
Abstract
Abstract
Statins could reduce the risks of coronary heart disease death and ischemic cardiovascular events. In this study, we aim to explore the role of rosuvastatin in ischemia/reperfusion (I/R)-injured cardiomyocytes and the possible mechanism. An I/R model was established by oxygen-glucose deprivation/reperfusion (OGD/R). The protective effects of rosuvastatin pretreatment on OGD/R-injured cardiomyocytes were performed using MTT assay, lactate dehydrogenase (LDH) release assay, and quantitative real-time polymerase chain reaction (qRT-PCR). Bioinformatics software TargetScan and miRTarBase were used to predict the targeted miRNAs with uncoupling protein (UCP)2. Furthermore, verify the binding capacity of hsa-miR-24-3p and UCP2 with qRT-PCR and dual-luciferase reporter assay. The expression of UCP2, cell viability, LDH level, and apoptosis level affected by downregulated hsa-miR-24-3p were assessed using qRT-PCR, western blotting, MTT, the LDH kit, and flow cytometry. Pretreatment with rosuvastatin could significantly augment cell viability, reduce LDH level, increase the expression of UCP2, and downregulate hsa-miR-24-3p in OGD/R-injured H9c2 cells. miR-24-3p was closely connected with UCP2, and downregulated miR-24-3p could promote UCP2 expression, which presented cell viability increasing, LDH release and cell apoptosis inhibition in OGD/R condition. Moreover, it decreased the protein expression of Cleaved-Caspase-9 and Cyto C. This is the first time our study suggests that rosuvastatin pretreatment protects cardiomyocytes from OGD/R through upregulating UCP2 through downregulation of hsa-miR-24-3p.
Introduction
Coronary heart disease (CHD), the most common type of cardiovascular disease and the highest mortality rate in developed countries, is also the leading killer of health in many developing countries (Gaziano et al., 2010). In 2001, about 7.3 million people died of coronary heart disease worldwide. There were 110 million people with coronary heart disease, and 8.9 million of them died of coronary heart disease in 2015 (GBD 2015 Disease and Injury Incidence and Prevalence Collaborators, 2016). According to the World Health Organization, the number of deaths from coronary heart disease in the world accounts for about 13% of the total deaths by 2030 (Mathers and Loncar, 2006).
CHD, also known as ischemic heart disease, is a heart disease caused by myocardial ischemia and hypoxia due to abnormal coronary artery structure or function of the coronary arteries. Among them coronary atherosclerosis is the most common, mainly including stable angina pectoris, unstable angina pectoris, myocardial infarction, and sudden cardiac death (Wong, 2014). In the process of myocardial ischemia rescue and treatment, recovery of blood flow is the key to reduce myocardial damage; but after the ischemic tissue restores blood supply, the ischemic tissue will burst to generate excessive free radicals, making ischemic myocardial tissue more serious, which is usually called ischemia/reperfusion (I/R) injury (Jennings et al., 1960).
In 1986, Murry research group found in canine myocardial ischemia-reperfusion injury experiment, four short-term coronary artery occlusion followed by open could protect myocardium in subsequent ischemia, showing that myocardial infarction size was lower than that of simple ischemia group. And they put forward the concept of ischemic preconditioning for the first time (Murry et al., 1986).
To make up for the deficiency of ischemic post-treatment, drug preconditioning has developed gradually on the basis of ischemic preconditioning. Rosuvastatin, one of the statins, is an inhibitor of HMG-CoA reductase. Statins not only reduce the risk of coronary heart disease death, myocardial infarction, and readmission, but also reduce the risk of ischemic cardiovascular events (Baigent et al., 2005; O'Keefe et al., 2004; Ray et al., 2005; Schwartz et al., 2001; Spencer et al., 2004). However, the important molecular mechanism of statins for cardiac protection has not been fully elucidated.
Uncoupling proteins (UCPs) are a family of proteins located in mitochondrial intima and involved in proton transport (Ricquier and Bouillaud, 2000). UCP2 can mediate mitochondrial proton leakage, resulting in decreased adenosine triphosphate (ATP) production and reducing the formation of reactive oxygen species (Collin et al., 2005). It is closely related to atherosclerosis, diabetes, obesity, fatty liver, and other diseases (Cadenas, 2018; Liu et al., 2013; Zhou et al., 2016, 2018). Ryu had shown that UCP2 played a protective role in atherosclerosis by reducing the production of reactive oxygen species in macrophages (Ryu et al., 2004).
MicroRNA is a group of endogenous noncoding single-stranded RNA molecules (composed of 17–25 nucleotides). It binds to target mRNA 3′UTR by base complementary pairing and regulates gene expression at post-transcriptional level by degrading target mRNA (Tay et al., 2011). There are >1000 known miRNA genes that regulate >50% of human protein coding (Rodriguez et al., 2004). In recent years, studies have shown that the regulation of miRNA is ubiquitous in the process of cytopathology and pathophysiology (Vickers et al., 2014). Scientists have conducted a great deal of research on the mechanism of miRNA leading to cardiovascular disease and its role in the pathophysiological process of disease progression (van Rooij and Olson, 2012). As far as we know, Statins have been reported to protect myocardial cells from I/R damage by inhibiting miRNA (Zuo et al., 2016).
In this study, we aim to explore the role of rosuvastatin on myocardial cell injury induced by oxygen-glucose deprivation/reperfusion (OGD/R), as well as the underlying mechanism involved in miR-24-3p/UCP2 pathway.
Materials and Methods
Cell culture and OGD/R model
Rat cardiomyocytes (H9c2 cells) were obtained from Shanghai Yu Bo Biological Technology Co., Ltd (Shanghai, China). Cells were cultured at 37°C and 5% CO2 in Dulbecco's minimum essential medium (DMEM) (Thermo Fisher, Waltham, MA) with 10% fetal bovine serum (FBS, Thermo Fisher), 1% penicillin/streptomycin (Gibco, Thermo Fisher), and 2 mM glutamine. The media was changed every other day.
The cardiomyocytes were washed with phosphate-buffered saline (PBS) for three times, and the medium was replaced by FBS-free and sugar-free DMEM medium. The cells were placed in the incubator of 5% CO2 and 1% O2 cultured at 37°C for 6 hours, then changed with normal medium containing 5% CO2 for 1 hour. Cells included two types: treated with rosuvastatin (BioChemPartner, Shanghai, China) and without rosuvastatin treatment; then both cells were injured by OGD/R. The pretreatment group was DMEM high sugar medium adding rosuvastatin for 3 hours.
Cell viability assay
Cell viability was performed by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT; Solarbio Life Sciences, Beijing, China). H9c2 cells (1 × 104 cells/well) were seeded in 96-well plates and incubated for 24, 48, and 72 hours. Then cells were washed with PBS buffer and 20 μL MTT reagent was added to each well and cultured for another 4 hours. One hundred fifty microliters of DMSO was added after MTT was removed. The optical density was measured at 490 nm using a microplate reader (Thermo Fisher, MA).
Lactate dehydrogenase release assay
Under pathological conditions, the cell membrane structure was destroyed and lactate dehydrogenase (LDH) was released to the extracellular membrane through the damaged cell membrane. The activity of LDH in cell culture medium can be used as one of the indexes to judge the degree of cell injury. Cell suspension was centrifuged at 1000 r/min for 1 minute. Then the activity of LDH was detected by an LDH release assay kit (Beyotime, Shanghai, China).
Cell transfection
H9c2 cells were seeded in six-well plate (1.0 × 105) for 24 hours before transfection. Hsa-miR-24-3p inhibitor and negative control plasmids were purchased from QIAGEN. Lipofectamine 2000 (Invitrogen) was used to determine the transient transfection according to manufacturer's protocol. A total of hsa-miR-24-3p inhibitor, negative control, and Lipofectamine 2000 were added to Opti-MEM high-sugar medium and incubated at 25°C for 10 minutes, respectively. Then Lipofectamine 2000 was mixed into each well incubated with normal cultured H9c2 cells. After 3 hours culturing, the fluid was changed back to DMEM medium containing 10% FBS.
Luciferase assay
Either empty vector or miR-24-3p and luciferase reporter comprising 3′UTR of UCP2 wild type or mutant fragment (Gene Chem, Shanghai, China) co-transfected by Lipofectamine 2000 (Invitrogen) in 293T cells cultured in 96-well plate. Cells were harvested after transfection for 48 hours. Luciferase was then quantified using the Dual-Luciferase Reporter Assay (Promega, Madison, WI) according to the manufacturer's protocol.
Flow cytometry
The suspension was washed twice with cold PBS and then added Annexin V-FITC and propidium iodide (Sigma) at 4°C for 15 minutes. After cold binding buffer was added into per well, flow cytometry was performed to assess cell apoptosis within 1 hour.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from cells use of Trizol (Invitrogen) according to the manufacturer's protocol. Reverse transcription was performed with GoScript™ Reverse Transcription kit (Promega) at 37°C for 60 minutes and 4°C for 5 minutes. Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out on SYBR Fast qPCR Mix (Invitrogen, USA) for UCP2, hsa-miR-195-5p, hsa-miR-24-3p, hsa-miR-185-5p, hsa-miR-214-3p, and GAPDH. The primer sequences are listed in Table 1. Samples ran using the following cycling parameters: 95°C for 15 minutes, 94°C for 15 seconds, followed by 40 cycles of 60°C for 30 seconds and 70°C for 30 seconds. After completion of the PCR amplification, comparative quantification was determined using the 2−ΔΔCT method. Primers were synthetized commercially (BioSune Biotechnology, Shanghai, China).
Primers Used in Quantitative Real-Time Polymerase Chain Reaction
Western blotting analysis
Proteins were extracted from cells using RIPA lysis buffer (Beyotime). BCA protein kit (Beyotime) was used to determine the concentration of proteins. Aliquots protein was separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS–PAGE) and resolved proteins were transferred to polyvinylidene fluoride membranes (Millipore). Membranes were blocked in 5% milk PBS with 0.1% Triton X-100 and incubated with different primary antibodies: anti-UCP2 antibody (ab97931, 1:1000; Abcam), anti-Cleaved-Caspase-9 antibody (ab2324, 1:1000; Abcam), anti-cyto C antibody (ab90529, 1:1000; Abcam), anti-GAPDH antibody (ab9485, 1:2500; Abcam) overnight at 4°C.
The membranes were then incubated with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibody (Protein Tech). Protein bands were detected with enhanced chemiluminescence (ECL) (Thermo Fisher Scientific, Inc.) and visualized using Quantity one (Bio-Rad).
Statistical analysis
Statistical analysis was detected by Prism GraphPad version 6.0 software. All the data are presented as mean ± standard deviation. Differences were performed using one-way analysis of variance after Turkey's multiple comparison. A p < 0.05 was considered significant.
Results
Rosuvastatin alleviates OGD/R-induced cardiomyocytes injury
To study the effects of rosuvastatin on OGD/R-induced cardiomyocytes injury, the cell viability and LDH release level were detected by MTT and LDH release assay kit. As Figure 1A shows, H9c2 cell viability increased in a time-dependent manner. Rosuvastatin could partially reverse the effect of OGD/R on cell viability. LDH release was commonly considered as one of the indexes of cell injury. We could find OGD/R significantly increased LDH release (p < 0.01; Fig. 1B). However, with rosuvastatin pretreatment, the release of LDH presented a clear attenuation (p < 0.01).
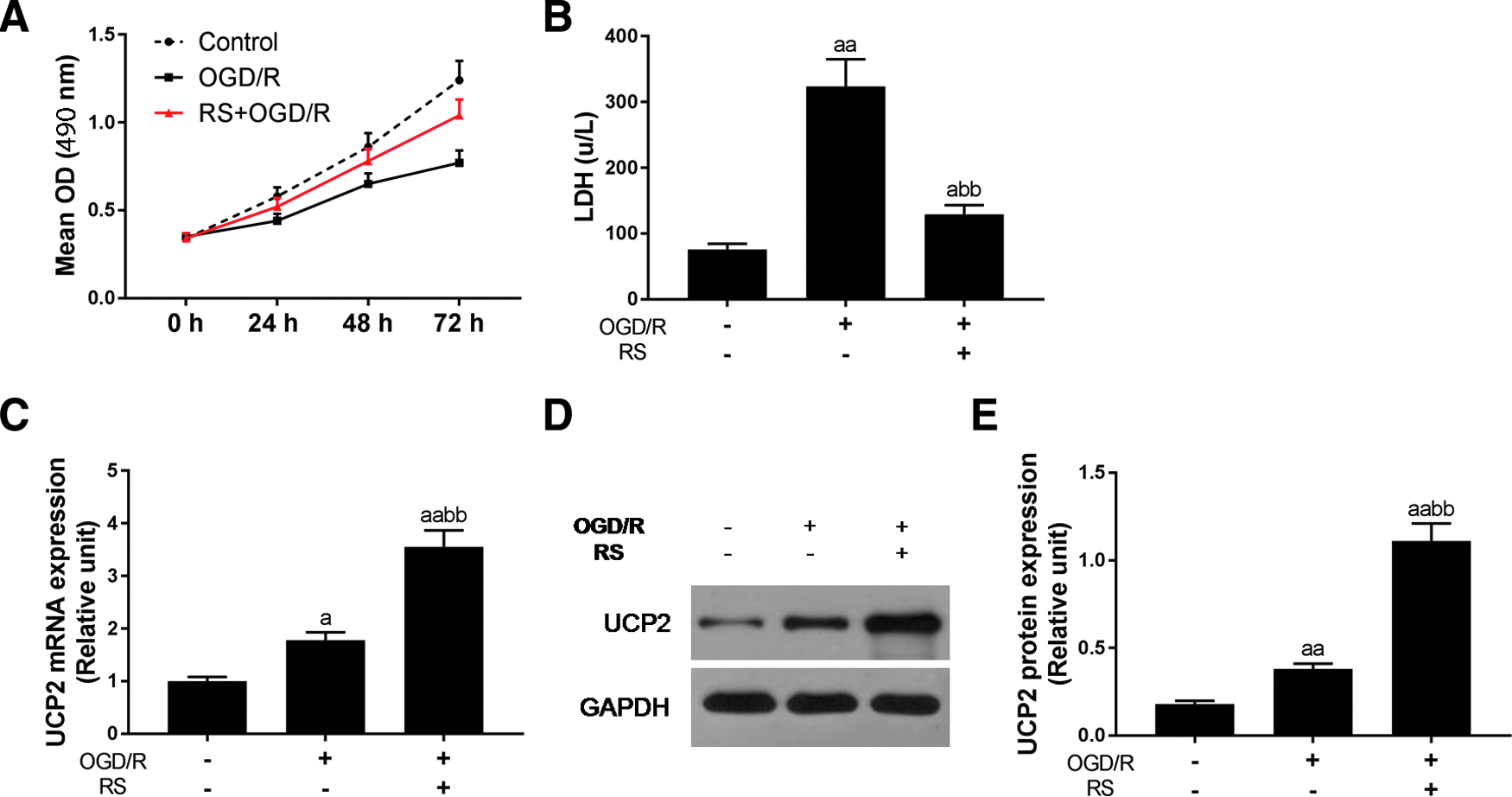
The effect of rosuvastatin on OGD/R-induced cardiomyocytes injury and the expression of UCP2.
Rosuvastatin augments the expression of UCP2
To explore how rosuvastatin act as the protective effect in H9c2 cells, we assessed the expression of UCP2. The qRT-PCR showed that both OGD/R model (p < 0.05) and rosuvastatin (p < 0.01; Fig. 1C) pretreatment could significantly increase the mRNA expression of UCP2 in comparison with control. The protein level of UCP2 was similar with mRNA (Fig. 1D, E). Moreover, the augment expression of UCP2 by rosuvastatin was stronger than OGD/R model (p < 0.01).
Prediction of the target relationship between hsa-miR-24-3p and UCP2
Using bioinformatics software TargetScan (www.targetscan.org/vert_72/) and miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/php/search.php) to predict the targeted miRNAs with UCP2, 312 and 40 miRNAs were obtained, respectively. As shown in Figure 2A Wayne diagram, the intersection of four microRNAs was obtained, namely, hsa-miR-195-5p, hsa-miR-24-3p, hsa-miR-185-5p, and hsa-miR-214-3p.

Prediction of the target relationship between hsa-miR-24-3p and UCP2.
Then we further selected the related microRNA in H9c2 cells by exploring the relationship with OGD/R and rosuvastatin. We found both in single OGD/R or rosuvastatin and OGD/R + rosuvastatin condition, the expression of hsa-miR-185-5p and hsa-miR-214-3p showed no clear difference with control (p > 0.05; Fig. 2D, E). In hsa-miR-195-5p mRNA level, it revealed significant high expression exposed to OGD/R (p < 0.01; Fig. 2B). The group with rosuvastatin pretreatment, the expression of hsa-miR-195-5p completely decreased in comparison with OGD/R alone (p < 0.01). The decrease of the hsa-miR-195-5p expression was stronger by rosuvastatin than that by OGD/R + rosuvastatin (p < 0.01).
As Figure 2C shows, the hsa-miR-24-3p commonly existed in normal H9c2 cells, when OGD/R-injured cells, the expression of hsa-miR-24-3p significantly downregulated (p < 0.05; Fig. 2C).
Moreover, adding to rosuvastatin, the hsa-miR-24-3p expression attenuated more pronounced (control, p < 0.01). Luciferase reporter subcloned with the wild or mutant sequence of UCP2 and then co-transfected with hsa-miR-195-5p (Fig. 2F, H), hsa-miR-24-3p (Fig. 2G, I), or miR-NC into 293T cells. In regard to hsa-miR-195-5p, the result showed that both UCP2-3′-UTR and UCP2-3′-UTR mut had no obvious difference with control (Fig. 2F). When hsa-miR-24-3p was co-transfected into 293T cells, the relative luciferase activity of UCP2-3′-UTR was significantly decreased (p < 0.01; Fig. 2G), but the relative luciferase activity of UCP2-3′-UTR mut had no effect by co-transfection with hsa-miR-24-3p (p > 0.05).
The effect of downregulated hsa-miR-24-3p on the expression of UCP2
When miR-24-3p inhibitor was transfected into H9c2 cells, the expression of miR-24-3p sharply decreased in comparison with control or NC (p < 0.01; Fig. 3A). In addition, under OGD/R-injured condition, both rosuvastatin and the miR-24-3p inhibitor could obviously decrease the miR-24-3p expression, and the decrease effect was more significant when rosuvastatin combination with inhibitor (p < 0.01; Fig. 3B). Compared with single OGD/R-injured group, both rosuvastatin pretreatment and inhibitor treatment, the mRNA expression of UCP2 completely increased (p < 0.01; Fig. 3C). When rosuvastatin and inhibitor were combined in the OGD/R condition, the UCP2 mRNA expression showed higher (p < 0.01). The protein level of UCP2 was in agreement with mRNA level (Fig. 3D, E).
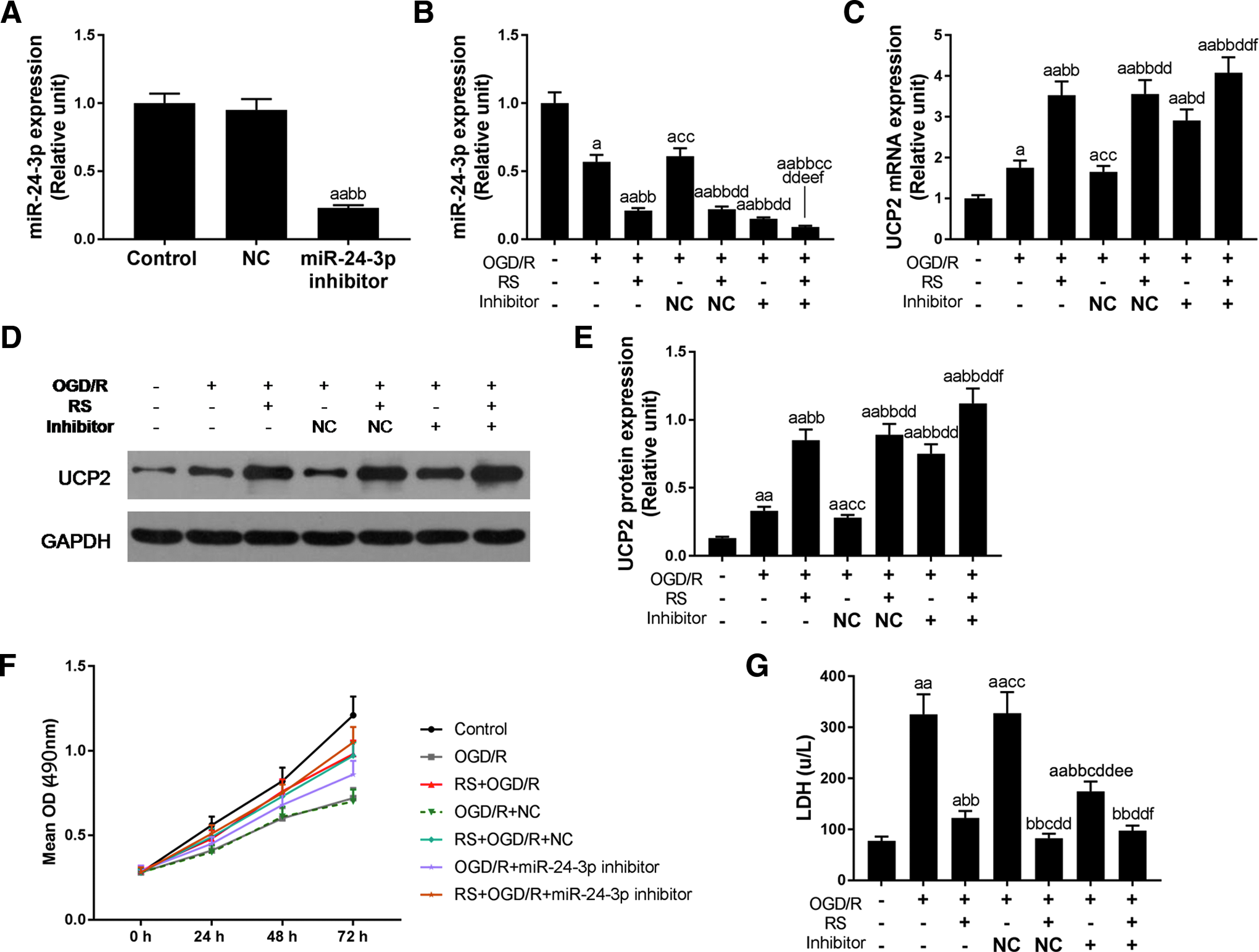
The effect of downregulated miR-24-3p on the expression of UCP2, cell viability, and LDH release in H9c2 cells.
The effect of downregulated hsa-miR-24-3p on the cell viability and LDH release in H9c2 cells
MTT assay was performed to detect the effect of downregulated miR-24-3p on the cell viability. Both rosuvastatin and inhibitor could enhance cell viability in OGD/R-injured H9c2 cells (Fig. 3F). Moreover, when rosuvastatin and inhibitor were combined, decreased cell viability induced by OGD/R would reverse. As Figure 3G shows, miR-24-3p inhibitor could significantly attenuate LDH release induced by OGD/R (p < 0.01; Fig. 3G). However, the effect of decreased LDH release was weaker than rosuvastatin (p < 0.01). When rosuvastatin was combined with miR-24-3p inhibitor, LDH release level was significantly reduced in OGD/R injured cells compared to that of inhibitor treatment alone (p < 0.01).
The effect of downregulated miR-24-3p on cell apoptosis
As Figure 4A shows, cell apoptosis was detected by flow cytometry. The result showed that rosuvastatin could significantly decrease cell apoptosis induced by OGD/R (p < 0.05; Fig. 4A). miR-24-3p inhibitor had an inconspicuous attenuation cell apoptosis coursed by OGD/R. When rosuvastatin and inhibitor were combined, OGD/R promoted H9c2 cell apoptosis effect would remarkably inhibit (p < 0.01).
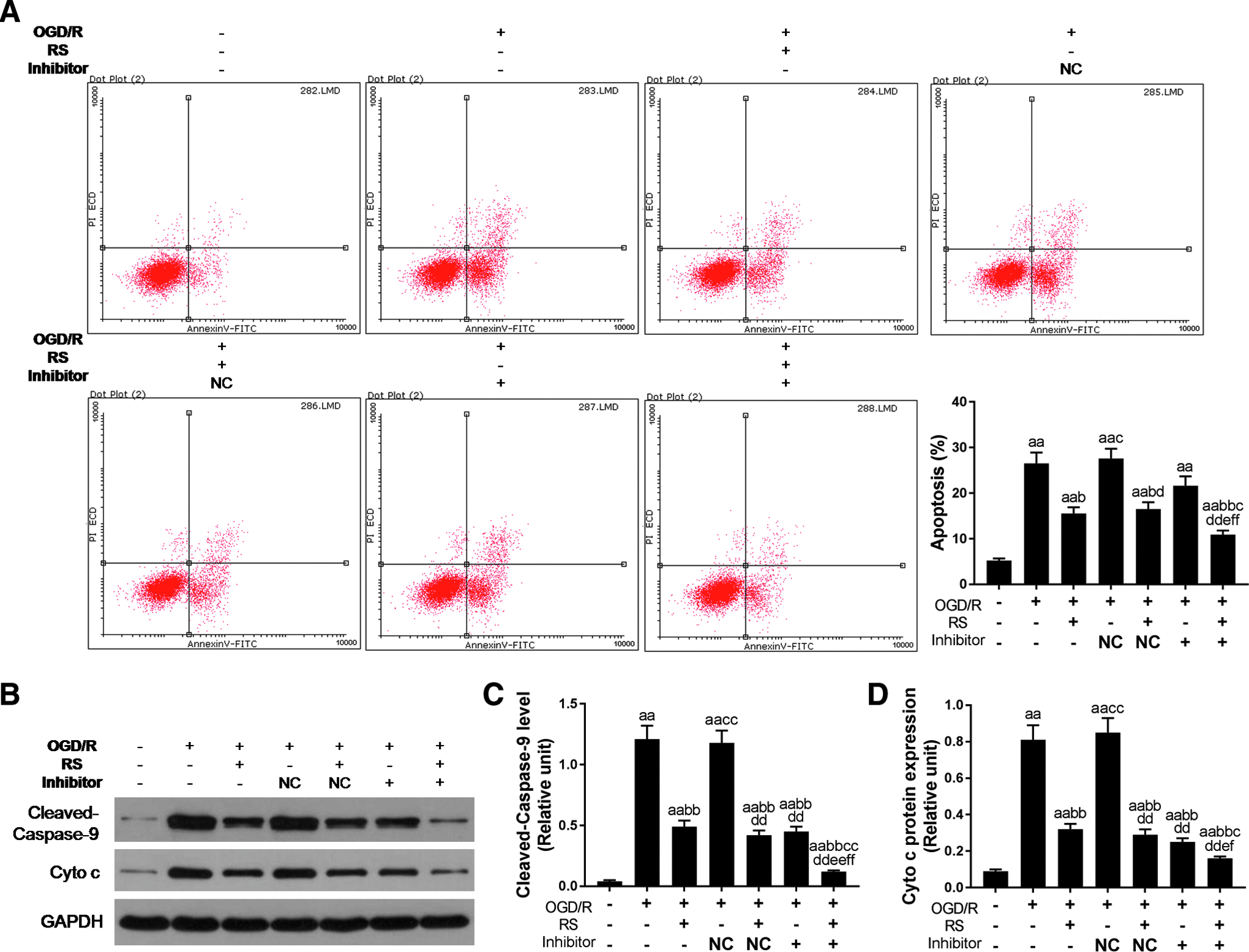
The effect of downregulated miR-24-3p on cell apoptosis.
Furthermore, western blotting was performed to detect apoptosis-related protein Cleaved-Caspase-9 and Cyto C (Fig. 4B). Under OGD/R condition, the Cleaved-Caspase-9 level showed significant high expression (p < 0.01; Fig. 4C) and Cyto C release also remarkably increased (p < 0.01; Fig. 4D). The apoptosis-related proteins, cleaved-caspase-9 and cyto C expressions were decreased not only by rosuvastatin treatment alone but also by miR-24-3p inhibitor treatment alone. Moreover, the attenuation expression effect was more obvious when rosuvastatin and miR-24-3p inhibitor were combined in OGD/R-injured H9c2 cells (p < 0.01).
Discussion
In recent years, the application of coronary intervention, thrombolytic therapy, and cardiopulmonary bypass has made a major breakthrough in the treatment of ischemic heart disease (Fanari et al., 2015; Kanmatsuse et al., 1997; Svagzdiene et al., 2015). These treatments undergo a process of blood flow reperfusion after ischemia, which usually leads to increased myocardial injury, increased infarct size, and arrhythmia (Roffi et al., 2016). In this study, we applied oxygen-glucose deprivation and reperfusion to establish myocardial injury model. Cell viability inhibition, LDH release augment, and apoptosis increasing proved that myocardial ischemia-reperfusion model was established successfully in rats. It is consistent with the report that LDH commonly was used to judge the extent of cell injury (Liu et al., 2018).
Ischemic preconditioning is the most effective protective intervention for myocardial I/R damage (Shen et al., 2008). Then researchers found that a variety of drugs can mimic IPC to produce exact myocardial protective effects, such as volatile anesthetics sevoflurane, opioid sedatives, and statins (Lee et al., 2018; Zhong et al., 2017). Statins have protective effects on important organs. Recent experimental studies have found that short-term preconditioning with statins can reduce heart, brain, and kidney injury caused by ischemia-reperfusion (Birnbaum et al., 2003; Naidu et al., 2003; Yokota et al., 2003).
In different I/R models, atorvastatin pretreatment could protect cardiomyocytes by activation of mitochondrial KATP channel (Zhao et al., 2015). Hu et al. (2013) also suggested that rosuvastatin could protect heart from ischemic injury as well as preserve the cardiac function. In our study, rosuvastatin increased cell viability, reduced LDH release, as well as promoted cell apoptosis inhibition in OGD/R-injured H9c2 cells. Moreover, it could induce the expression of UCP2 in mRNA and protein level.
The pathogenesis of ischemia-reperfusion injury is complicated. It is thought to be related to free radical injury, calcium overload, energy metabolism disorder, apoptosis, leukocyte-mediated inflammation, and microcirculation disorder (Arslan et al., 2008; Cao et al., 2009; Kaul et al., 1993; Kohler et al., 2011; Levi et al., 1997; Shiroto et al., 2005).
In general, UCP2 can regulate the proton leakage of mitochondrial membrane potential and change mitochondrial membrane potential, thus regulating ATP synthesis and ROS level (Pecqueur et al., 2001). Studies have shown that UCP2 regulates ROS levels and exogenous peroxides can activate the proton leakage dependent on UCP2. When the UCP2 gene was knocked out, the levels of ROS in islet cells and macrophages increased significantly (Brand et al., 2004; Krauss et al., 2002). In view of the important role of UCP2 in energy metabolism and ROS production, UCP2 may be an attractive target for prevention and treatment of ischemia-reperfusion injury.
In addition, we used bioinformatics software TargetScan and miRTarBase to predict the targeted miRNAs with UCP2. The importance of miRNA to the cardiovascular system has been confirmed in murine cardiomyocyte-specific deletions of Dicer and Dgcr8 (Olson, 2014). Our result showed that UCP2 and hsa-miR-24-3p had stronger ability to combine, as performed by qRT-PCR and dual-luciferase reporter assay. Furthermore, the miR-24-3p inhibitor was transfected into H9c2 cell to render its low expression. In OGD/R model, the expression of miR-24-3p was decreased by rosuvastatin or miR-24-3p inhibitor alone, and the decrease expression effect was more significant when rosuvastatin was combined with miR-24-3p inhibitor.
In regard to the expression of UCP2, we found that not only rosuvastatin but also miR-24-3p inhibitor could increase the mRNA and protein expression of UCP2 in OGD/R-induced cells. Without surprise, rosuvastatin in combination with miR-24-3p inhibitor could significantly induce UCP2 expression. Taken together, rosuvastatin protected rat cardiomyocytes from OGD/R injury by downregulation of hsa-miR-24-3p to target the upregulated UCP2 expression.
In the heart of adult mice, miR-1 accounts for 40% of the total miRNAs, suggesting that it plays a key role in maintaining cardiac function (Rao et al., 2009). miR-26 plays an important role in cardiovascular repair (Icli et al., 2014). The miR-181 family inhibits vascular inflammation (Sun et al., 2014). miR-29 regulates a variety of extracellular matrix proteins and involves a variety of fibrotic diseases.
We also found that downregulated hsa-miR-24-3p could enhance cell viability, reduce LDH release, and attenuate cell apoptosis through suppressing Caspase-9 activation and Cyto C expression in OGD/R-induced cells. Moreover, adding to rosuvastatin, the effect of downregulated hsa-miR-24-3p on OGD/R-induced cells could be more significant. Meloni study showed that the expression of miR-24 was decreased in infarcted area, peripheral cardiomyocytes, and fibroblasts, but increased in vascular endothelial cells. In vitro, inhibition of miR-24 enhances the activity, proliferation, and capillary network of human vascular endothelial cells by upregulating the expression of eNOS gene (Meloni et al., 2013).
Our results would be more convincing if we used another miR-24 inhibitor to demonstrate its role in this study. A previous study has also reported that miR-24 was upregulated in failing cardiomyocytes (Xu et al., 2012) and miR-24 antagonism was documented to prevent ischemia reperfusion injury in renal cells (Lorenzen et al., 2014). These studies were consistent with our results.
Collectively, this is the first time our study suggests that rosuvastatin pretreatment protects cardiomyocytes from OGD/R through upregulating UCP2 through downregulation of hsa-miR-24-3p.
Footnotes
Author Disclosure Statement
The authors declare there are no conflicting financial interests.