
Abstract
Abstract
PRDM14, a member of the PRDM family protein, plays an important role in the regulation of epigenetic reprogramming. Knockdown of Prdm14 in germ cells can lead to female and male subfertility, and the function of PRDM14 appears to be conserved across mammalian species. Thus, we analyzed the expression of Prdm14 in parthenogenetic embryos at all stages of preimplantation and then evaluated the effect of Prdm14 downregulation on porcine parthenogenetic embryonic development. We found that Prdm14 transcripts are highly expressed in the metaphase II (MII) oocyte, and their level gradually increased from the 2-cell to 8-cell stage and slightly declined at the blastocyst stage during the development of parthenogenetic porcine embryos. Prdm14 was knocked down in oocytes at the MII stage by the injection of siRNA to assess the role of this protein in epigenetic remodeling and embryo development. Prdm14 knockdown significantly decreased the cleavage and blastocyst rates without changing the total cell number of the blastocysts. In addition, the expression levels of the antiapoptotic gene BCL-2 and the pluripotency-related genes OCT4 and SOX2 were also significantly decreased. These results showed that PRDM14 could affect embryonic development through regulating the expression levels of the pluripotency and antiapoptotic genes during the development of parthenogenetically activated porcine embryos.
Introduction
N
DNA methylation is a crucial epigenetic modification that regulates the imprinting (differential silencing of maternal or paternal alleles) and repression of retrotransposons and other parasitic DNA and might also control X-chromosome inactivation and cellular differentiation (Kang et al., 2013). During DNA methylation, methylcytosine is formed by the addition of a methyl group to the 5′ position of cytosines, and this reaction is catalyzed by DNA methyltransferases (Dnmts) (Messerschmidt et al., 2014). In addition to methylation, active demethylation is mediated by enzymes of the TET (ten–eleven translocation methylcytosine dioxygenase) family and, specifically, these enzymes oxidize methylcytosine to hydroxymethylcytosine, which is further converted to unmethylated cytosine (Tahiliani et al., 2009).
PRDM14 (PR domain [PRDI-BF1 and RIZ homologous region] containing 14), a member of the PRDM family of proteins, bears a single PR domain with six tandemly repeated zinc fingers (Nakaki and Saitou, 2014). There is evidence showing that PRDM14 plays an important role in epigenetic modification, such as histone modification and DNA methylation. This protein can recruit TET proteins (Wu and Zhang, 2014) and repress the de novo Dnmt3 through the recruitment of polycomb repressive complex 2 to their promoters (Boyer et al., 2006); therefore, PRDM14 activates the expression of these genes through promoter demethylation (Okashita et al., 2014). However, the precise mechanisms underlying this phenomenon remain unknown and require further investigation.
Currently, there has been little study of PRDM14 among species other than mice and humans. Thus, we explored the expression of Prdm14 in embryos at all stages of preimplantation and assessed the effect of PRDM14 on parthenogenetic porcine embryos. Our data suggest that PRDM14 could affect embryonic development through regulating the expression levels of the pluripotency genes and antiapoptotic genes during the development of parthenogenetically activated porcine embryos.
Materials and Methods
Oocyte collection, in vitro maturation, and PA
Porcine ovaries were collected from a local abattoir and transported to the laboratory at 38.5°C in 2–4 hours. Then, a 10-mL syringe attached to an 18-gauge needle was used to aspirate the cumulus-oocyte complexes from the follicle and washed three times in manipulation fluid (TCM-199 supplemented with 0.1% polyvinyl alcohol). Cumulus cells with a relatively uniform cytoplasmic density and at least three layers of cells were selected and cultured at 38.5°C in 500 μL of maturation medium (Liu et al., 2012) (TCM-199 supplemented with 10 ng/mL epidermal growth factor, 0.5 μg/mL porcine luteinizing hormone, 0.5 μg/mL porcine follicle-stimulating hormone, 26 mM sodium bicarbonate, 3.05 mM glucose, 0.91 mM sodium pyruvate, 0.57 mM cysteine, 0.1% PVA, 10% fetal calf serum, 75 mg/mL penicillin G, and 50 mg/mL streptomycin) in a humidified 5% CO2.
After 42–44 hours, cumulus cells were removed by exposure to 0.2% hyaluronidase for 3 minutes. Oocytes with the first polar body and an intact cytoplasm were subjected to PA using two DC pulses of 1.2 kV/cm for 30 microseconds in fusion medium. After this stimulation, the oocytes were cultured in PZM-3 medium (NaCl 108.00 mmol/L, KCl 10.00 mmol/L, KH2PO4 0.35 mmol/L, MgSO4·7H2O 0.40 mmol/L, NaHCO3 25.07 mmol/L, Na-pyruvate 0.20 mmol/L, Ca− (lactate) ·5H2O 2.00 mmol/L,
siRNA microinjection
An RNA interference-based strategy similar to that described by Okada et al. (2010) was used. After 42–44 hours, oocytes were digested in 0.1% hyaluronidase for 2 minutes at 37°C. When most of the cumulus cells were separated from the oocytes, oocytes with an intact cytoplasm and the first polar body were selected and transferred into TCM199-Hepes medium and four candidate siRNAs (si-PRDM14-1, si-PRDM14-2, si-PRDM14-3, and nonsense RNA) were chemically synthesized to target porcine Prdm14 and was microinjected into MII-stage oocytes using the Eppendorf CellTram vario system (Eppendorf, Hamburg, Germany). Then, oocytes for knockdown were cultured in PZM-3 medium, selected by detection of FAM fluorescence and then cultured until use, as previously described (Matsuoka et al., 2008; Tsunemoto et al., 2008).
To determine the knockdown efficiency, mRNA isolated from Prdm14 knockdown in 50 4-cell embryos was subsequently subjected to qRT-PCR. The cleavage rates, blastocyst rates and total cell number per blastocyst were calculated at 2 and 6 days after siRNA injection, respectively.
RNA isolation, cDNA synthesis, and qRT-PCR
Total RNA extracts from ∼200 porcine oocytes or parthenogenetic embryos blastomere at each stage were purified with the RNeasy Mini Kit (Qiagen, Hilden, Germany) (Zhang et al., 2017). According to the manufacturer's recommended protocol, cDNA synthesis was performed using the First-Strand cDNA Synthesis Kit (Promega, Madison, WI). The primers used for PRDM14 are listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/cell).
The real-time PCR (qRT-PCR) mix contained 10 μL of SYBR green premix, 0.5 μL of each forward and reverse primer (0.5 μM), 2 μL of cDNA (1000 ng), and 7 μL of RNase-free water. The samples were denatured at 95°C for 30 seconds and then subjected to 40 cycles of amplification (95°C for 5 seconds and 60°C for 30 seconds), one cycle for melting curve analysis (95°C for 5 seconds, 60°C for 1 minute, and 95°C for 1 second) and a cooling step at 4°C. The data were performed three times for each sample replace. In each sample, the expression level was calculated relative to the expression level of the pig GAPDH mRNA. We used the 2−ΔΔCT method to calculate the relative transcription levels for each gene (Hou et al., 2016).
Immunofluorescence staining
We removed the zona pellucida by 0.5% pronase in PZM-3 at 37°C for 3 minutes. The zona pellucida-free oocytes and embryos were washed three times in PBS-PVA (PBS contains 0.1% polyvinyl alcohol), fixed with 4% paraformaldehyde for 30 minutes at room temperature, permeabilized for 20 minutes with 0.2% Triton X-100 prepared in PBS-PVA, and then the samples were separated into two groups. One group of embryos was subjected to staining for PRDM14. To prevent the binding of nonspecific sites, the samples were incubated in 2% BSA for 1 hour at 37°C, and then incubated with the primary antibody at 37°C for 1.5 hours. Rabbit anti-PRDM14 antibody was diluted in PBS for 1:200. The samples were then washed with PBS-PVA and incubated for 1 hour with red-labeled secondary antibodies for PRDM14 diluted 1:200 in PBS containing 1% BSA.
The second group of samples was subjected to TUNEL staining. After permeabilization, the samples were then stained with TUNEL according to the manufacturer's specifications (Su et al., 2011). DNA was stained with 10 μg/mL Hoechst 33342 for 10–15 minutes. All samples were immediately mounted and observed under a fluorescence microscope. Images were analyzed using ImageJ software.
Statistical analysis
All experiments were replicated at least three times. The data were analyzed by one-way ANOVA using SPSS 19.0 software (SPSS, Inc., Chicago, IL). A value of p < 0.05 was considered statistically significant, and p < 0.01 was considered extremely significant.
Results
Expression of PRDM14 in parthenogenetically activated porcine embryos
We used the fluorescent quantitative PCR technique to explore the dynamic changes in Prdm14 expression during the early embryonic period. Prdm14 exhibited the highest transcription level in porcine MII oocytes (Fig. 1A). The results also showed that during the development of parthenogenetic porcine embryos, the Prdm14 expression level decreased from the oocyte stage to the 2-cell embryo stage, gradually increased until 8-cell embryo stage, and then decreased until the blastocyst stage.
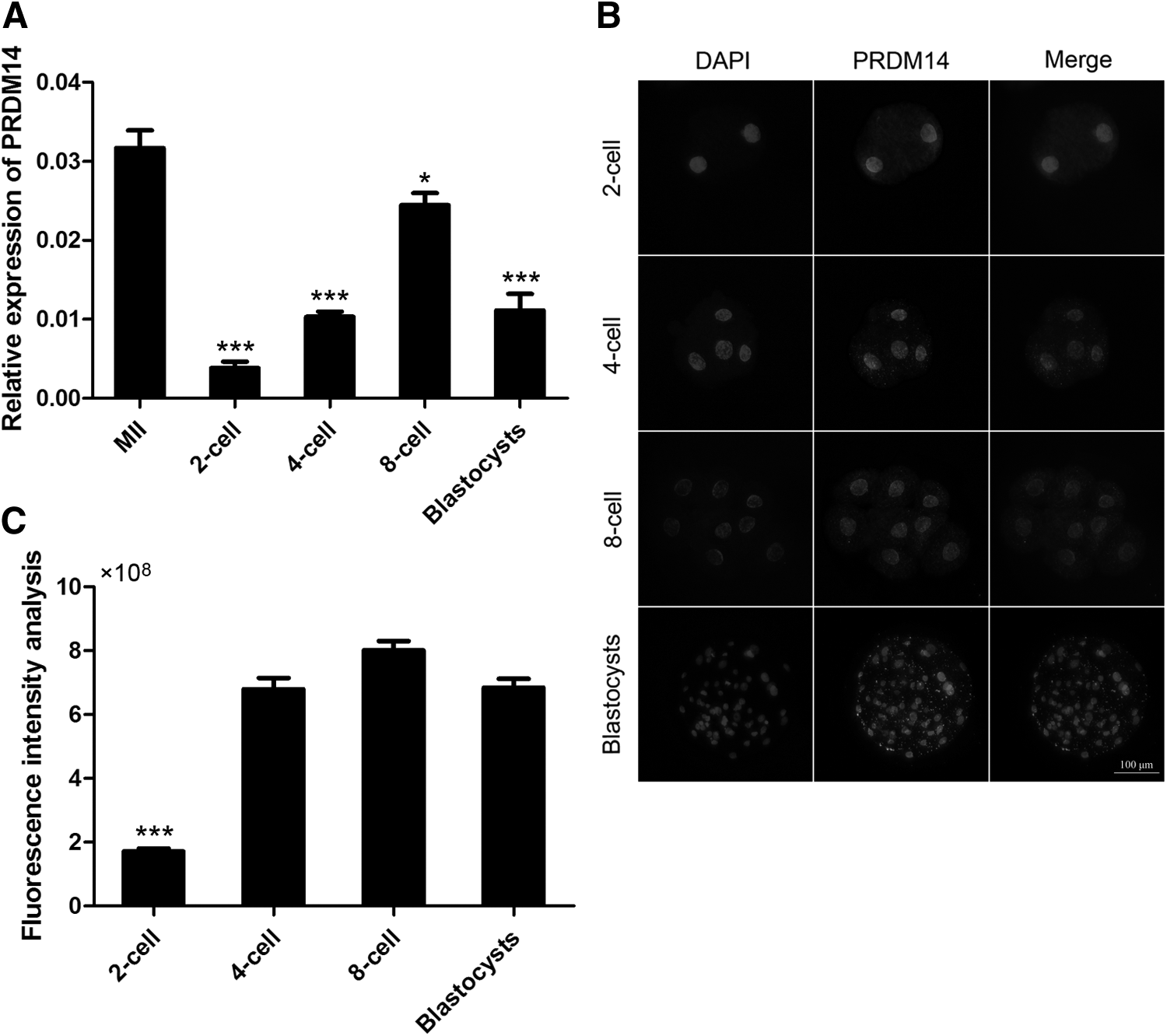
Relative abundance of PRDM14 in parthenogenetic preimplantation porcine embryos.
We subsequently examined the PRDM14 protein expression profile in parthenogenetically activated porcine embryos through an immunofluorescent (IF) staining analysis (Fig. 1B, C). The IF staining results indicated that PRDM14 protein was present in all parthenogenetically activated porcine embryos, and that the predominant localization of this protein was the nucleus. Consistent with the mRNA levels, the signal gradually increased during the development of parthenogenetic porcine embryos and then decreased at the blastocyst stage.
Downregulation of PRDM14 mRNA levels in porcine MII oocytes by siRNA injection
An RNAi approach was implemented to decrease the PRDM14 mRNA expression level in porcine MII oocytes to study the function of this transcript during parthenogenetic embryo development. A fluorescein-labeled siRNA was used to optimize the efficiency of the injection of porcine oocytes (Fig. 2A). Fluorescence could be detected under a fluorescence microscope after the injection of all tested siRNAs [si-PRDM14-1, si-PRDM14-2 si-PRDM14-3, and N.C. (control-siRNA)], and the levels of Prdm14 in 4-cell embryos were analyzed by real-time qPCR (Fig. 2B). Control siRNA-injected and noninjected embryos were also analyzed. As shown by an analysis of the mRNA level, si-PRDM14-1 had higher interference efficiency than the other.
Based on these results, si-PRDM14-1 was selected as the optimal siRNA and used in the following experiments. We also analyzed the expression of Pdm14 at the blastocyst, and the results showed that there was no difference in Prdm14 expression at the blastocyst (p < 0.05).
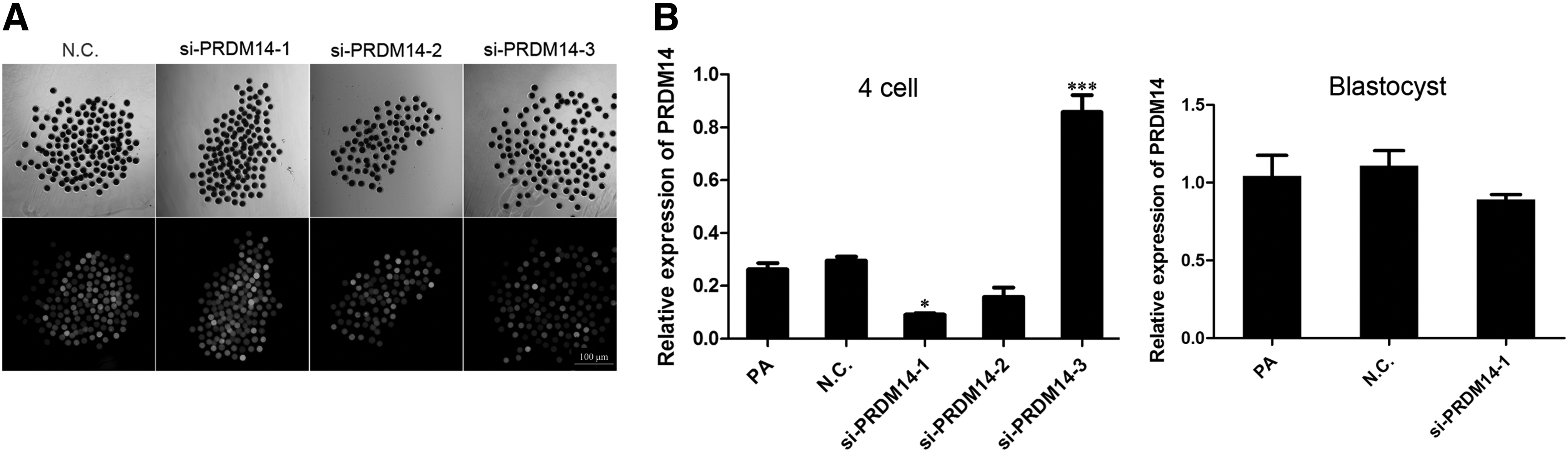
Efficient knockdown of Prdm14 mRNA levels in 4-cell embryos by siRNA injection into MII oocytes.
Effect of PRDM14 knockdown on embryo development and quality
The developmental potential of PRDM14 siRNA-injected oocytes was evaluated by PA of MII oocytes and their subsequent culture to the blastocyst stage. As shown in Table 1, the rate of embryo cleavage in the PRDM14-siRNA group was significantly lower than that of the control groups: 80.51% ± 0.68%, 79.59% ± 1.19%, and 61.20% ± 1.44% of the oocytes in the noninjected (p < 0.05), control siRNA-injected, and PRDM14-siRNA groups, respectively, showed cleavage after PA. The proportion of activated oocytes that developed to the blastocyst stage was significantly lower in the PRDM14-siRNA group (9.97% ± 0.57%) compared with the noninjected and control siRNA-injected groups (25.43% ± 0.29% and 21.37% ± 0.35%, respectively) (p < 0.05). In addition, the proportion of cleaved oocytes in the PRDM14-knockdown group that reached the blastocyst stage was lower than that in the control group.
Effects of Interfering PRDM14 Expression on the Development of Embryos
Data are the mean ± SEM of at least three trials. Values in the same column with different superscripts differ significantly (p < 0.05).
N.C., control-siRNA; PA, parthenogenetic activation; SEM, standard error of the mean.
Expression of pluripotency-related genes in PRDM14-knockdown embryos
The effect of PRDM14 knockdown on the expression levels of the pluripotency genes OCT4 and SOX2 at the 4-cell and blastocyst stages during the development of parthenogenetic porcine embryos was evaluated by qRT-PCR (Fig. 3). The mRNA expression levels of OCT4 and SOX2 at the 4-cell stage were significantly lower in the PRDM14-siRNA group than in the noninjected and control siRNA-injected groups (p < 0.05). However, no significant differences in the OCT4 and SOX2 mRNA levels at the blastocyst stage were found among the three groups (p > 0.05).
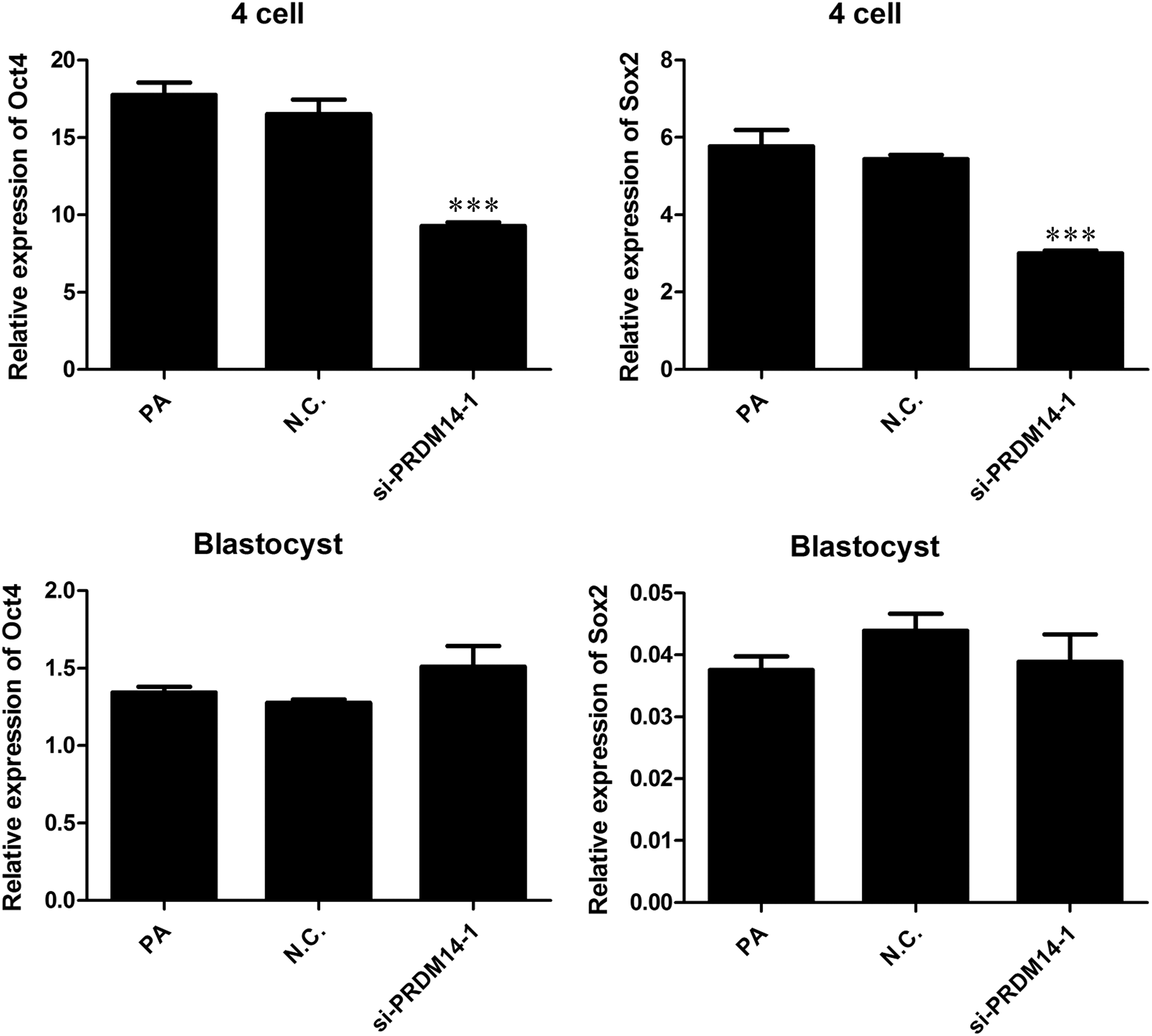
The relative abundance of OCT4 and SOX2 mRNA at the 4-cell and blastocyst stages among the three groups. PRDM14 knockdown reduced the relative abundances of OCT4 and SOX2 mRNA at the 4-cell stage but did not influence the expression levels of OCT4 and SOX2 at the blastocyst stage. The results shown are the means ± standard deviations from three independent experiments; ***p < 0.001, compared with the noninjected control group. N.C., control-siRNA; PA, noninjected control; si-PRDM14-1, PRDM14-siRNA.
Effect of PRDM14 knockdown on cell apoptosis in parthenogenetic preimplantation porcine embryos
To assess the quality of the embryos that developed into blastocysts, we determined the total cell number per blastocyst and the number of apoptotic cells in the blastocysts in each experimental group. The results indicated no differences in the total cell number and the number of apoptotic cells among the three groups (Fig. 4A).
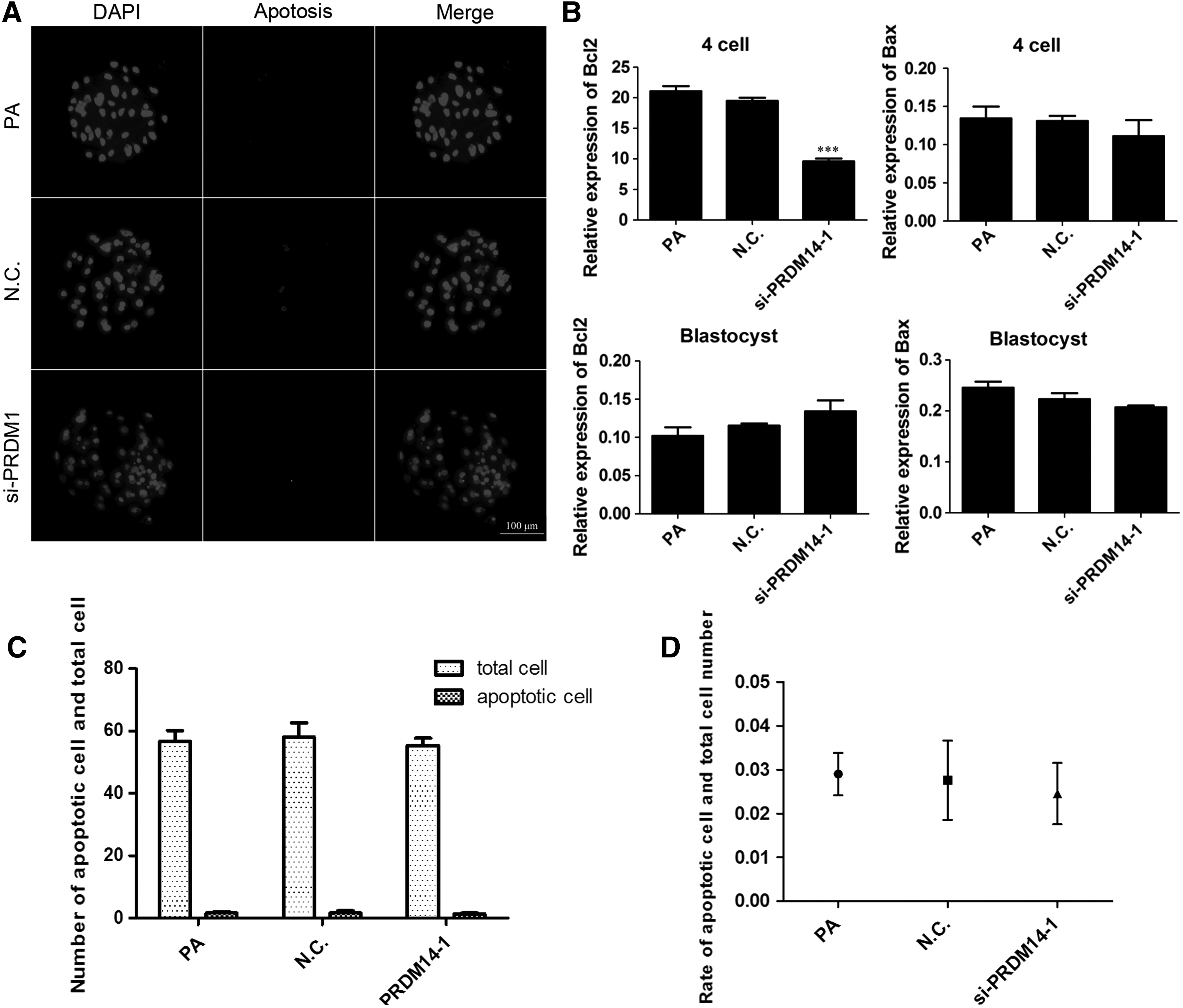
Effect of PRDM14 knockdown in porcine oocytes on apoptosis. TUNEL assay of blastocyst stage embryos of the three groups. The fragmented nuclei are apoptotic, and each sample was counterstained with Hoechst 33342 to visualize DNA.
For the various groups, the mRNA levels of the apoptosis-related genes BAX and BCL-2 in embryos at the 4-cell and blastocyst stages were examined using qRT-PCR (Fig. 4B). The results showed that the mRNA expression levels of BCL-2 at the 4-cell stage were significantly lower in the PRDM14-siRNA group than in the noninjected and control siRNA-injected groups (p < 0.05). However, there was no significant difference in the BCL-2 mRNA levels at the blastocyst stage among the three groups. The expression of BAX has no change in the embryos at all preimplantation stages (p > 0.05) compared with that of the noninjected and control siRNA-injected groups.
Discussion
PRDM14 belongs to the family of PR domain-containing (PRDM) transcriptional regulators (Volkel and Angrand, 2007), and among the members of this family, PRDM14 shows specific expression in preimplantation embryos, primordial germ cells (PGCs), and embryonic stem cells in vitro, and accordingly plays a pivotal role in the regulation of their pluripotency and epigenetic reprogramming (Nakaki and Saitou, 2014).
However, the role of PRDM14 in parthenogenetically activated porcine embryos has not been fully elucidated. Here, the changes in the expression levels of apoptosis and pluripotency-related genes during the development of preimplantation embryos subjected to RNAi were assessed and compared with the normal group, and the findings will provide insights for further study of the mechanisms underlying the effect of PRDM14 on parthenogenetically activated porcine embryos.
Previous research (Burton et al., 2013) found that Prdm14 gene expression is highest in 2-cell embryos, decreases after the 8-cell stage, and recovers in blastocysts (Kurimoto et al., 2006; Yamaji et al., 2008). In humans, Prdm14 is detected starting at the 8-cell stage and shows increased expression in blastocysts (Xue et al., 2013; Yan et al., 2013). In this study, we found that Prdm14 mRNA had the highest transcription levels during oogenesis and decreased after PA, which differs from what is observed in mice and humans. This difference might be due to the storage of factors in MII oocytes, and to the expression of the majority of genes during this period; thus, the process requires the adjustment and control of transcription factors.
After PA, the level decreased at the 2-cell stage, gradually increased until the 8-cell stage, and then decreased at the blastocyst stage. Similar findings have also been observed at the protein level and the protein was located mainly in the nucleus, suggesting that Prdm14 is involved in transcriptional activation and might play a role during embryogenesis. In mice, after implantation, Prdm14 is no longer expressed in epithelial cells and is again expressed in PGCs (Yabuta et al., 2006; Yamaji et al., 2008). The expression of Prdm14 in PGCs continues to about E13.5, and after that, the male PGCs undergo mitotic arrest for development as prospermatogonia, whereas the female PGCs begins meiosis as oogonia (Nakaki and Saitou, 2014). Therefore, cells expressing PRDM14 acquire the potential for pluripotency and the ability to undergo epigenetic reprogramming.
PRDM14 is thought to be involved in the epigenetic control of gene expression in undifferentiated ESCs and PGCs (Tsuneyoshi et al., 2008). To better understand the role of PRDM14 in porcine zygote and embryo development, we assessed the effect of Prdm14 knockdown in oocytes on developmental competence. The injection of PRDM14 siRNA into MII oocytes resulted in significantly lower cleavage and blastocyst rates, suggesting that PRDM14 could affect embryonic development during the development of parthenogenetically activated porcine embryos. A similar approach has been used to study the role of other genes in bovine oocytes and early embryos (Lee et al., 2009).
In mice, PRDM14 is essential for germ cell development (Yamaji et al., 2008), whereas in humans, PRDM14 is thought to be involved in the epigenetic control of gene expression in undifferentiated ESCs (Tsuneyoshi et al., 2008). In this study, we found that knockdown of Prdm14 can significantly decrease the expression level of the apoptosis-related genes BCL-2 and the pluripotency-related genes OCT4 and SOX2, which are considered upstream factors of these genes.
In contrast, PRDM14 is thought to be involved in epigenetic modifications, such as histone modification and DNA methylation, by repressing DNMT or recruiting TET proteins (Okashita et al., 2014). DNA methylation usually acts cooperatively to regulate gene expression in mammals (Delcuve et al., 2009). Proper epigenetic modification is essential for the formation of healthy embryos. It is possible that PRDM14 activates the expression of these genes by promoter demethylation (Okashita et al., 2014). Hence, PRDM14 likely influences embryonic development by promoting active DNA demethylation in pluripotency-associated genes and apoptosis-related genes through the TET or DNMT pathway in parthenogenetically activated porcine embryos. Similar findings have been obtained in mouse studies (Nakaki and Saitou, 2014).
However, there was no difference in gene expression at the blastocyst stage, because gene silencing in mammalian embryos through the microinjection of siRNA was transient (Lochmatter and Mullis, 2011) and the duplexes were progressively diluted by consecutive cell divisions (Leung and Whittaker, 2005).
These results and our finding, that embryos from PRDM14 siRNA-injected oocytes tend to have lower cleavage and blastocyst rates, suggest that PRDM14 may play a role in embryonic development. As an alternative possibility, PRDM14 may regulate the expression of apoptosis and pluripotency-related genes through demethylation/remethylation of their promoters and thus influence the development and quality of parthenogenetically activated porcine embryos. The identification of a compound that interacts with PRDM14 could find the function of this transcription factor in mediating the critical stages of development from the oocyte to the blastocysts (Peng et al., 2012).
In conclusion, our data suggest that PRDM14 could affect embryonic development through regulating the expression levels of the pluripotency genes and antiapoptotic genes during the development of parthenogenetically activated porcine embryos.
Footnotes
Acknowledgments
This work was supported by grants from Natural Science Foundation of China (No. 31472093), National Science and Technology Major Projects (No. 2014ZX-08-007-002-008), and PCSIRT (IRT_16R32).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.