely resemble human ESCs (hESCs), which grow larger than mESC colonies and constitute monolayer colonies of epithelial structure. In addition, these are dependent on the growth factor FGF2 and activin/nodal signaling rather than the leukemia inhibitory factor (LIF) to sustain their pluripotent status (Alberio et al., 2010; Hanna et al., 2010; Nichols and Smith, 2009).
Apart from this, Takahashi and Yamanaka (2006) first developed induced pluripotent stem cells (iPSCs) in mice by reprogramming differentiated cells with four exogenous genes (OCT-3/4, SOX2, cMYC, and KLF4) transferred into progenitor somatic cells using the integrating retroviral vector, thereby fulfilling the requirements of pluripotency. However, the reprogramming genes integrated into the differentiated cells remain expressed persistently in iPSCs. The integrated exogenous genes must be effectively deleted or silenced in the reprogrammed cells. An alternative method used to avoid the use of integrating vectors has been required.
An animal model is closely related to human than mice as a source of stem cells for therapeutic and regenerative medicine (Klimanskaya et al., 2008; Thomson et al., 1998; Trounson and Pera, 2001). Therefore, the pig is an excellent model for developing therapeutic tools due to the anatomical and physiological similarities with human (Kobayashi et al., 2017). Furthermore, genetic modification technologies have been consistently developed for modeling human disease in this species (Klymiuk et al., 2016). For a couple of decades, there have been many attempts to establish ESCs or EpiSCs from swine for degenerative human disease (Brevini et al., 2007; Chen et al., 1999; Evans et al., 1990; Haraguchi et al., 2012; Hochereau-de Reviers and Perreau, 1993; Li et al., 2004; Moore and Piedrahita, 1997; Notarianni et al., 1990; Siriboon et al., 2015; Talbot et al., 1993; Wianny et al., 1997).
However, germ line transmission in the pig has not been reported yet, and the characterization of pluripotent markers in putative porcine ESC or EpiSC lines has not appeared uniformly. On the contrary, several researchers have reported the construction of porcine iPSCs (piPSCs) based on variations of the technology introduced first in mice (Esteban et al., 2009; Ezashi et al., 2009). Nowadays, piPSCs provide the promise of permitting pluripotent stem cells to be derived from animal models important in biomedical research.
In porcine pluripotent stem cells, the establishment of efficient single-cell dissociation and cryopreservation protocol of pluripotent stem cells needs to remove the loss in viability, spontaneous differentiation for cell passaging, and cell line expansion and preservation. The vulnerability of porcine pluripotent stem cells highly similar with the characteristics of hESCs was easily disrupted by single-cell dissociation and cryopreservation (Kim and Hyun, 2016). Until now, only partially dissociated colonies or small aggregates of porcine pluripotent stem cells have been alternatively performed for single-cell dissociation and cryopreservation.
However, the colonies of these stem cells must be dispersed into single cells to enhance the efficiency of stem cell research. Although the use of serum and dimethylsulfoxide (DMSO) as the most common cryoprotectant provides for conventional freezing and thawing methods, most of the hESCs have resulted in differentiation and apoptosis (Reubinoff et al., 2001; Richards et al., 2004). Therefore, with consideration for limited loss in viability and low differentiation rates, the establishment of a novel cryopreservation protocol for porcine pluripotent stem cells would be quite important for the widespread use of these cell lines.
Activation of a Rho-associated coiled-coil containing kinase (ROCK) prevents cell adhesion to tissue culture plates and induces apoptosis. It was reported that a specific inhibitor of ROCK (Y-27632) enhances cell adhesion to the substrate, viability, and in vitro differentiation in human pluripotent stem cells (Ohgushi et al., 2010; Okumura et al., 2009). Generally, Rho family proteins regulate a variety of cell behaviors such as cell migration, cell adhesion to substrate, cell-to-cell communication, as well as inhibit apoptosis (Bustelo et al., 2007; Riento et al., 2003).
In addition, Y-27632-treated hESCs grown at low density were positively expressed for the undifferentiated state markers such as E-cadherin, OCT-3/4, and SSEA-4 (Watanabe et al., 2007). After five low-density passages, Y-27632-treated hESCs have retained the capacity to differentiate into neural cells, mesodermal cells, and endodermal precursors in vitro (Watanabe et al., 2007).
Finally, unlike somatic cells, hESCs have a shortened G1cell cycle phase (2–3/16–18 hours) thorough autocrine mechanisms (Becker et al., 2006, 2007). Therefore, self-renewal of hESCs is premitotically committed to initiate a new round of cell division. However, differentiation of hESCs alters a lengthened G1 cell cycle phase within 72 hours, suggesting that the abbreviated G1 phase of the hESCs is functionally associated with the pluripotency state (Becker et al., 2010a, 2010b).
Therefore, in the present study, we have examined that Y-27632 as an ROCK inhibitor would improve viability, efficiency of colony formation, retention of pluripotency, competence of in vitro differentiation, and stability of dissociated porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sendai-virus iPSCs [Sev-iPSCs], and lentivirus iPSCs [Lenti-iPSCs]), providing animal models important in biomedical research.
Materials and Methods
All chemicals and reagents were purchased from Sigma-Aldrich Co. (St. Louis, MO) unless otherwise mentioned.
Culture and maintenance of porcine pluripotent stem cells
Four porcine pluripotent stem cells [epiblast stem cell (pEpiSCs) 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs] were used as experiment groups for the present study. Porcine epiblast stem cells lines were derived from epiblasts, which isolated from in vivo porcine embryos of spherical or ovoid type collected after 9–9.5 days postinsemination (d.p.i.). These stem cells lines were maintained on mitomycin C-inactivated mouse embryonic feeder cells (iMEFs) in Dulbecco's modified Eagle's medium (DMEM/F12; GIBCO) containing 1% minimal essential medium (MEM) nonessential amino acids, 1% penicillin/streptomycin, 2 mM
The Sev-iPSC line was generated from porcine fetal fibroblasts (PFFs) using the CytoTune™-iPS Reprogramming Kit (Life Technologies) following the manufacturer's instructions. Sev-iPSCs were cultured on iMEFs in knockout DMEM/F12 (Thermo Fisher Scientific) containing 1% MEM nonessential amino acids, 1% penicillin/streptomycin, 2 mM
Cell dissociation, colony formation, and embryoid body aggregation
To determine dissociation efficiency of porcine pluripotent stem cells, the porcine stem cells were incubated with 0.05% Trypsin/EDTA, 0.05% Accutase, 1 mg/mL collagenase (GIBCO), 1 mg/mL dispase (GIBCO), or a mixture of 1 mg/mL collagenase/dispase for exposure times, respectively. Single dissociated cells of porcine pluripotent stem cells were counted using a hematocytometer (Marienfeld Superior™, Germany). Dissociated porcine stem cells were seeded (5 × 103 cells/cm2) onto iMEFs and then calculated for the percentage of colony formation.
For the production of embryoid bodies (EBs), dissociated porcine pluripotent stem cells were collected in differentiation medium (DMEM/F12 supplemented with 1% MEM nonessential amino acids, 1% penicillin/streptomycin, 2 mM
Y-27632 treatment and colony formation of dissociated porcine pluripotent stem cells
To increase the survival of dissociated porcine pluripotent stem cells, colonies of the porcine stem cells were incubated with 10 μM Y-27632 (a commonly used working concentration) in culture medium before and after cellular dissociation. The colonies of stem cells were detached from iMEFs treated with 0.05% Accutase at 39°C for 5 minutes. For removal of mixing iMEFs, the detached colonies were suspended on a gelatin-coated dish in culture medium at 39°C for 30 minutes. The colonies were dissociated into single cells by gentle pipetting and centrifugation, and then washed with culture medium. Dissociated pluripotent stem cells were seeded at low density (5 × 103 cells/cm2) onto fresh iMEFs. The number of colony formations was counted after culture of seeded cells on day 3.
Cryopreservation of porcine pluripotent stem cells
Dissociated porcine pluripotent stem cells were cryopreserved using several freezing/thawing systems [open pulled straw (OPS) vitrification method (Reubinoff et al., 2001), serum- and xeno-free chemically defined freezing methods (Holm et al., 2010), and conventional slow freezing methods (Freshney, 1994)]. In the case of OPS, the porcine stem cells were cryopreserved by using vitrification solution-I and then stored in liquid nitrogen. On the occasion of serum- and xeno-free chemically defined freezing method, the stem cells were cryopreserved by using STEM-CELLBANKER™ (SCB) (ZENOAQ, Fukushima, Japan) containing 10% DMSO and a high polymer.
In the conventional slow freezing method, porcine stem cells were cryopreserved using freezing medium (10% DMSO in serum). The dissociated porcine stem cells suspended in OPS vitrification method, STEM-CELLBANKER™, and 10% DMSO freezing medium were transferred into cryo-vial (Nunc, Roskilde, Denmark) placed in a Nalgene® Cryo 1°C Mr. Frosty™ Freezing Container (Nunc, Hereford, United Kingdom) and then stored at −80°C. Cryo-vials were stored in liquid nitrogen the next day.
Y-27632 treatment, viability measurement, and colony formation in freezing/thawing systems
To examine the protective effect of Y-27632 on viability and colony formation of dissociated porcine pluripotent stem cells in freezing/thawing systems, the stem cells were incubated in the presence of 10 μM Y-27632 before freezing. After thawing, viability of the dissociated stem cells was counted by using 0.8 mM Trypan blue staining and a hematocytometer. After plating these stem cells, cells were incubated in the presence of 10 μM Y-27632. The colony formation was determined as ratio between the number of colonies generated in culture medium for 3 days and the total number of initially seeded cells (5000 cells).
RNA extraction, polymerase chain reaction, and quantitative real-time polymerase chain reaction
The effect of Y-27632 on pluripotency in porcine stem cells obtained after freezing and thawing was examined using RT-polymerase chain reaction (PCR) and quantitative real-time PCR (qPCR) (Tables 3 and 4). Total RNAs of porcine stem cells were extracted using Agilent RNA 6000 Nano Kit (Agilent Technologies, Lubbock) and RNeasy Plus Mini Kit (Qiagen, Valencia). cDNA was synthesized using a RevoScript™ RT Premix (iNtRON Biotechnology, Inc., Korea) and total RNA and cDNA were measured using MaestroNano® Spectrophotometer (MAESTROGEN, Inc.).
Quantitative Real-Time Polymerase Chain Reaction Primer Lists Used in This Study
GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
RT-Polymerase Chain Reaction Primer Lists Used in This Study
RT-PCR was performed by Maxime PCR Premix (iNtRON Biotechnology, Inc.) and conditions of RT-PCR were as follows: predenaturation for 10 minutes at 95°C, denaturation for 30 seconds at 94°C, annealing at a temperature specific for each primer set for 40 seconds, extinction for 60 seconds at 72°C, and final extension for 10 minutes at 72°C for 40 cycles using Pro s6325 (Eppendorf, Germany). These PCR products were analyzed by 1.5% agarose gel in 1 × TAE buffer.
On the contrary, qPCR was performed using SYBR Green (TOYOBO Ltd., Osaka, Japan) on the CFX connect™ Real-time PCR Detection system (BIO-RAD) and conditions were as follows: predenaturation for 30 seconds at 95°C, denaturation for 5 seconds at 95°C, annealing at a temperature specific for each primer set for 10 seconds, and extinction for 15 seconds at 72°C for 40 cycles. Data analysis was used for 2−ΔΔCt method and gene expression was normalized relative to reference gene (GAPDH).
Immunocytochemistry and flow cytometry analysis
For immunocytochemistry, porcine pluripotent stem cells treated with 10 μM Y-27632 after freezing/thawing were fixed by 4% paraformaldehyde for 15 minutes and then incubated with blocking solution containing 5% bovine serum albumin for 1 hour at room temperature. The stem cells were then incubated with primary antibodies under the following conditions at 4°C overnight: OCT-3/4 (1:100; Santa Cruz Biotechnology, Santa Cruz), NANOG (1:100; Abcam, Cambridge), SOX2 (10 ng/mL; R&D System, Minneapolis, MN), and histone H3 at lysine 27 (H3K27me3) (1:200; Abcam).
After overnight, the stem cells were incubated with secondary antibodies under the following conditions at room temperature for 1 hour: Alexa Fluor® 568 Donkey Anti-Goat IgG (1:200; Invitrogen, Carlsbad), Alexa Fluor 546 Goat Anti-Rabbit IgG (1:200; Invitrogen), and Alexa Fluor 555 Donkey Anti-Mouse IgG (1:200; Invitrogen). To indicate the nuclei in cells, 5 μg/mL of Hoechst 33342 (Life Technologies, Carlsbad) was treated at room temperature for 10 minutes. All images were examined using a fluorescence microscope (LEICA DM 2500; Leica, Germany).
Cell cycle, population doubling, and karyotyping analysis
For cell cycle analysis, porcine pluripotent stem cells incubated with Y-27632 after freezing/thawing were dissociated with 0.05% Trypsin/EDTA or Accutase at 39°C for 5 minutes. The porcine stem cells separated to single cells were incubated in 50 μL/mL of propidium iodide (PI) staining solution (0.1 mg/mL RNase A, Triton X-100 in phosphate-buffered saline) at 37°C for 40 minutes. The samples were analyzed using BD FACS Calibur flow cytometer (BD Biosciences, Becton Dickinson) and CellQuest software. Maximum excitation of PI bound to DNA was at 483 nm and emission was at 635 nm. The results were analyzed using FlowJo software, version 10.0.7 (TREE STAR, Inc.). On the contrary, the population doubling time (PDT) of dissociated porcine stem cells was calculated using the duration × log (2)/log (final concentration) − log (initial concentration) formula at each passage.
For karyotyping analysis, porcine stem cells were cultured with 10 μL/mL of colcemide (Biological Industries Israel Beit Haemek Ltd., Israel) in culture medium at 37°C for 1 hour and then harvested using 0.05% Trypsin/EDTA. Harvested single cells were incubated with hypotonic solution (0.4% NaCl and 0.4% KCl in H2O) at 39°C for 6 minutes and then fixed in fixative (3:1 methanol:acetic acid). The cell pellet suspended in 1 mL of fixation solution was dropped onto cold slide and then dried. The chromosomes of metaphase stage stained with Giemsa were patterned by standard G-banding techniques.
Statistical analysis
All experiments were conducted in triplicate in three-four independent cultures considered independent experiments. Table 1 and Figure 1A, 2B, and 6B data were analyzed with Bonferroni's post hoc test using the one-way ANOVA test in SPSS (SPSS, Inc.). Table 2 and Figure 1B, 3B, and 5A data were analyzed with the least significant difference post hoc test using the generalized linear model in SAS version 9.3 (SAS Institute, Inc.). A probability of p < 0.05 was considered significant.

Effect of Y-27632 on colony formation derived from dissociated porcine pluripotent stem cells. Porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) were dissociated into single cells with treatment of 0.05% Accutase for 5 minutes.
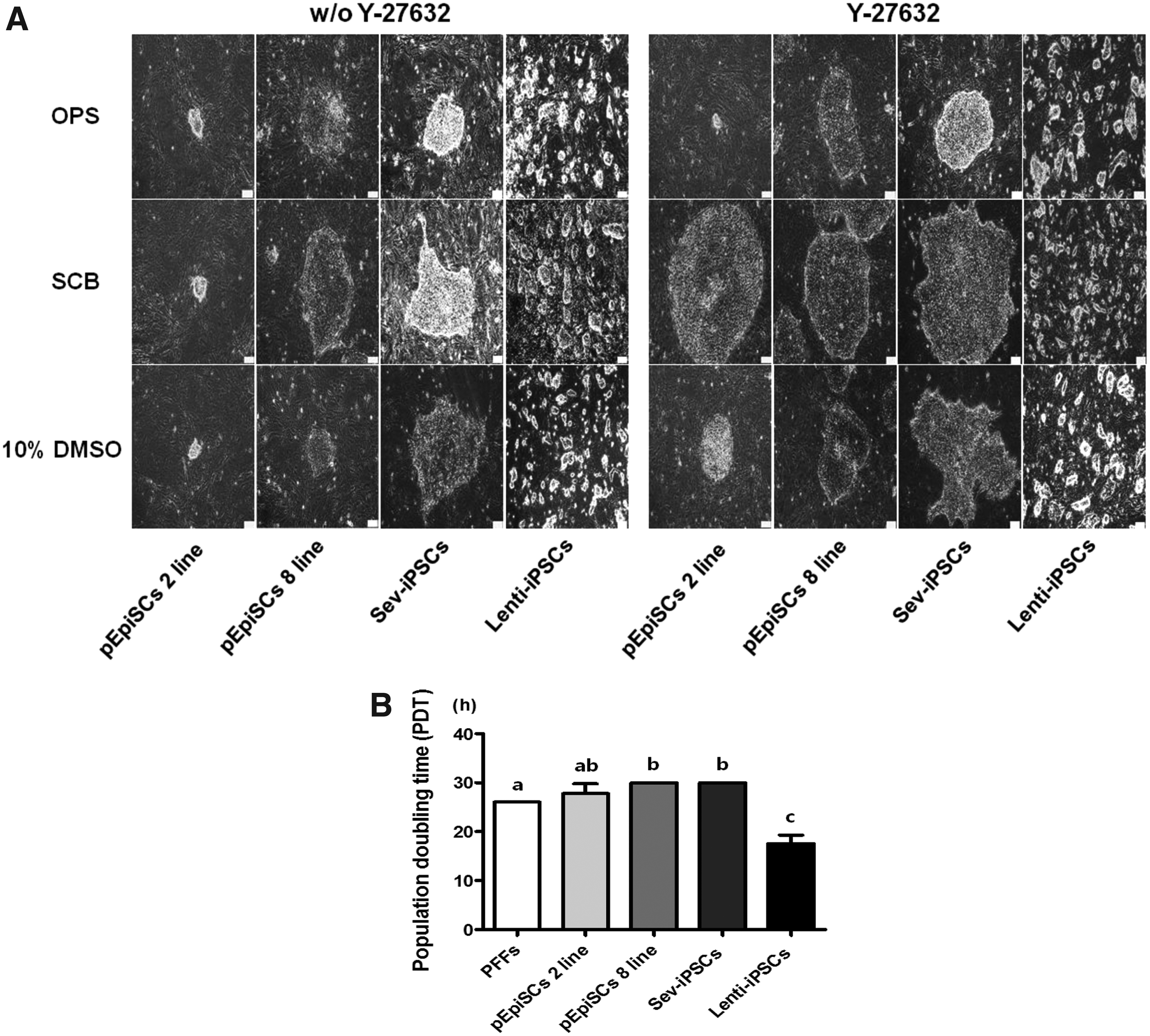
Effect of Y-27632 on colony formation and PDT of dissociated porcine pluripotent stem cells after treatments of freezing/thawing.
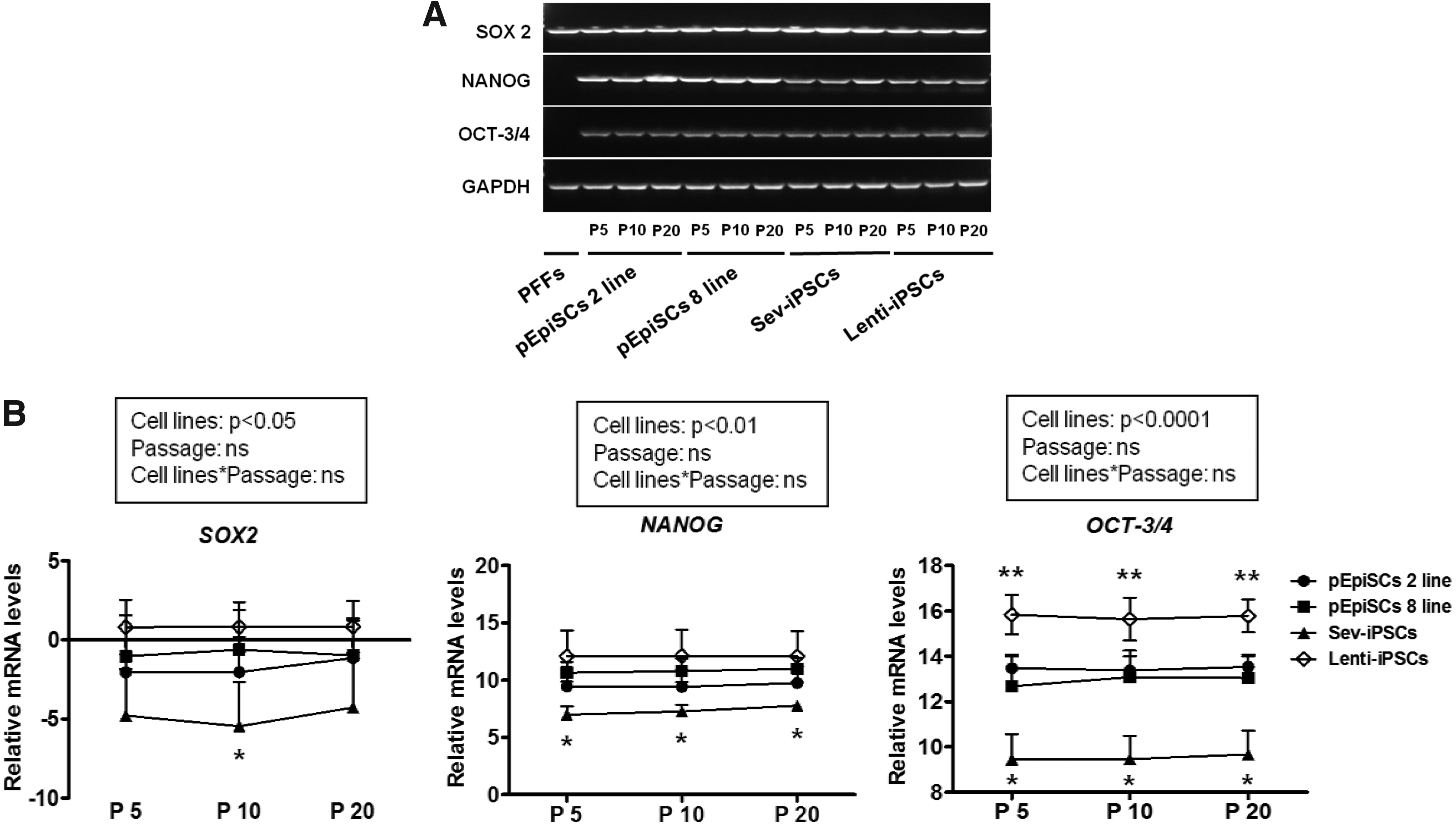
Expression of pluripotency marker genes in porcine pluripotent stem cells treated with Y-27632.
Effect of Enzymes on Single-Cell Dissociation for Colony Formation of Porcine Pluripotent Stem Cells
Approximately 50,000 single cells of porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) were dissociated by using enzymes with the following conditions: 0.05% Trypsin for 5 minutes, 0.05% Accutase for 5 minutes, 1 mg/mL collagenase for 60 minutes, 1 mg/mL dispase for 60 minutes, and 1 mg/mL mixture of collagenase/dispase for 60 minutes, respectively. The recovery rate of dissociated single cells was counted using a hematocytometer. The rate of colony formation was calculated as the percentage of colonies formed as per the number of single cells seeded (5000 cells) on day 3. All values are represented as mean ± SD. a,b,cValues in same columns with different superscripts are significantly different (p < 0.05) with one-way ANOVA and Bonferroni's post hoc test.
Lenti-iPSC, lentivirus-induced pluripotent stem cell; Sev-iPSC, Sendai virus-induced pluripotent stem cell; SD, standard deviation.
Effect of Y-27632 on Viability and Colony Formation of Dissociated Porcine Pluripotent Stem Cells After Freezing/Thawing
Porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) were frozen using OPS vitrification, SCB), and 10% DMSO in serum in the absence or presence of 10 μM Y-27632. The number of single cells that survived after thawing was counted using a hematocytometer. The number of colonies formed after freezing/thawing of porcine pluripotent stem cells was counted. The rate of colony formation was calculated as the percentage of colonies formed as per the number of survived single cells seeded (5000 cells) for 72 hours. Vehicle: water, −: without Y-27632, +: with Y-27632. Values are represented as mean ± SE. a,b,c,d,e are different (p < 0.05) groups by GLM followed by LSD post hoc test.
DMSO, dimethylsulfoxide; GLM, generalized linear model; LSD, least significant difference; OPS, open pulled straw; SCB, STEM-CELL BANKER®.
Results
Effect of enzymes on single-cell dissociation of porcine pluripotent stem cells
To examine the dissociation of porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) into single cells, 0.05% Trypsin/EDTA, 0.05% Accutase, 1 mg/mL collagenase, 1 mg/mL dispase, or the mixture of 1 mg/mL collagenase/dispase was treated to these stem cells for individual times. As shown in Table 1, the number of porcine pluripotent stem cells recovered after treatment with 0.05% Trypsin/EDTA or 0.05% Accutase for 5 minutes was significantly increased compared with 1 mg/mL collagenase, 1 mg/mL dispase, and the mixture of 1 mg/mL collagenase/dispase for 60 minutes (p < 0.05).
However, the efficiency of colony formation from dissociated porcine stem cells (pEpiSC 2, 8 lines and Sev-iPSCs) treated with 0.05% Accutase for 5 minutes was significantly improved compared with 0.05% Trypsin/EDTA, 1 mg/mL collagenase, 1 mg/mL dispase, and the mixture of 1 mg/mL collagenase/dispase (p < 0.05) (Table 1). In addition, the rate of colony formation derived from dissociated Lenti-iPSCs was significantly higher compared with pEpiSC 2, 8 lines and Sev-iPSCs when treated with 0.05% Accutase for 5 minutes (p < 0.05) (Fig. 1A).
ROCK inhibitor, Y-27632, improves the efficiency of colony formation of dissociated porcine pluripotent stem cells
To determine the efficiency of colony formation of dissociated porcine stem cells, 10 μM Y-27632 as an ROCK inhibitor was added to the culture medium of dissociated pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs for 3 days. As shown in Figure 1B, using 10 μM Y-27632 treatment, pEpiSC 2, 8 lines and Sev-iPSCs significantly increased the rate of colony formation when compared with that in vehicle (water) and the absence of Y-27632 (p < 0.05). However, there was no difference in the rate of colony formation of Lenti-iPSCs between the absence and presence of Y-27632. The addition of 10 μM Y-27632 to porcine pluripotent stem cells has shown undifferentiated morphological features such as well-defined borders and small cells with high nuclear:cytoplasm ratios when compared with those of Y-27632-untreated porcine stem cells (Fig. 2A). Interestingly, the morphology of pEpiSC 2, 8 lines and Sev-iPSCs was highly similar to that of hESCs, whereas the morphology of Lenti-iPSCs was similar to that of mESCs on day 3.
Effect of Y-27632 on viability and colony formation of dissociated porcine pluripotent stem cells after various freezing/thawing protocols
We also evaluated the viability and efficiency of colony formation of dissociated porcine pluripotent stem cells in the presence of 10 μM Y-27632 following different cryopreservation methods. As seen in Table 2, there was no significantly difference in the number of viable single cells obtained from dissociated porcine pluripotent stem cells between OPS vitrification, STEM-CELLBANKER® (SCB), and 10% DMSO in serum without reference to Y-27632.
However, the rate of colony formation of dissociated porcine stem cells after SCB freezing/thawing protocol was significantly enhanced compared with that of OPS vitrification and 10% DMSO in serum protocols (p < 0.05) (Table 2). Noteworthy is the rate of colony formation of dissociated single cells following any freezing/thawing method, which was the highest in Lenti-iPSCs among all porcine pluripotent stem cells. This might be strongly related with the PDT in the porcine pluripotent stem cells. As shown in Figure 2B, PDT of Lenti-iPSCs treated with 10 μM Y-27632 was significantly shortened compared with other porcine pluripotent stem cells (pEpiSC 2, 8 lines and Sev-iPSCs) (p < 0.05).
On the contrary, the morphology of porcine pluripotent stem cells treated with 10 μM Y-27632 has exhibited more extended and well-defined border features compared with Y-27632-untreated porcine stem cells (Fig. 2A). The size of colony derived from dissociated porcine pluripotent stem cells following the SCB freezing/thawing protocol was highly increased when compared with that after the OPS vitrification method and 10% DMSO in serum freezing/thawing protocols. Unlike the feature of pEpiSC 2, 8 lines and Sev-iPSCs, the morphology of Lenti-iPSCs has shown dome-shaped colonies.
Y-27632 induces expression of pluripotency genes in porcine pluripotent stem cells
We next analyzed that the expressions of pluripotency marker genes (OCT-3/4, NANOG, and SOX 2) of porcine pluripotent stem cells sustained in the presence of 10 μM Y-27632 with different passage courses (5, 10, and 20) following the SCB freezing/thawing protocol using qPCR and immunocytochemistry. All pluripotency marker genes mentioned above were positively expressed in porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs), whereas these genes apart from SOX2 were not expressed in differentiated cells (PFFs) (Fig. 3A). Although the expression of pluripotency marker genes in Lenti-iPSCs was highly expressed compared with the other porcine pluripotent stem cells (pEpiSC 2, 8 lines and Sev-iPSCs), Sev-iPSCs had a significantly lower expression compared with pEpiSC 2, 8 lines and Lenti-iPSCs (p < 0.05) (Fig. 3B).
Consistent with these observations, all pluripotency marker proteins (OCT-3/4, NANOG, and SOX2) are positively expressed in 10 μM Y-27632-treated porcine pluripotent stem cells (Fig. 4A). The trimethylation of H3K27me3 as a differentiated marker was strongly induced in the porcine fibroblasts compared with the negative expression in colonies of porcine pluripotent stem cells (Fig. 4B).

Expression of pluripotency marker genes in porcine pluripotent stem cells treated with Y-27632.
Y-27632 enhances embryonic body formation aggregated from dissociated porcine pluripotent stem cells
To examine the efficiency of embryoid bodies (EB) formation aggregated from dissociated porcine pluripotent stem cells in the presence of Y-27632, porcine stem cells were cultured in differentiation medium and then aggregated in hanging drop at a seeding density of 1 × 103 cells/drop for 3 days. As shown in Figure 5A, the rate of EB formation aggregated from dissociated pEpiSC 2, 8 lines and Sev-iPSCs treated with 10 μM Y-27632 was significantly increased rather compared with Y-27632-untreated porcine pluripotent stem cells (p < 0.05).

Effect of Y-27632 on EB formation derived from porcine pluripotent stem cells aggregated by a hanging drop protocol.
However, there was difference in the rate of EB formation aggregated from dissociated Lenti-iPSCs, with or without Y-27632. The EBs aggregated from dissociated pEpiSC 2, 8 lines and Sev-iPSCs represented more clearly delineated EB features, whereas Y-27632-untreated porcine pluripotent stem cells showed the morphology of poor clearly delineated EBs (Fig. 5B). Interestingly, Lenti-iPSCs did not show the difference in the morphology between with and without Y-27632 at all.
According to differentiation marker genes expressed commonly in primary embryonic germ layers (Vasa [primordial germ cell marker], Cardiac actin [mesoderm marker], Nestin [ectoderm marker] and GATA 6 [endoderm marker]), EBs aggregated from 10 μM Y-27632-treated pEpiSC 8 line and Lenti-iPSCs positively expressed all the differentiation marker genes such as Vasa, Cardiac actin, Nestin, and GATA 6 (Fig. 5C). However, EBs derived from Y-27632-treated pEpiSC 2 line did not show expression of Vasa, Cardiac actin, and Nestin genes. Similarly, Sev-iPSCs treated with Y-27632 did not induce expression of Vasa, Cardiac actin, and GATA 6 at all. All porcine pluripotent stem cells did not show expression of Vasa, Cardiac actin, Nestin, and GATA 6 genes, except the expression of GATA 6 in pEpiSC 2 line and Lenti-iPSCs.
The effect of Y-27632 on cell cycle and chromosome in porcine pluripotent stem cells
The cell cycle of porcine pluripotent stem cells in the presence of Y-27632 was analyzed after cryopreservation using FACS and FlowJo. Almost differentiated porcine cells (PFFs) were placed in G0/G1 stage, whereas the porcine stem cell lines treated with 10 μM Y-27632 were significantly decreased in G0/G1 stage but increased in G2/M phase (p < 0.05) (Fig. 6A, B).
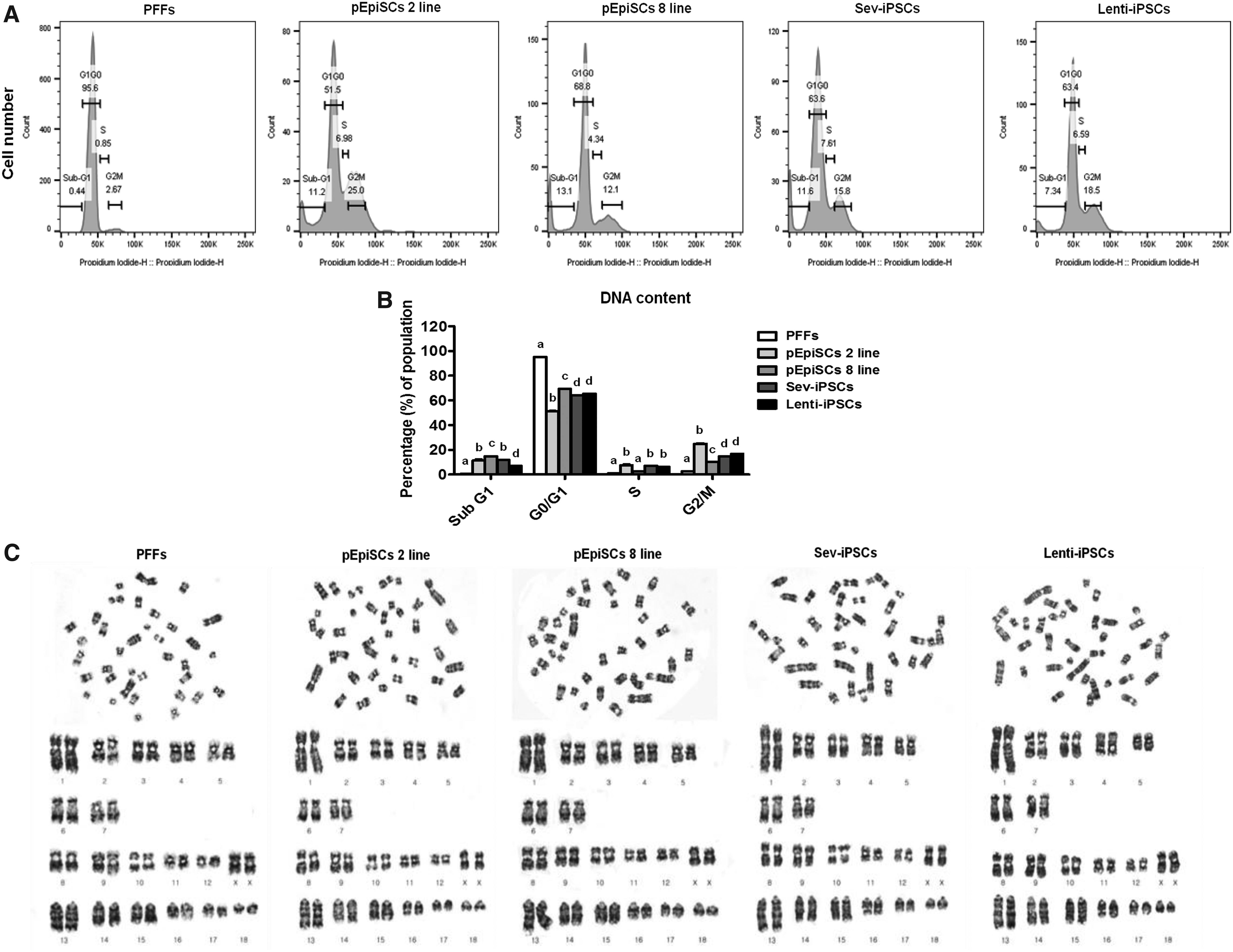
Flow cytometric analysis of cell cycle phase-specific population and karyotyping analysis of porcine pluripotent stem cells.
On the contrary, karyotyping analysis was performed in all porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) at passage 20 following treatments of Y-27632 and SCB freezing/thawing protocol to demonstrate the presence of chromosomal normalities. The results showed porcine stem cells, a normal female 38, XX karyotype (Fig. 6C).
Discussion
Pluripotency of hESCs and iPSCs (hiPSCs) is generally sustained when these cells are formed as the epithelial or flattened structure of colony. If hESCs or hiPSCs are dispersed into single cells, they encounter spontaneous differentiation and programmed cell death (apoptosis) (Amit et al., 2000; Thomson et al., 1998). Therefore, the dissociation of colony-formed stem cells into single cells using trypsinization results in the loss of pluripotency and low survival rate. In the present study, it was also demonstrated that dissociation of porcine epiblast stem cells (pEpiSC 2, 8 lines) and Sev-iPSCs using enzymes such as trypsin, accutase, collagenase, and dispase eventually results in poor colony formations except from Lenti-iPSCs (0%–0.2% vs. 1.1%–3.2%).
To resolve the problem of dissociated-induced apoptosis in porcine pluripotent stem cells, we used Y-27632 as a specific of Rho-associated protein kinase (ROCK inhibitor), which presented a high efficiency of colony formation and induced clearly delineated and compact colony features in human pluripotent stem cells (Martin-Ibañez et al., 2008; Watanabe et al., 2007). By contrast, Y-27632-untreated stem cells exhibited an unclear demarcation to a surrounding of epithelial colony. In addition, it was proved that Y-27632 treatment to human pluripotent stem cells inhibits ROCK activation and causes Rac1 activation, thereby preventing apoptosis and enhancing their culture efficiency (Ohgushi et al., 2010; Okumura et al., 2016).
In the present study, the addition of 10 μM Y-27632 to porcine pluripotent stem cell enhanced the efficiency of colony formation and induced clearly delineated and compact colony features.
The conventional cryopreservation (slow-freezing and rapid-thawing) methods commonly used for gametes (oocytes and sperms) and preimplantation embryos are not suitable for hESCs and hiPSCs (Reubinoff et al., 2001; Richards et al., 2004). This limited success of cryopreservation for human stem cells might be caused by being vulnerable to the damage inflicted by the freezing procedure. Although this method provided a high efficiency for the cryopreservation of mESCs, the survival rate of human pluripotent stem cells remained still low (Kuleshova and Lopata, 2002; Robertson, 1987). Therefore, human pluripotent stem cells have been usually cryopreserved as small clumps to avoid dissociation-induced apoptosis.
A decade ago, it was reported that the addition of Y-27632 to freezing and thawing mediums improves the efficiency of colony formation and increases the survival rate for human pluripotent stem cells (Li et al., 2008; Martin-Ibañez et al., 2008). Our results also presented that the addition of 10 μM Y-27632 retains a high colony formation of frozen/thawed porcine pluripotent stem cells with a clear and larger demarcation to a surrounding of epithelial flattened, but low colony formation of Y-27632-untreated stem cells with an unclear and smaller demarcation to a surrounding of flattened cells.
In general, STEM-CELLBANKER® is a commercial cell-freezing medium that allows for stable cryopreservation with a high viability of cells after the freeze/thaw procedure. In the present study, frozen/thawed porcine pluripotent stem cells (pEpiSC 2, 8 lines and Sev-iPSCs) treated with both Y-27632 and STEM-CELLBANKER® increased sizes of colony compared with the OPS vitrification method or 10% DMSO in serum-treated stem cells. This result was highly similar to a previous study that the addition of Y-27632 to freezing and post-thawing mediums increases the average size and the number of hESCs colonies (Sivasubramaniyan et al., 2010).
However, Lenti-iPSCs being highly similar to the characteristics of mESCs exhibited no difference in the STEM-CELLBANKER® protocol. Interestingly, PDT of Lenti-iPSCs was reduced ∼1.5-fold compared with that for the other porcine pluripotent stem cells (17 hours vs. 30 hours). This may be related to the particular characteristics of Lenti-iPSCs, which typically grow as compact and domed colonies that are insensitive to single-cell dissociation.
The pluripotency state of dissociated porcine stem cells following a frozen/thawed protocol must be maintained before the inducement of differentiation. Generally, the transcription factors (OCT-3/4, NANOG, and SOX2) as the key regulators of pluripotency in the mouse have maintained the characteristics of ESCs by binding to their own promoters, thereby forming core transcriptional regulatory circuitry (Avilion et al., 2003; Chambers et al., 2003; Schöler et al., 1990).
Similar to the result of hESCs (Martin-Ibañez et al., 2008), our results have shown that the level of pluripotency-related gene (OCT-3/4, NANOG, and SOX2) expression in 10 μM Y-27632-treated porcine pluripotent stem cells (pEpiSC 2, 8 lines, Sev-iPSCs, and Lenti-iPSCs) at a range of time points (passages 5, 10, and 20) after cryopreservation retains sustainably. In the case of porcine epiblasts, NANOG expression was also dependent on activin A signaling (Alberio et al., 2010) and was maintained in a subset of posterior epiblast cells presumed to become primordial germ cells (Wolf et al., 2011). In the present study, the expression of NANOG in pEpiSC 2, 8 lines, Lenti-iPSCs, and Sev-iPSCs was highly induced, whereas the differentiated cells did not express it.
On the contrary, characteristics of inactive X-chromosome are the activation of trimethylation of H3K27me3 as a repressive mark that impedes transcription by excluding the binding of RNA-pol II (Plath et al., 2003). Our results have shown that in the porcine pluripotent stem cells, H3K27me3 was negatively expressed when compared with the differentiated cells (PFFs), thereby confirming the undifferentiated state of the porcine pluripotent stem cells. This result was similar to a previous study that Y-27632 can spatiotemporally alter the balance between pluripotency and early differentiation events in hESCs (Sivasubramaniyan et al., 2010).
According to previously reported experiments, the addition of Y-27632 to suspension culture medium of dissociated human pluripotent stem cells induced the formation of EB-like aggregates and then converted them into telencephalic or retinal progenitors (Osakada et al., 2009; Watanabe et al., 2007). Our results also demonstrated that the treatment of 10 μM Y-27632 significantly increased the formation of EBs in the suspension culture medium of pEpiSC 2, 8 lines and Sev-iPSCs.
However, there was no difference in in vitro differentiation in Lenti-iPSCs between the absence and presence of Y-27632. Based on the distinct properties of stem cells, Lenti-iPSCs such as the naive-type stem cells were not affected in their potential to differentiate toward EB with or without Y-27632, whereas the other porcine stem cells were thoroughly affected to differentiate into EB in vitro with Y-27632. Therefore, the use of Y-27632 is able to enhance the reproducible ability and the efficiency of in vitro differentiation induction in porcine pluripotent stem cells (pEpiSC 2, 8 lines and Sev-iPSCs) like the prime-type stem cell.
Finally, mESCs usually retain their pluripotency state, which is associated with cell cycle regulation characterized by a shortened G1 phase (Coronado et al., 2013; Savatier et al., 1996). Consistent with the above phenomenon, hESCs have also shown a very short G1 phase (2–3 hours) of an abbreviated cell cycle (16–18 hours) (Becker et al., 2006, 2007; Ghule et al., 2011). However, our results have differently exhibited that the cell cycle of porcine pluripotent stem cells treated with 10 μM Y-27632 was arrested at G0/G1 phase (51.5 ± 0.1–69.4 ± 0.6) and G2/M phase (10.2 ± 1.7–25.0 ± 0.4), whereas the cell cycle of the differentiated cell was mainly arrested at G0/G1 phase (95.3 ± 0.4). The dissociation and cryopreservation of porcine pluripotent stem cells did not alter chromosomal abnormalities at all.
In conclusion, we have established efficient protocols for dissociation and cryopreservation of porcine pluripotent stem cells derived from different derivations. These techniques may facilitate handling porcine pluripotent stem cells and provide the widespread use of these stem cells for research activities in domestic animals.
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea funded by the Korean Government (2017R1A2B1011438), Republic of Korea. S.-K.B. and Y.-S.C. were supported by a scholarship from the BK21Plus Program, Ministry of Education, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.