
Abstract
Abstract
The objective of the present work was to investigate the effect of Periostin (POSTN) silencing on autophagy in osteoblasts, and provide an experimental basis for studying the mechanism of dental eruption. The cells were divided into the following four groups according to their viral number: the NC group, pFU-GW-016PSC53349-1; group KD1, LVpFU-GW-016PSC66471-1; group KD2, LVpFU-GW-016PSC66472-1; and group KD3, LVpFU-GW-016PSC66473-1. The lentiviral vector was infected at MOI = 100 in the ENi.S medium containing 5 g/mL Polybrene. The target gene expression was observed by a Celigo® Image Cytometer at 72 hours after infection, and the positive rate of fluorescence was noted. A two-step method of quantitative real-time PCR (qRT-PCR) was used to detect the silencing effect of POSTN. Western blotting was then performed to assess the expression of autophagy-related proteins Beclin-1 and LC3 in the group showing the best gene silencing effects. The experimental results showed that there was strong green fluorescence in group KD3. As confirmed via qRT-PCR analysis, the POSTN silencing efficiency in group KD3 reached 92.1%. The Western blotting revealed that the expression of Beclin-1 protein in group KD3 was significantly higher than that in the NC group. However, the LC3 protein expression was not significantly different from that of the control group. The lentiviral vector targeting POSTN in osteoblasts was constructed successfully. In addition, the expression of autophagy protein in mouse osteoblasts increased after POSTN silencing. This finding may provide new approaches for understanding the molecular signal transduction of POSTN during the tooth eruption process.
Introduction
The formation of tooth eruption channels is the premise of normal tooth eruption (Shastri et al., 2014; Tuncer et al., 2017). This task is performed directly by osteoclasts. Therefore, the functional changes of osteoclasts have always been the focus of tooth eruption research (Oosterkamp et al., 2014; Rahul et al., 2018). However, few studies have attempted to investigate how osteoblasts initiate osteoclasts aggregation and differentiation, promote the formation of the eruption channel, or explored whether autophagy occurs during this process. Periostin (POSTN) is a multifunctional soluble intercellular matrix protein that is mainly expressed in the periosteum and regulates osteoblasts to increase value, differentiation, and adherence (Uysal et al., 2017; Zeng et al., 2018).
Previous studies have shown that cortical bone density and bone strength have decreased in POSTN-knockout mice, but the specific mechanism is not yet clear (Bonnet et al., 2013). Our previous studies have shown that osteoblasts themselves robustly induce autophagy (Qin et al., 2016). However, the effect of autophagy on osteoblasts during tooth eruption and the association between this effect and tooth eruption are unknown. Therefore, the present study silenced POSTN via lentiviral transduction, and then assessed the expression of autophagy-related proteins. The aim is to investigate the effect of POSTN silencing on autophagy in mouse osteoblasts. Our findings may provide new approaches for studying the molecular signal transduction mechanisms and the dynamic balance between osteoblasts and osteoclasts during the tooth eruption process.
Materials and Methods
In vitro culture of osteoblast cell line
The mouse osteoblast-like (MC3T3-E1) cells in a frozen storage tube were removed from the liquid nitrogen jar, put quickly into a 37°C water bath, and shaken occasionally to thaw. After completely thawing, the cells were centrifuged at 1300 rpm for 3 minutes, and then the frozen tube was disinfected with 75% alcohol and moved to the biological safety cabinet. The supernatant was aspirated, and 1 mL fresh complete medium was added. The cell suspension was transferred to 6 cm dish containing 3 mL of complete medium, and the cells were incubated at 37°C, with 5% CO2 in static culture. After 24 hours, the culture medium was replaced. The cells were subcultured at ∼80% to maintain a good growth state.
Lentivirus-infected target cells
Cells in logarithmic growth were digested with pancreatic enzymes, and a cell suspension (3–5 × 104/mL) was prepared with complete medium. The cells were then inoculated onto culture plates and allowed to develop colonies. The lentivirus infection was conducted at the target cells when the cell density reached ∼15–30%. Based on the results obtained in the preliminary experiments, LVpFU-GW-016PSC66471-1, LVpFU-GW-016PSC66472-1, and LVpFU-GW-016PSC66473-1 lentiviral vectors were infected for 10 hours in the ENi.S medium containing 5 g/mL Polybrene (MOI = 100). The medium was then replaced with a conventional growth medium, and the cells were cultured for an additional 16 hours after infection. The target gene expression was observed by Celigo® Image Cytometer at 72 hours after infection, during which the positive rate of fluorescence was noted.
Real-time PCR
Cells were centrifuged at 1300 rpm for 3 minutes. The supernatant was aspirated and 1 mL TRIzol was added. The cells were then placed in static culture conditions for 5 minutes. The lysate was transferred to a new 1.5 mL EP tube. cDNA was obtained by reverse transcription (using the Promega M-MLV Kit). Then, 1 μL Oligo dT (0.5 μg/μL) and 2.0 μg total RNA was added to the PCR tube, and RNase-free H2O was added to achieve a final volume of 10 μL. The samples were thoroughly mixed and then centrifuged before incubation in warm water bath at 70°C for 10 minutes. The samples were immediately transferred into ice water to facilitate the annealing of oligo dT and the DNA template.
After mixing and short centrifugation, the system was then subjected to a water bath reaction at 42°C for 1 hour, after which, the reaction was halted by inactivating the RT enzyme in a subsequent water bath reaction at 70°C for 10 minutes. The obtained RT product (-cDNA) was stored at −20°C. Then, two-step methods of real-time PCR were used to detect and analyze the silencing effect of POSTN. The design and synthesis of internal reference genes and target gene primers were accompleted by Shanghai Genechem Co., Ltd.
The detailed information is provided below. The primer information for GAPDH is as follows: the upstream primer sequence is TGGTGAAGGTCGGTGTGAAC, the downstream sequence is GCTCCTGGAAGATGGTGATGG, and the amplification fragment size (bp) is 231. The primer information for POSTN is as follows: the upstream primer sequence is TTTCACCGACCTGGTAGCC, the downstream sequence is TTGGTCCATGCTCAGAGTGTC, and the amplification fragment size (bp) is 118.
Western blotting
The MC3T3-E1 cells in the group of best gene silencing effects were harvested, washed two times with precooled PBS and then lysed in extraction buffer. The cell samples were then centrifuged at 4°C for 15 minutes. The protein concentrations of the cell lysates were quantitated using the BCA Protein Assay Kit. Different concentrations of gel were prepared based on the molecular weight of the target protein. Equal protein amounts were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto a polyvinylidene difluoride (PVDF) membrane.
After blocking in sealing solution for 1 hour, the membrane was incubated overnight with rabbit anti-Beclin-1 (Abcam Company; NO ab207612, 1:300, 52 kDa) and anti-LC3 (CST Company; NO #12741, 1:300, 14, 18 kDa) polyclonal antibody at 4°C. The PVDF membrane was then reacted with horseradish peroxidase-conjugated anti-rabbit IgG antibody (Santa Cruz Company; NO sc-2004, 1:2000) for 2 hours at room temperature.
After a thorough washing with TBST, the PVDF film was placed on a cling film, and the mixture was dropped uniformly onto the PVDF film for 5 minutes so as to avoid light reaction. The membrane was then transferred into the cassette and exposed for 1–2 minutes after the excess ECL substrate reactive fluid was drained. The X-ray film was then taken out and put into the developing liquid for ∼1 minute. The film was then rinsed in clear water for a few seconds, and then transferred into the fixing liquid and kept there for at least 2 minutes. The X-ray film was then taken out and dried for further analysis. GAPDH (Santa Cruz Company; NO sc-32233, 1:2000, 36 kDa) was also detected on the same membrane and used as the loading control.
Statistical analysis
All experimental data are presented as the mean ± SEM. Statistical significance was determined by the one-way analysis of variance test and significance was considered at p < 0.05.
Results
Lentiviral transduction of MC3T3-E1 cells
The target gene expression was observed by Celigo Image Cytometer at 72 hours after infection, and the positive rate of fluorescence was noted. There was no green fluorescence in group NC and group KD2; a small amount of fluorescent expression in group KD1 and obvious green fluorescence in group KD3 (Fig. 1). The result of real-time PCR amplification curve showed that all amplification curves were smooth, and the curves of each tube were parallel (Fig. 2). The results of the melting curves of GAPDH and POSTN show that they were single-peak type, the base was narrow and the peak height was high (Fig. 3). The 2−ΔΔCt method of data analysis was used in the present study. From the quantitative real-time PCR (qRT-PCR) results, it can be seen that POSTN silencing efficiency was as high as 92.1% (Table 1 and Fig. 4).
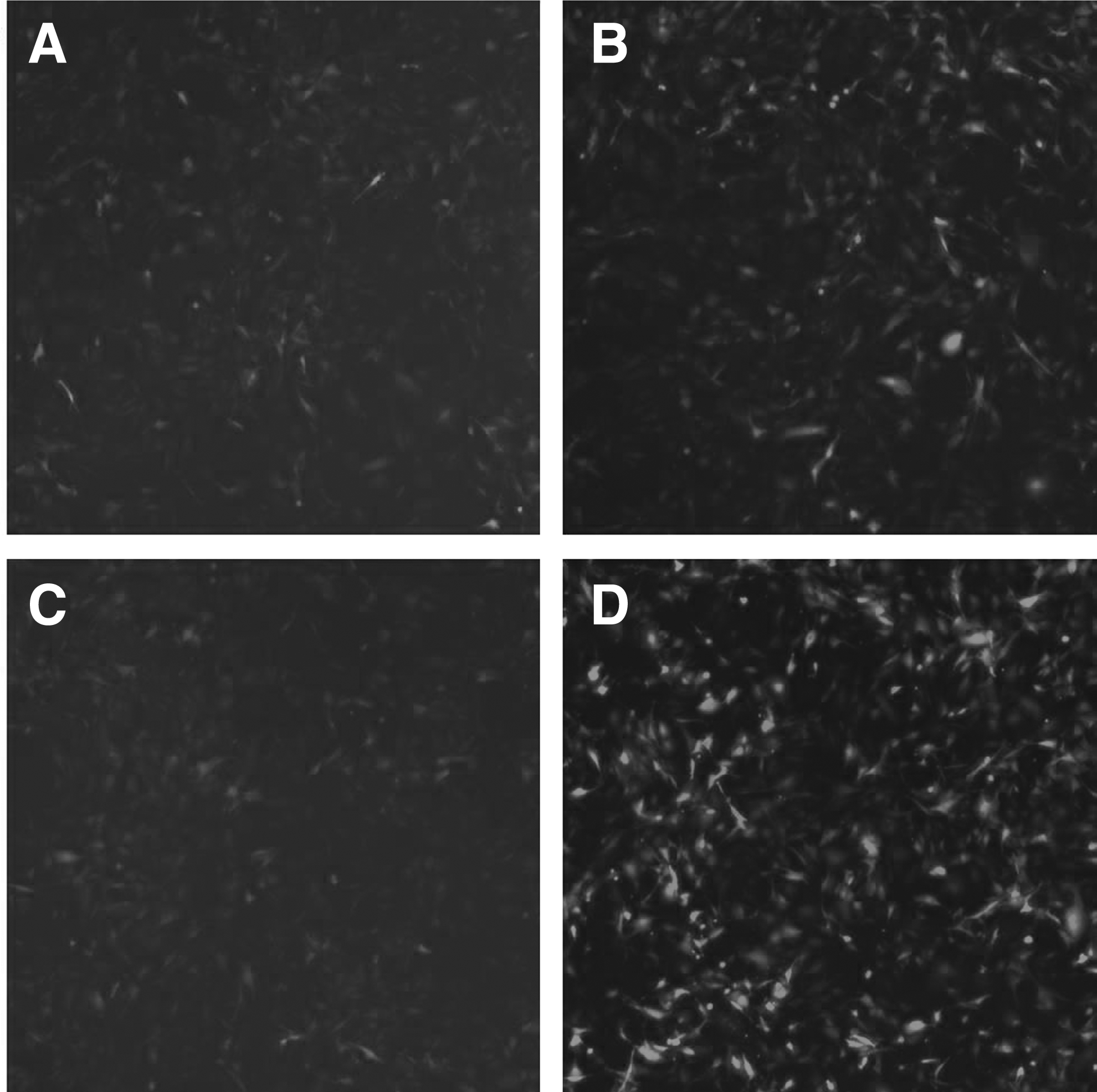
The target gene expression was observed by Celigo® Image Cytometer at 72 hours after infection, and the positive rate of fluorescence was noted. There was no fluorescence in group NC and group KD2; a small amount of fluorescent expression in group KD1 and obvious fluorescence in group KD3.
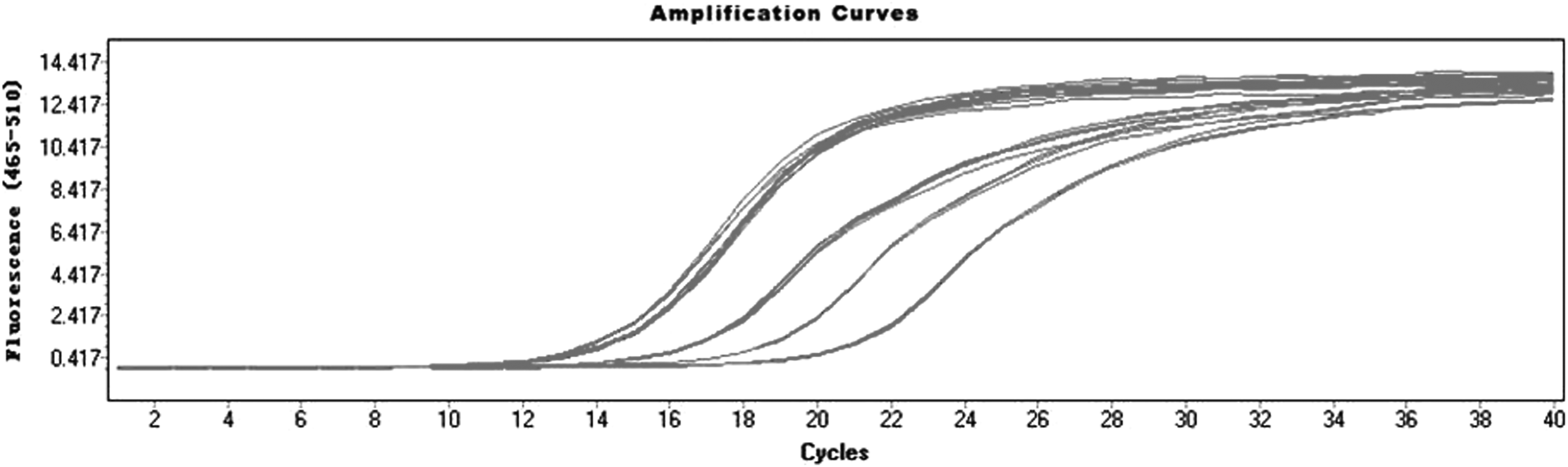
All amplification curves were smooth, the curve inflection point was clear, and the curves of each tube were parallel, indicating that the amplification effect was similar.
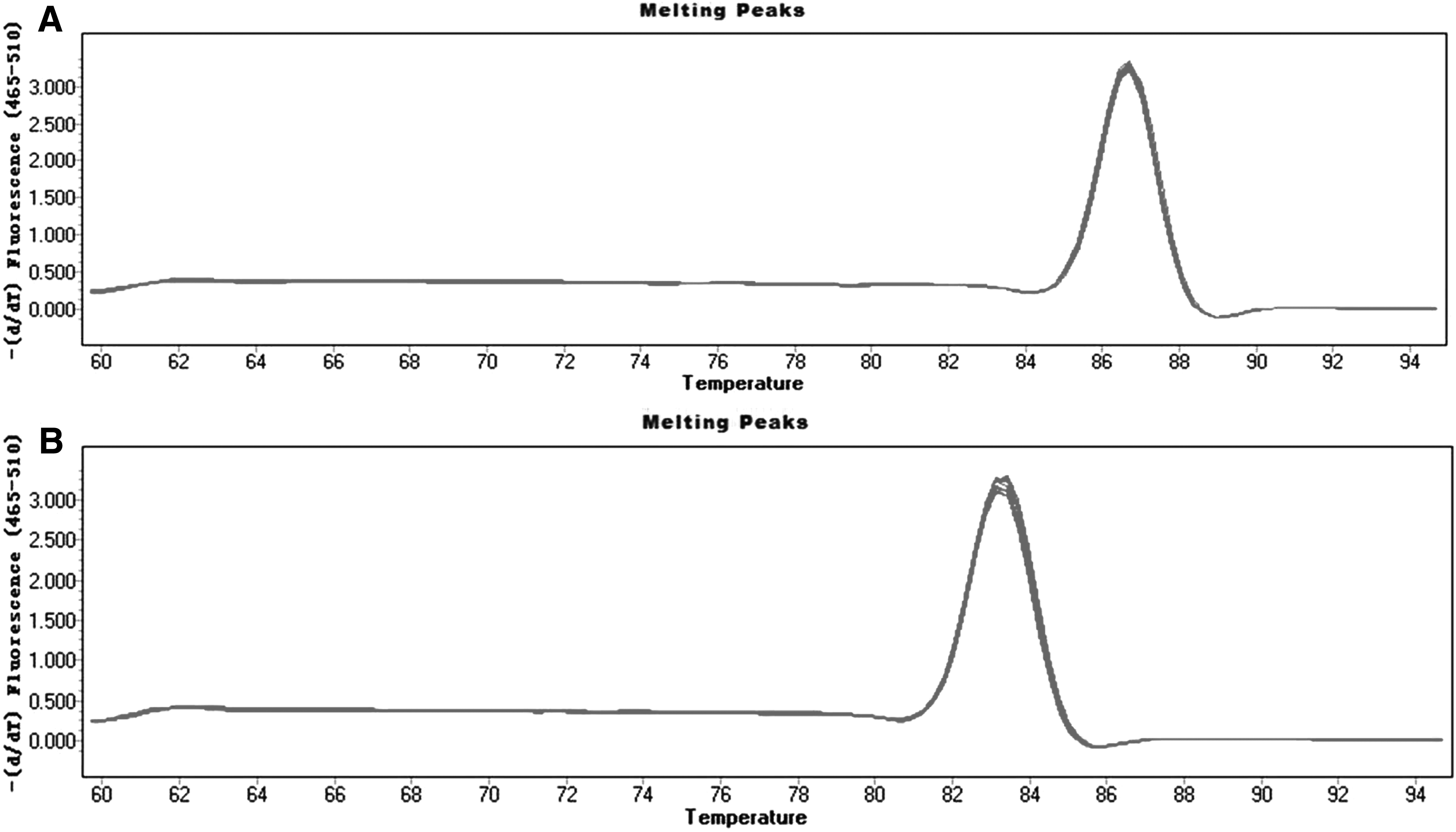
The melting curves of GAPDH and Periostin were single-peak type, the base was narrow and the peak height was high, indicating that there was no primer-dimer or nonspecific amplification. This indicated that the primer design was good and the specificity was strong.
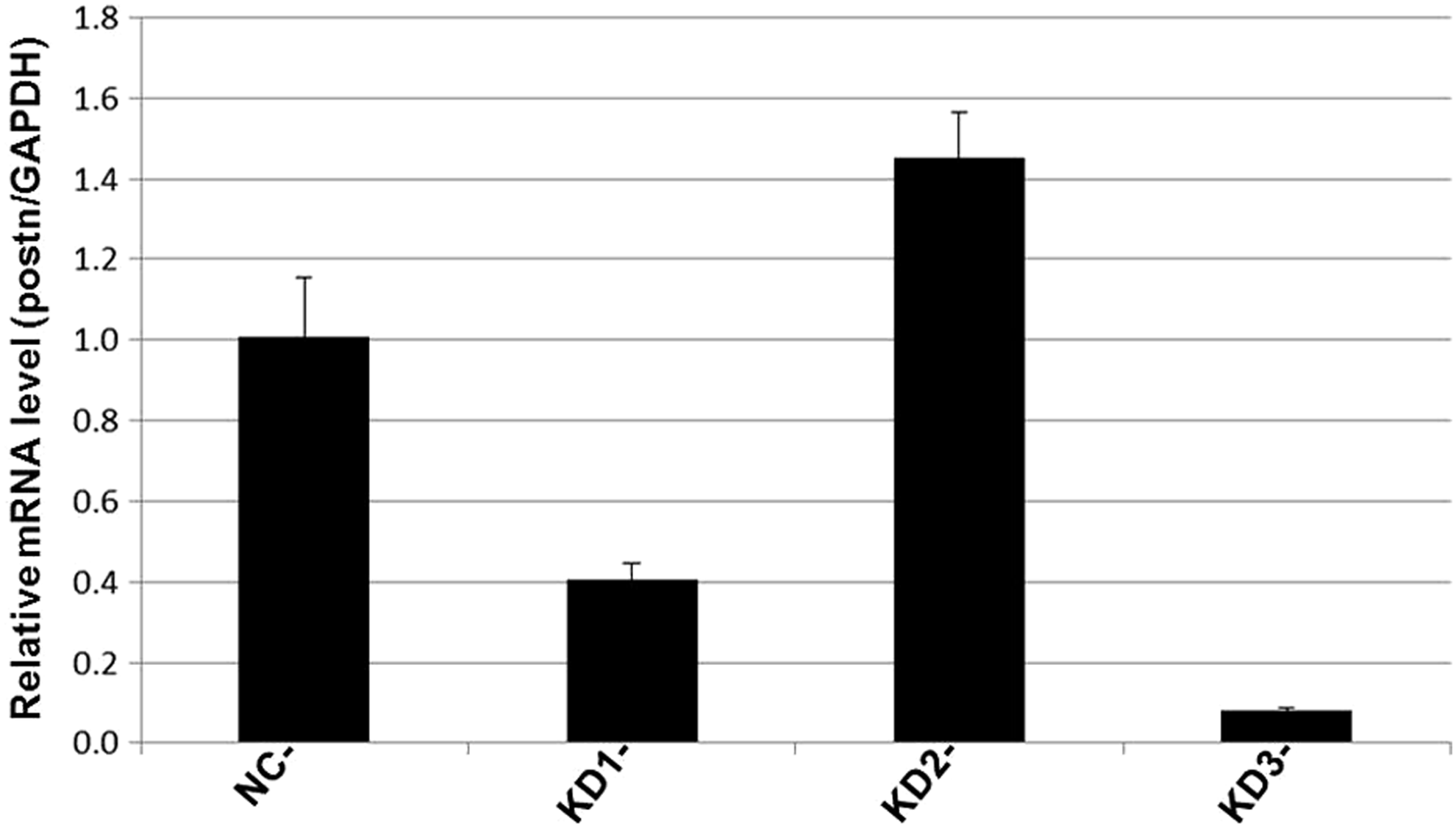
Data analysis: Using the 2−ΔΔCt method of data analysis, ΔCt = target gene Ct value - internal reference gene Ct value; −ΔΔCt = means value of ΔCt in NC group −ΔCt values of each sample; 2−ΔΔCt reflects the relative expression level of the target genes of the samples relative to group NC. From the quantitative PCR results, it can be seen that in MC3T3-E1 cells, compared to group NC, the Periostin silencing efficiency was higher in group KD3 (p < 0.05).
Analysis of the Periostin Real-Time Polymerase Chain Reaction Results in Each Group
Western blotting
Beclin-1 was significantly upregulated in the experimental group compared with those in the control group (Figs. 5 and 6). However, LC3 expression levels remained largely unaltered between the control and experimental groups (Figs. 7 and 8).
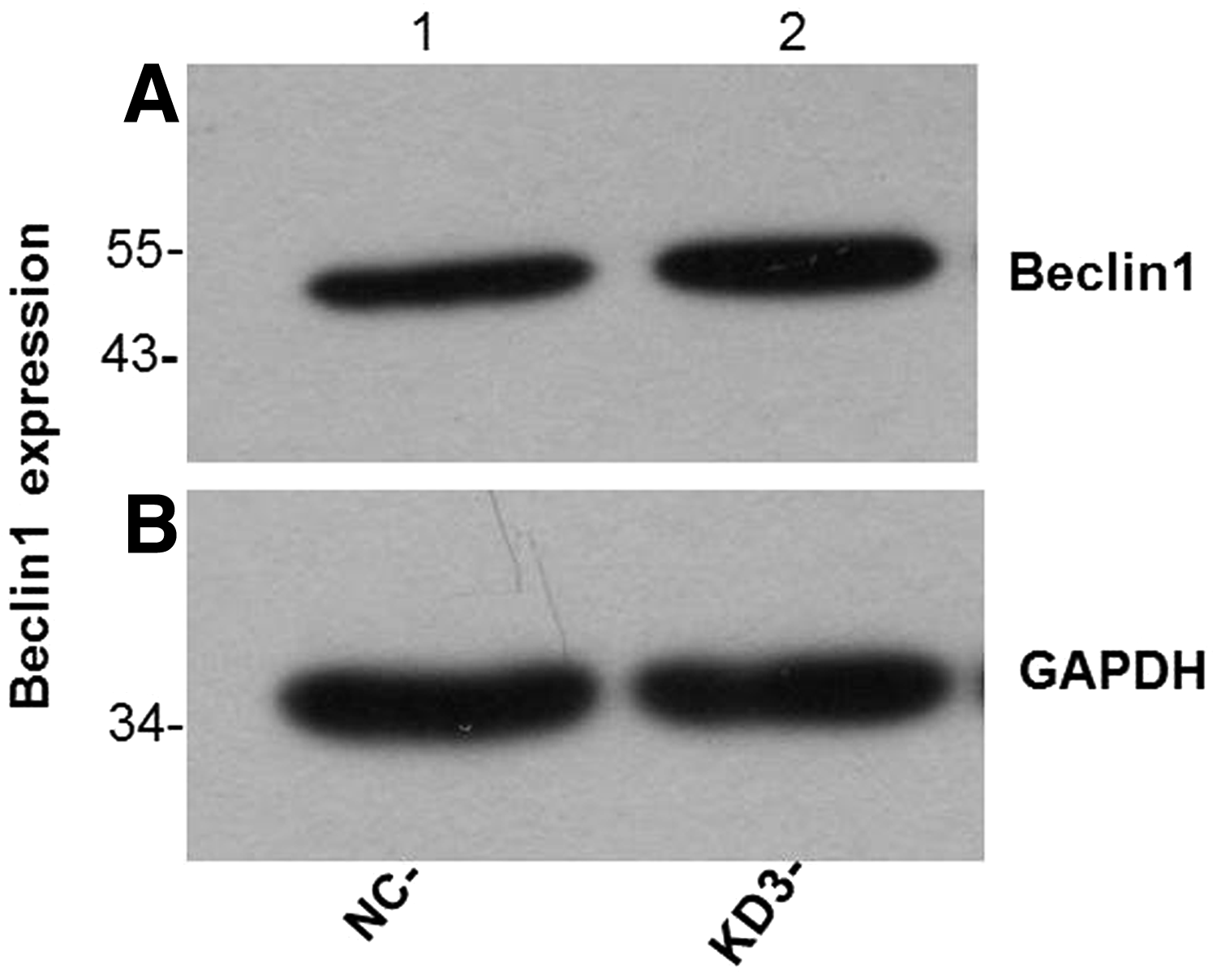
Western blotting results showed that the expression of Beclin-1 protein in group KD was higher than that in group NC.
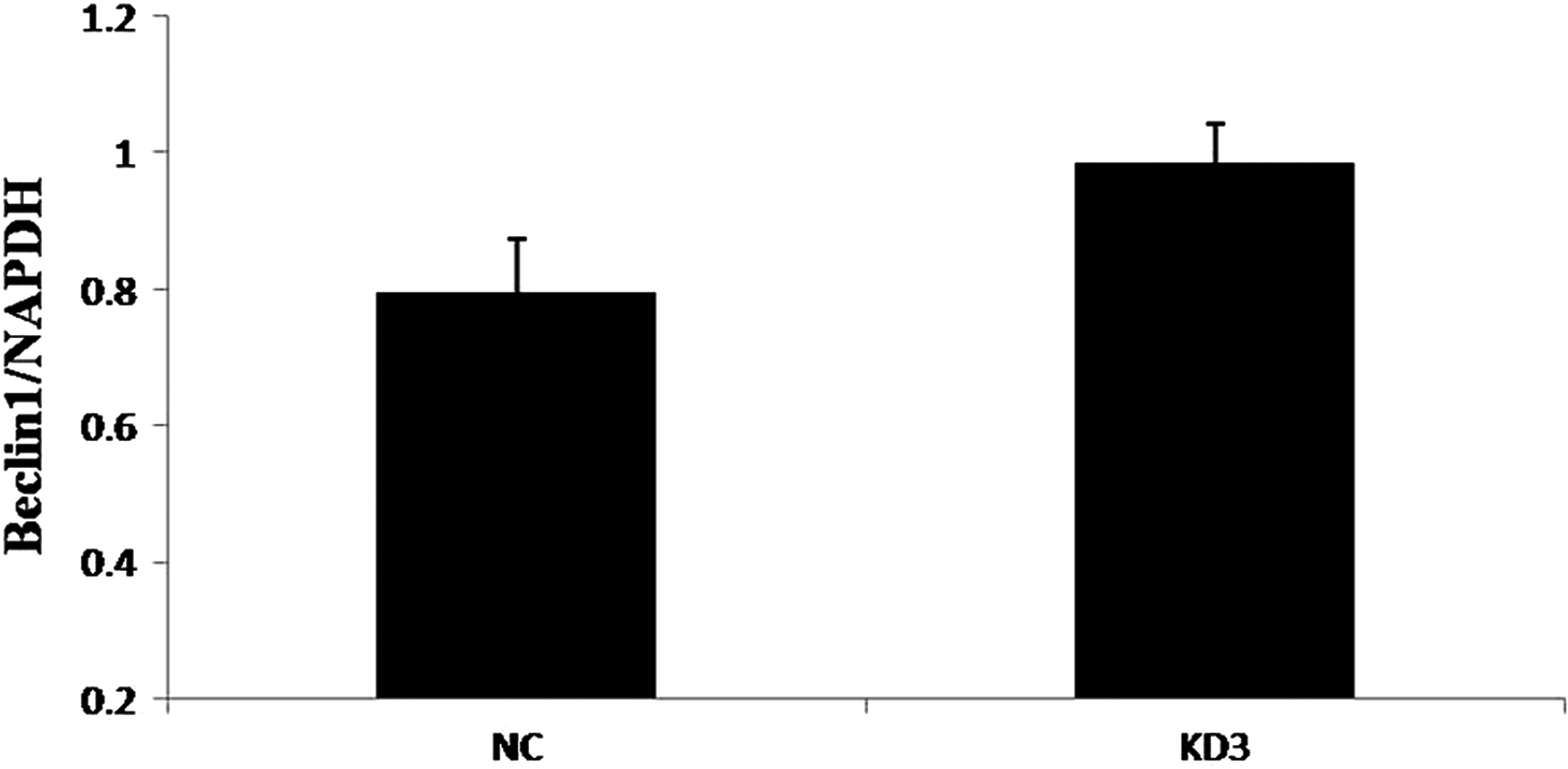
Compared to group NC, the expression of Beclin-1 protein in group KD was higher than that in group NC (p < 0.05).
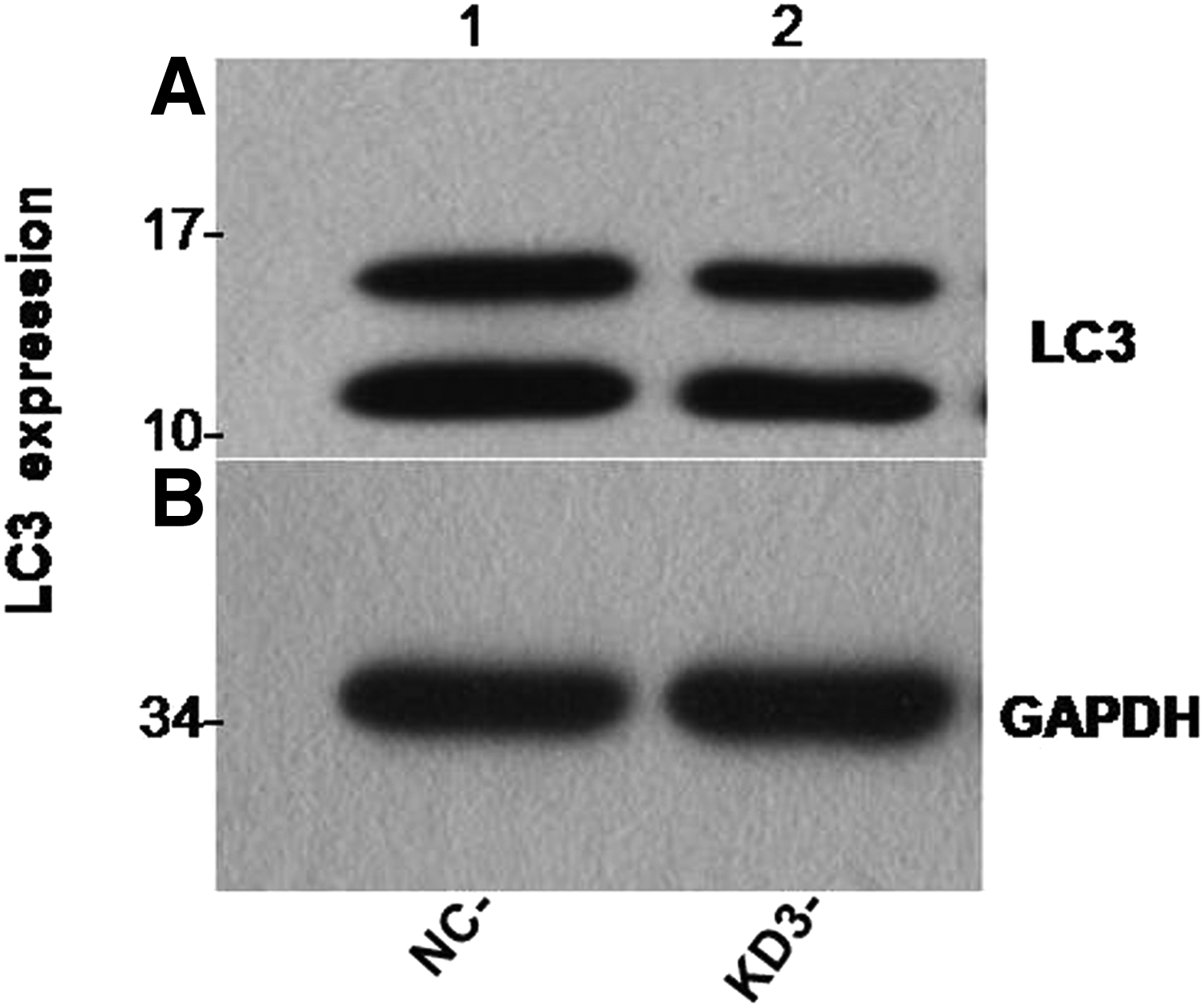
Western blotting results showed that there was no significant change in LC3 protein expression between group KD and group NC.

There was no significant change in LC3 protein expression between group KD and group NC (p > 0.05).
Discussion
POSTN can promote both osteoblast differentiation and adhesion, and it plays an important role in maintaining the integrity of periodontal tissue, tooth development, and eruption, but the specific mechanism is not yet clear (Gerbaix et al., 2015; Wang et al., 2016). During the tooth eruption process, osteoblasts and osteoclasts need to maintain a dynamic balance. If this balance is uneven, it may lead to tooth eruption disorder (Dutra et al., 2013; Elhaddaoui et al., 2017). Previous studies on tooth eruption mainly focused on changes in the molecular mechanisms associated with osteoclasts (Chidiac et al., 2018; Kwon et al., 2016). There is less research on how osteoblasts initiate differentiation, induce osteoclasts, and promote the formation of eruption channels.
There are a limited number of studies assessing the regulation of POSTN, the mechanism underlying its induction of osteoblasts, and functional changes in osteoblasts after POSTN silencing. Therefore, this study aimed to construct a mouse osteoblast virus carrier targeting POSTN, and then carry out a related experimental study on the changes in osteoblast function after POSTN silencing, to understand the theoretical basis for the study of tooth eruption.
The methods of silencing gene expression mainly include targeting the genomic DNA level or the RNA level (Broderick and Jones, 2014; Han et al., 2014). The earliest and most mature DNA level technology is gene knockout technology. The advantage of this approach is that the gene inactivation effect is accurate and reliable. However, the disadvantages are that the technology is complex, time-consuming, and laborious. The study of RNA levels refers to RNA interference, which leads to the degradation of homologous dsRNA in cells caused by the introduction of exogenous or endogenous double-stranded RNA (doubles transRNA, dsRNA), which inhibits the corresponding gene expression.
Compared with gene knockout technology, RNA interference technology is relatively simple to operate, and gene silencing is reliable (Foda and Singh, 2015; Park et al., 2013; Suresh et al., 2016). In this experiment, POSTN silencing was achieved using RNA interference technology by exogenous lentivirus transfection.
The virus's affinity to cells can vary due to the different species and activity of the cells. The preexperiment results showed that, for the mouse osteoblast-like MC3T3-E1 cells, the virus was more easily transfected by the MOI = 100 in the Eni medium containing 5 μg/mL polybrene. Then, the MC3T3-E1 cells were infected by lentivirus according to the preliminary experimental conditions for 10 hours. The cells were then divided into four groups according to the viral number: group NC; group KD1; group KD2, and group KD3. The medium was replaced at 16 hours after infection, and the target gene expression was observed by Celigo Image Cytometer at 72 hours after infection.
The results showed that the cells in each group were in good condition and no substantial cell death occurred. The fluorescent marker lentiviral infection results showed there was no green fluorescence in either group NC (Fig. 1A) or group KD2 (Fig. 1C). However, there was a small amount of fluorescent expression in group KD1 (Fig. 1B) and obvious green fluorescence in group KD3 (Fig. 1D). The real-time PCR results showed that compared with the control group, group KD3 received the best POSTN silencing effect (Fig. 4 and Table 1).
Through this experiment, we screened out the virus that specifically inhibited POSTN in osteoblast, and preliminarily determined the time of its action and the corresponding measurement. This information was used as an important reference in the later experiment. We then chose group KD3 as the experimental group for our further functional study.
We previously reported that osteoblasts themselves robustly induce autophagy, and the expression of autophagy-related proteins was altered significantly by the action of NF-kappa B inhibitor SN50. The results suggested that the autophagy function of osteoblasts during tooth eruption plays an important role through the NF-kappa B signaling pathway (Qin et al., 2016). As an important regulator of osteoblasts, it is important to determine how POSTN relates to the autophagy function of osteoblasts and how it regulates the dynamic balance between osteoblasts and osteoclasts.
In this experiment, autophagy factor proteins (Beclin-1 and LC3) were detected from the KD3 group of osteoblasts that had the best POSTN silencing effect. Beclin-1 and LC3 are homologs of yeast autophagy-related genes in mammalian cells and are important genes involved in autophagy regulation (Chen et al., 2016; Jacquin et al., 2017; Kang et al., 2018). Both genes have been used as important indicators for detecting autophagy activity due to their simple detection and high specificity (Jin et al., 2016; Kwon et al., 2017; Sui et al., 2018). The western blotting results showed that Beclin-1 protein expression was significantly higher in the experimental group than in the control group (Figs. 5 and 6). However, LC3 protein expression was not significantly changed between group KD and group NC (Figs. 7 and 8).
There are several possible reasons for this finding: (1) Autophagy is a complex process and previous studies reported that a variety of signal pathways participate in autophagy regulation (Efeyan et al., 2015; Lee et al., 2014; Noh et al., 2016), also, there are multiple signal molecules in each pathway and these various signal molecules can affect the autophagy level of cells in various ways.
(2) The roles of Beclin-1 and LC3 in the autophagy process are different. LC3 is located on the surface of the anterior autophagy bubble, and the autophagy bubble participates in the formation of autophagy. The Beclin-1 protein mainly forms an axial complex by the combination of type III phosphatidylinositol kinase and P150, which plays an important regulatory role in the development of autophagy (He et al., 2018; Wu et al., 2017). The expression of Beclin-1 and LC3 can be irrelevant.
(3) Different factors have different effects on autophagy function in different cells (Liu et al., 2018; Shi et al., 2014). Our study suggests that POSTN may inhibit the autophagy function of osteoblasts, which provides a new idea for the study of the molecular mechanism of tooth eruption.
This experiment successfully constructed the lentiviral vector targeting Periostin in MC3T3-E1 cells. We initially found that the autophagy function of osteoblasts may be enhanced when the POSTN was silenced. This finding will provide a theoretical basis for the further study of the functional changes of osteoblasts after POSTN silencing and the mechanism controlling the dynamic balance of osteoblasts and osteoclasts during tooth eruption. Importantly, because this part of the research is only a preliminary experiment, the detection indicators are limited. Thus, the research is not definitive. Our next study will focus on the changes of osteoblast-related factors and cell differentiation function in mice after POSTN silencing, and we will observe changes in osteoblasts differentiation function in animal models.
Footnotes
Acknowledgment
This research was funded by the Natural Science Foundation of China [Grant number: 81500893].
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.