
Abstract
Abstract
Embryonic stem cells (ESCs) are characterized by their ability to self-renew and their potential to differentiate into any cell type. Therefore, identification of novel molecular markers to verify the pluripotent status of mouse ESCs (mESCs) is of great significance. Kinase D interacting substrate of 220 kDa (Kidins220)/ankyrin repeat-rich membrane spanning (ARMS) plays a crucial role in the integration of growth factor receptor pathways during embryonic development. However, the role of Kidins220/ARMS in ESCs is still unknown. To elucidate the effects of Kidins220/ARMS on ESCs, we performed a knockdown of the Kidins220/ARMS gene by RNA interference. To our surprise, downregulation of Kidins220/ARMS did not alter the pluripotent state of mESCs. In contrast, it was essential for the proliferation and survival of ESCs. Furthermore, downregulation of the ARMS gene limited the migration of embryoid body cells derived from mESCs. This study indicates novel roles of Kidins220/ARMS in ESCs, which may represent valuable targets for future clinical applications of ESCs.
Introduction
M
However, the fact that very few cell surface markers in mESCs have been identified limits the feasibility of isolating and studying mESCs. Therefore, it is crucial to expand the repertoire of ESC markers and to understand the underlying molecular mechanisms that modulate their proliferation, pluripotency, and differentiation.
The kinase D interacting substrate of 220 kDa (Kidins220), also known as ankyrin repeat-rich membrane spanning (ARMS) protein, has been found to play a substantial role in neuronal survival and differentiation, activation of T and B cells, and proliferation and apoptosis of tumor cells.
The nonsense variants in Kidins220/ARMS present at precise moments in nervous system development, eventually causing spastic paraplegia, intellectual disability, nystagmus, and obesity (SINO) (Josifova et al., 2016). The expression of Kidins220/ARMS declines with the differentiation of neural cells both in vitro and in vivo (Cortes et al., 2007), suggesting that it exerts an influence on nervous system development. Meta-analysis of Kidins220/ARMS expression of peripheral neuroblastic tumors in different differentiation states also demonstrated that the expression of Kidins220/ARMS was downregulated as the tumors differentiated (Rogers and Schor, 2013).
These results indicated that Kidins220/ARMS was associated with embryonic development, tumor progression, and other biological processes. In the previous study, we carried out a large-scale proteome analysis to identify the specific surface markers of mESCs. Nearly 1000 secreted and transmembrane cell surface proteins were identified, including a large number of functional proteins associated with signal transduction, adhesion, and transporting. As an essential scaffold protein involved in nervous and cardiovascular system development, as well as spatial and temporal regulation during development, Kidins220/ARMS was detectable and overexpressed on the surface of mESCs (Gu et al., 2010).
However, whether Kidins220/ARMS was involved in the self-renewal, proliferation, or differentiation of mESCs was still unclear. To elucidate these possible effects of Kidins220/ARMS on mESCs, in this study we investigate the functions of Kidins220/ARMS in mESCs.
Materials and Methods
Cell culture
Mouse embryonic fibroblast (MEF) were obtained from mouse embryos 13.5 days postfertilization and cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS; Gibco). mESCs of line D3, from ATCC (www.atcc.org/), were cultured on mitomycin-c (Sigma-Aldrich, St. Louis, MO) treated MEFs in mESC culture medium (85% DMEM [Gibco], 15% FBS [Gibco], 0.1 mM 2-mercaptoethanol [Sigma], 1000 U/mL mouse leukemia inhibitory factor [LIF] [Chemicon, Temecula, CA]) at 37°C in 5% CO2.
Immunocytochemistry
The mESCs were fixed in 4% paraformaldehyde (Sigma-Aldrich), and permeabilized with 0.1% Triton 100. Then, samples were blocked with phosphate-buffered saline (PBS) containing 5% goat serum for 30 minutes. Rabbit polyclonal antibody against Kidins220/ARMS (Abcam, Cambridge, United Kingdom) was diluted in PBS containing 5% goat serum followed by incubation for 1 hour. After washing twice, the second antibody was diluted and incubated for 1 hour at 37°C. The labeled cells were stained with Hoechst 33258 (Merck, Darmstadt, Germany) and examined by fluorescence microscopy (Leica, Wetzlar, Germany).
Construction of plasmids and lentivirus production
DNA sequences coding for shRNA were acquired from the RNAi Consortium (www.broadinstitute.org). pLKO TRC-cloning vector (Addgene plasmid 10878) was obtained from Addgene. The vector sequence of the puromycin selection marker was replaced with enhanced green fluorescence protein (EGFP). After that, the shRNA-coding DNA were cloned into the new vector. To produce the lentivirus, PsPAX (Addgene plasmid 12260), pMD2.G (Addgene plasmid 12259), and the modified pLKO plasmids were co-transfected into the 293T cells. The mESCs were seeded into 96-well plates at a density of 3000 cells per well for virus infection and incubated for 24 hours with packaged lentivirus. Monoclones with strong green fluorescence protein (GFP) fluorescent intensity were picked out. The pLKO-shscramble-pools (Addgene plasmid 1864) virus was used as a control.
Acquisition of interfering shRNA-ARMS pools and monoclone H10, F6
To research the role of Kidins220/ARMS in mESCs, the interfering shRNA plasmids of Kidins220/ARMS were constructed. After transfection, EGFP was expressed in the shRNA-ARMS pool (Supplementary Fig. S1A; Supplementary Data are available online at www.liebertpub.com/cell). At an average density of 0.5 cells per well, we transferred the diluting shRNA-ARMS pool into each well of a 96-well plate. After undisturbed incubation for 7–14 days, monoclones were selected. The quantitative real-time polymerase chain reaction (qRT-PCR) confirmed that the expression of Kidins220/ARMS in monoclones H10 and F6 were significantly more inhibited than in other clones (monoclone H10, p < 0.01, F6, p < 0.05) (Supplementary Fig. S1B), so both of them were selected for further research.
qRT-PCR analysis
Total RNA was extracted from the mESCs pools or monoclones with TRIzol reagent (Invitrogen). Reverse transcription was performed using Moloney murine leukemia virus (Promega, Fitchburg, WI) for 1 hour at 42°C. Then qRT-PCR was used to measure Kidins220/ARMS gene expression relative to β-actin gene expression with the primers, as follows. Forward: ACGCCACTGATGATAGCTGC, Reverse: TGCCCCTCTTTAGATGCTGAT.
MTT assay
Mouse embryonic stem cells (MESCs) with MEF were dissociated with 0.25% trypsin digestion and added with fresh DMEM culture medium, followed by differential attachment in T25 bottles incubated with 0.5% gelatin. After 1.5 hours incubation, unattached MESCs were harvested and seeded in 96-well plates with 5000 cells per well, and cultured for 1–5 days. The cell viability after time-dependent treatments was determined using MTT assay. MTT [(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)] (5 mg/mL) were added to each well and the plates were incubated for 3 hours at 37°C. Then, the purple formazan crystals were dissolved in DMSO, and the absorbance was recorded with a microplate reader (Bio-Rad) at a wavelength of 590 nm.
Alkaline phosphatase and colony-forming assay
Alkaline phosphatase (ALP) staining was performed using the Leukocyte Alkaline Phosphatase Kit (Sigma-Aldrich). Images were acquired using an inverted microscope (Olympus BX-70) and processed by the ImageJ software. For the colony-forming assay, about 300 mESCs were transferred to each well containing MEFs in a 96-well plate. At day 6, colonies were stained for ALP. The ALP-positive colonies and total colonies were counted and the colony-forming rate was calculated.
Embryoid body formation
After clones of D3 cells were digested into single cell with 0.25% trypsin plus 0.2% EDTA (Thermo Fisher Scientific, MA), 20 μL/drop cell suspension of mESCs were pipetted onto the inverted lid of a 10 cm Petri plate. Then, the inverted lid was placed on the culture plate, which was full of sterilized PBS. After 2 days, embryoid bodies (EBs) formed in the hanging drops were collected together from the lids and were cultured in differentiation media (LIF free).
Statistical analysis
All statistical analyses were carried out using SPSS 19.0 (SPSS, Chicago, IL). Statistical comparisons were performed with one-way ANOVA tests and two-way ANOVA tests. A p-value of <0.05 was considered statistically significant. Data were presented as mean ± standard error of the mean of at least three independent experiments.
Results
Kidins220/ARMS was expressed on the surface of mESCs
In our previous study, we found, using mass spectrometric analysis, that Kidins220/ARMS was expressed on the surface of mESCs (Gu et al., 2010). Based on this information, we further verified the expression level of Kidins220/ARMS on the surface of mESCs. The results from immunohistochemistry (IHC) indicated that Kidins220/ARMS was expressed on the surface of mESCs (Fig. 1A). The expression of Kidins220/ARMS can also be detected by qRT-PCR analysis; however, the expression of Kidins220/ARMS is relatively lower compared with that of Oct4 and Nanog in mESCs (Fig. 1B).
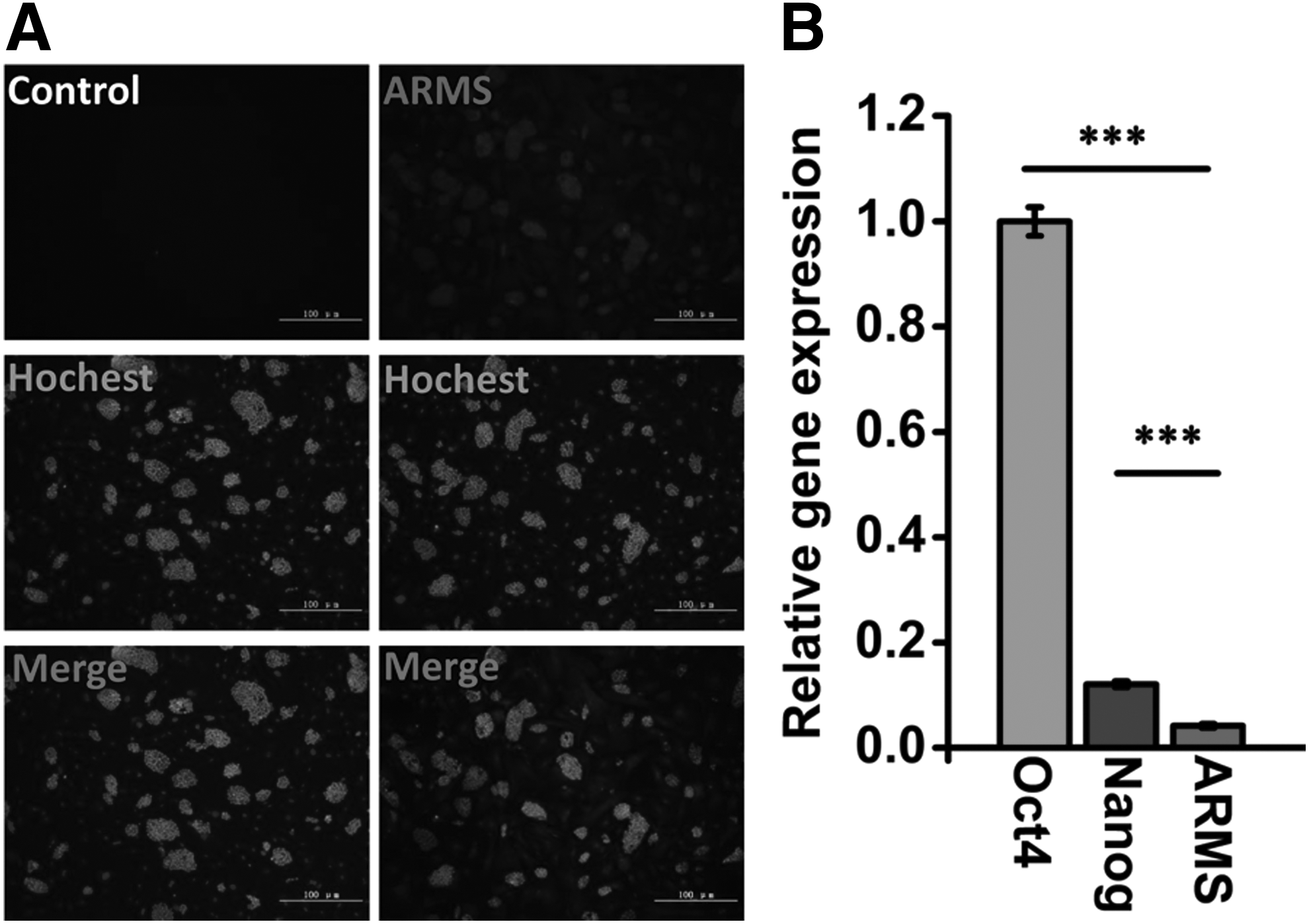
Kidins220/ARMS gene expression was evaluated in mESCs.
The proliferation and survival of mESCs were markedly inhibited due to the downregulation of Kidins220/ARMS
The mESCs, shscramble-pools, and shARMS-pools were cultured on mouse embryonic feeder. Contrary to the D3 cell line and mock cell line, no obvious clones were observed in shARMS-pools during the first 2 days. As the cells proliferated, smaller clones appeared in shARMS-pools after 3 days (arrows) compared with D3 and mock (Fig. 2A). The results of cell counts also indicated that cell proliferation of shARMS-pools, shARMS-H10, and shARMS-F6 were significantly reduced compared with that of D3 and mock after 4 days culture (Fig. 2B). Passage time of shARMS-pools was nearly 8 days, which was longer than D3 and mock. This indicated that Kidins220/ARMS was involved in the regulation of cell growth in mESCs, an inference that was further confirmed by MTT assays (Fig. 2C).
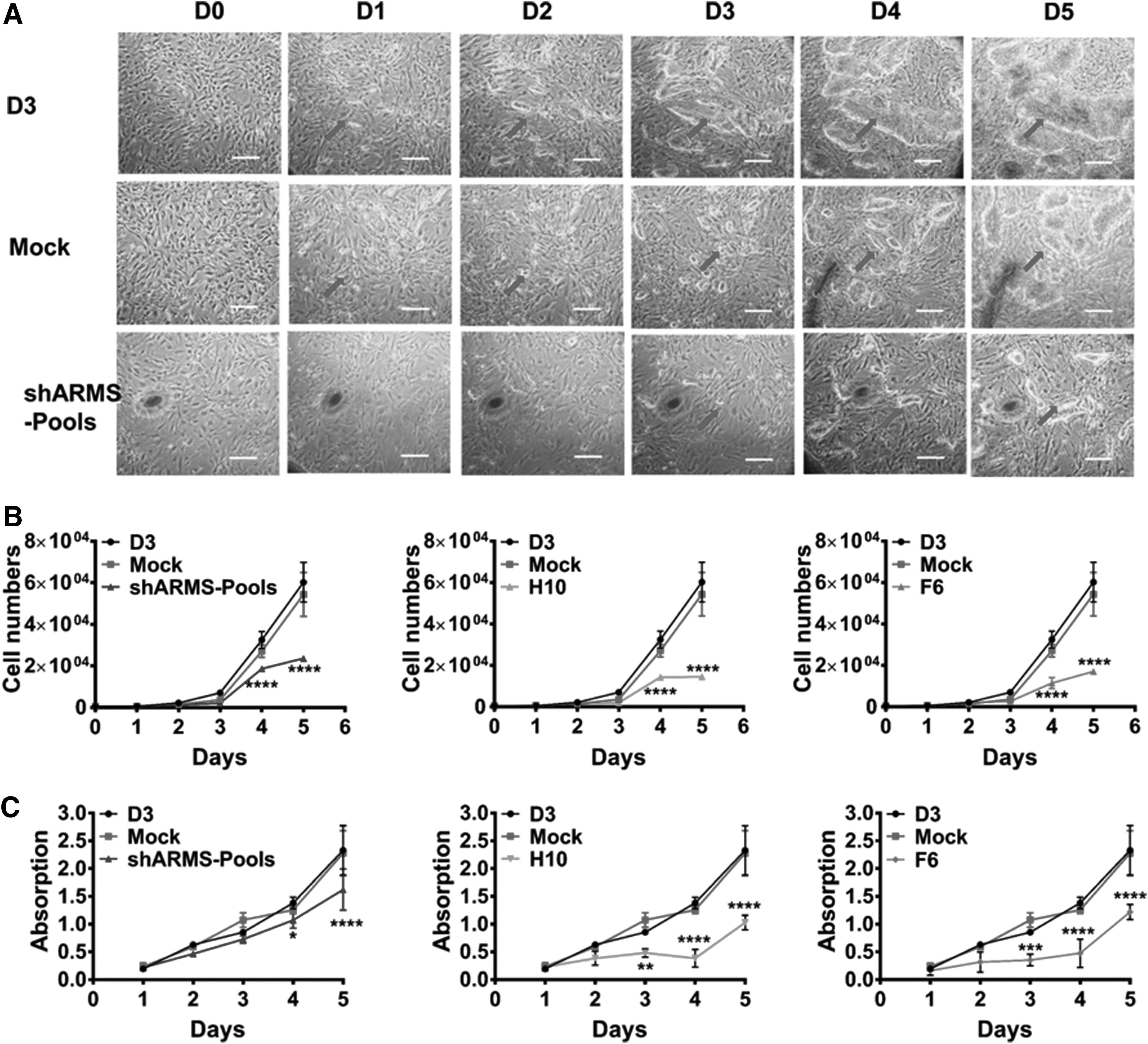
Cell proliferation was evaluated among the cell lines: wild D3, mock, shARMS-pools, monoclone shARMS-F6, and shARMS-H10.
Kidins220/ARMS was not involved in self-renewal of mESCs
Since Kidins220/ARMS is associated with the embryo development (Cesca et al., 2011), we investigated whether Kidins220/ARMS was involved in self-renewal maintenance in mESCs. By use of ALP staining assay, no obvious differences were observed between D3, mock, and shARMS-pools cell lines. Statistical analysis also indicated that there were no significant differences among these pools (Fig. 3A). This suggested that Kidins220/ARMS was not related to the self-renewal of mESCs.
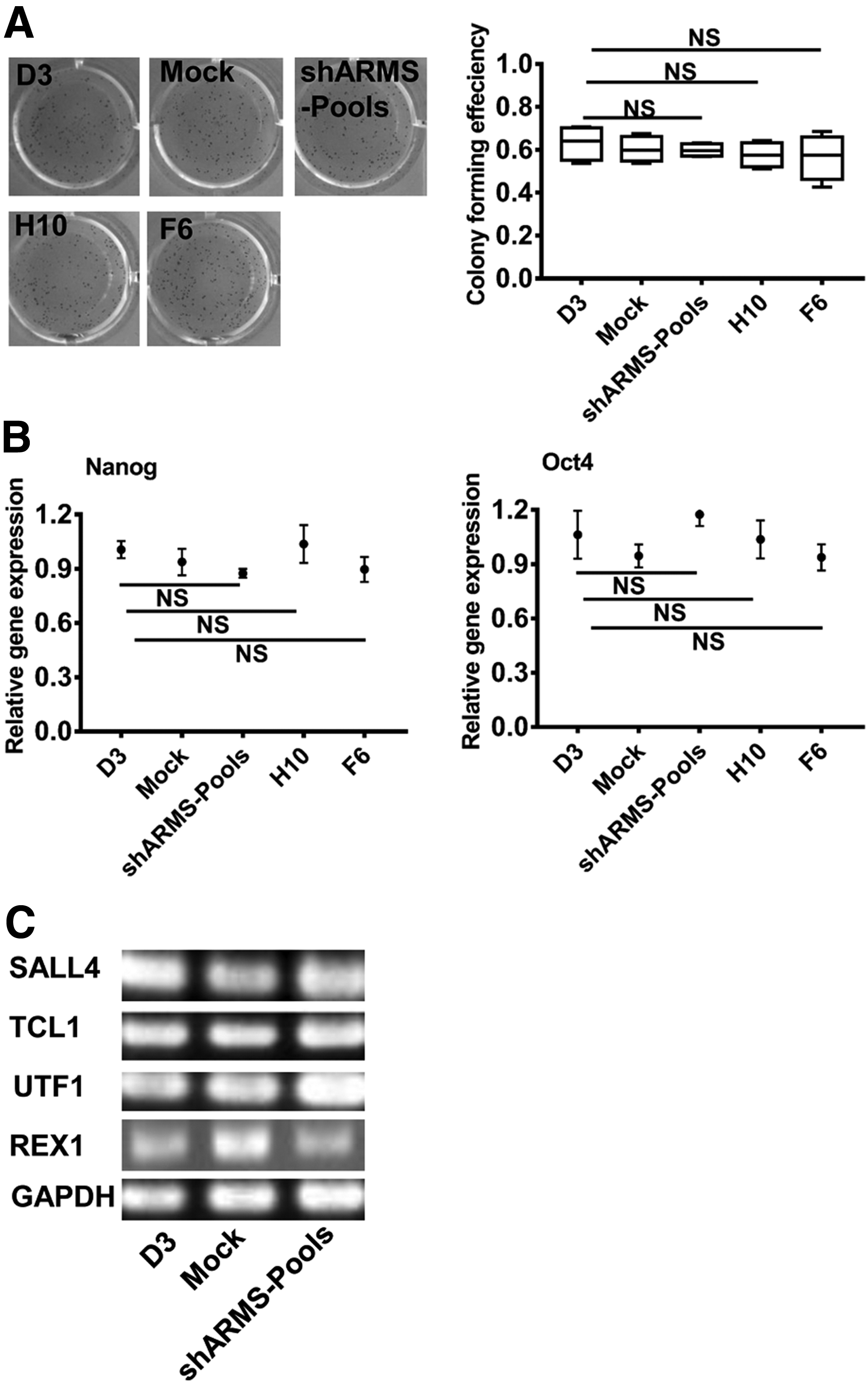
The downregulation of Kidins220/ARMS was not involved in self-renewal of mESCs.
To further investigate the relationship between Kidins220/ARMS and self-renewal of mESCs, we quantified the gene expression related to mESCs self-renewal. No significant expression changes in Nanog or Oct4 were detected among the three pools (Fig. 3B). We also performed further investigations associated with a variety of other markers, such as UTF1 (undifferentiated embryonic cell transcription factor 1), SALL4 (spalt-like transcription factor 4), TCL1 (T cell leukemia 1), and REX1 (reduced expression protein 1). In accordance with Nanog and OCT4, these genes showed similar expression levels among three pools (Fig. 3C).
Kidins220/ARMS influences the EB formation and cell migration
EBs obtained through the hanging-drop culture method were cultured in LIF-free media. Compared with EBs derived from D3 and mock cell, the EBs derived from shARMS-pools formed multicellular aggregates without any cell migration during adherent culture (Fig. 4). This suggested that Kidins220/ARMS played an important role in cell migration.
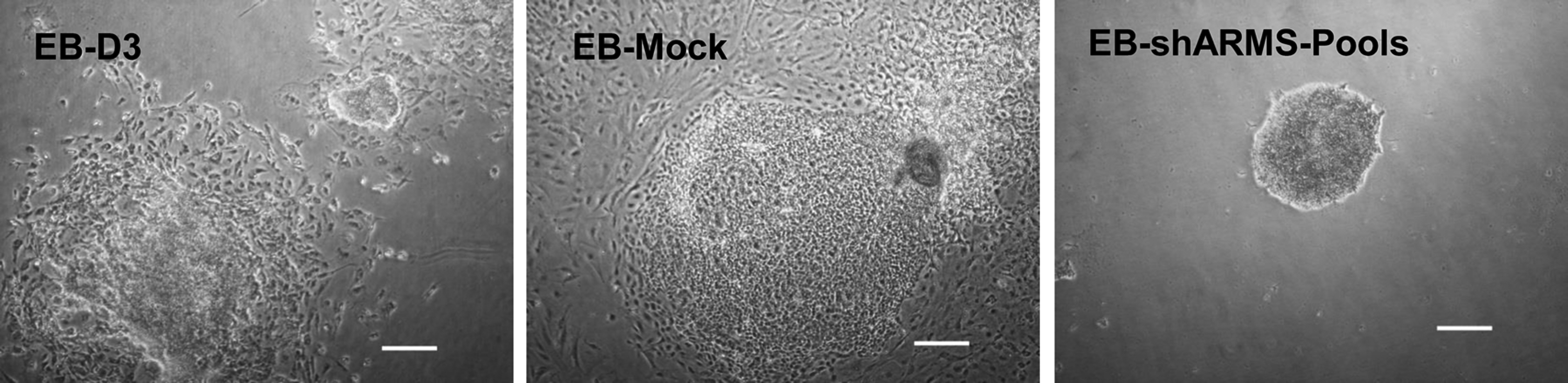
Cell migration was inhibited in EBs derived from shARMS-pools, relative to EB-D3 and EB-mock. Scale bars = 100 μm. EB, embryoid body.
Discussion
Kidins220/ARMS is a type of transmembrane protein that is abundant in developing neural tissues (Lopez-Menendez et al., 2013).
Previous studies concerning Kidins220 mainly focused on its effects on neuronal differentiation, survival, and function (Gamir-Morralla et al., 2015; Lopez-Menendez et al., 2013). Various studies have demonstrated that heterozygous Kidins220/ARMS nonsense variants had disastrous consequences, such as spastic paraplegia, nystagmus, intellectual disability, and obesity (Josifova et al., 2016; Mero et al., 2017). Other groups' studies have indicated that Kidins220/ARMS mediated the integration of neurotrophin and vascular endothelial growth factor (VEGF) signaling during neuronal and cardiovascular development. Kidins220/ARMS (−/−) embryos died at late stages of gestation (Cesca et al., 2011, 2012).
In this study, we demonstrated that the Kidins220/ARMS gene was highly expressed on the surface of mESCs. This result is consistent with a previous proteome study that demonstrated that Kidins220/ARMS was also enriched in mESCs (Gu et al., 2010). Quantitative PCR analysis demonstrated that the knockdown of Kidins220/ARMS did not influence the expression of OCT4 and Nanog, which suggested that the changes in Kidins220/ARMS might not contribute to the self-renewal maintenance of mESCs.
However, the downregulation of Kidins220/ARMS was associated with reduced proliferation of prostate cells and significantly inhibited the tumor growth (Wang et al., 2016). In neuroblastoma cells, Kidins220/ARMS is also a key regulator involved in cell proliferation (Jung et al., 2014). One study found that Kidins220/ARMS was crucial to neuronal survival and functioning, and its downregulation resulted in the neuronal death (Gamir-Morralla et al., 2015). In our study, the downregulation of Kidins220/ARMS also significantly inhibited the proliferation of mESCs and was associated with reduced clone size, indicating that Kidins220/ARMS had a notable impact on the proliferation and survival of mESCs.
These facts suggest that the cytoprotection conferred by Kidins220/ARMS is broad spectrum and deserves further study as a potential target for application to clinical repair of injured tissues or organs. Consistent with previous research, we found that Kidins220/ARMS played an important role in embryonic development. During EB formation, the knockdown of Kidins220/ARMS limited the outward migration of EB cells. This indicated that Kidins220/ARMS correlates with embryonic development and individual maturity.
In summary, we have confirmed that Kidins220/ARMS was expressed on the surface of mESCs and was associated with their proliferation, survival, and regulation during development. However, the gene was not involved in the self-renewal and maintenance of mESCs. The underlying molecular mechanism associated with Kidins220/ARMS during development should be further investigated for its potential role in the clinical application of ESCs.
Footnotes
Acknowledgments
This work was supported by a grant of medical scientific research foundation of Zhejiang Province of China (grant no. 2017KY394) and the National Basic Research Program of China (grant no. 2007CB947800).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.